

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Angiogenesis or neovascularization is a complex process, referring to the formation of new blood vessels from pre-existing ones and plays an important role in several vital processes including wound healing, restoration of blood flow to tissues after injury, and menstruation [1]. However, when the body is unable to control angiogenesis, due to proangiogenic factors becoming dominant over the antiangiogenic factors, diseases, such as age-related macular degeneration, rheumatoid arthritis, or psoriasis can occur [2]. Most importantly, angiogenesis promotes growth, proliferation, invasion, and metastasis of solid tumors [3], which in turn secrete a variety of angiogenic factors capable of stimulating angiogenesis as well [4].
Formation of new blood vessels is an essential step in tumor development, as without blood supply the tumor volume will not exceed more than 1-2 mm3 [5]. This in itself is a multi-step process that begins with degradation of the basement membrane by activated endothelial cells that migrate and proliferate, leading to formation of solid endothelial cell sprouts into the stromal space [6]. In due course, vascular loops are formed and capillary tubes develop with formation of tight junctions and deposition of a new basement membrane [7]. This is critical for tumor growth and metastasis, as these newly formed blood vessels provide tumor cells with the required nutrients and oxygen. Each step of this process represents a potential target for the inhibitory action of angiosuppressive drugs, allegedly leading to the desperately needed breakthrough in cancer therapy and other proangiogenic diseases [8].
To date, in excess of 300 angiosuppressive inhibitors have been reported and more than 80 drugs derived from them are in different phases of clinical trials [9]. Most angiosuppressive drugs are effective against a variety of tumors although high dosages of these substances are prescribed in order to suppress them in the event of any treatment strategy. In this perspective, natural resources hold great promise for discovery of novel antiangiogenic constituents. In fact, they have yielded a range of antiangiogenic constituents, including shark cartilage, curcumin, ω-3 and ω-6 fatty acids, green tea, licorice, quercetin, squalamine, and vitamin D3 [10]. They all have diverse mechanism of action but work by disrupting existing tumor vasculature. Besides, they can act synergistically with conservative treatments and are liable to have non-overlapping toxicities. Discovery of new potential therapeutic angiosuppressive drugs having all these properties would therefore be a valuable addition to the already growing pharmaceutical drug development initiatives.
Despite recent trends in the pharmaceutical industry toward rapid growth of synthesized de novo drugs, plants remain a traditional source of medicinal compounds; with upto 40% of modern drugs directly or indirectly linked to natural compounds. Marine plants such as seaweeds are believed to be beneficial for human health due to the myriad bio-protective activities that they possess. Additionally, they are used for a variety of purposes in the pharmaceutical/cosmetic industries for a variety of purposes due to their rich mineral and essential trace elements [11]. Since ancient times, seaweeds have also been consumed as human food, particularly in the regions surrounded by China, the Korean peninsula and Japan.
By and large, plants possessing an extensive array of pharmacological activities including antioxidant, anti-inflammatory, cardioprotective, and anticancer/antiangiogenic have been reported to be rich in carotenoids, polyphenols and triterpenes [12,13,14,15]. Also, some natural algal polysaccharides have been reported to suppress angiogenesis [16,17,18,19,20,21,22]. For this reason, seaweeds rich in carotenoids, polyphenols, polysaccharides, triterpenes are more inclined to possess angiosuppressive activity. A number of studies have indicated that marine seaweeds from India exhibit potent antioxidant properties [12,13,14,15,23,24,25]; however, they have not been systematically explored for their angiosuppressive potential. To the best of our knowledge, this is the first report on the angiosuppressive properties of marine Indian seaweed extracts.
In the current study, we primarily screened 15 marine seaweed extracts for their anti-proliferative activity against EAT (Ehrlich ascites tumor cells) and consequently selected a promising seaweed, Stoechospermum marginatum (C. Agardh) Kutzing for further investigations using both in-vitro and in-vivo models. It is hoped that the results will provide new insights for further research on angiogenesis using marine seaweed extracts from Indian coasts.
Go to : 
MATERIAL AND METHODS
Chemicals and bioreagents
3[H] Thymidine was purchased from Bhaba Atomic Research Centre, Mumbai, India. Dulbecco's Modified Eagle's Medium (DMEM), Fetal Bovine Serum (FBS), Penicillin-Streptomycin and Trypsin-EDTA were purchased from In-vitrogen, USA. Endothelial Growth Medium (EGM-2) was obtained from Cambrex Bioscience, Walkersville, USA. The antibodiy PECAM-1 and ABC reagent for CD 31 staining were procured from Santa Cruz Biotechnology, California USA. Matrigel was purchased from Chemicon International, CA, USA.
Collection of seaweed samples and preparation of their extract
The seaweeds Amphiroa fragilissima (Delile) Trevisan, Asparagopsis taxiformis (Linnaeus) Lamouroux, Caulerpa peltata J.V. Lamouroux, Caulerpa racemosa (Forsskal) J. Agardh, Caulerpa taxifolia (Vahl) C. Agardh, Chlorodesmis fastigiata (C.Agardh) Ducker, Codium elongatum (Turner) C. Agardh, Dictyopteris australis Lamouroux, Dictyopteris deliculata Lamouroux, Sargassum marginatum (C. Agardh) J.Agardh, Stoechospermum marginatum (C.Agardh) Kutzing, Padina Gymnospora (Kützing) Sonder, Padina tetrastromatica Hauck, Spatoglossum asperum J. Agardh, Spatoglossum variabile Figari and De Notaris were collected from in and around the coasts of Goa and Maharashtra, India. The seaweeds were botanically identified by a taxonomist and the identification was confirmed by comparing with the already available voucher specimens at the Herbarium of the National Institute of Oceanography, India.
Shade dried seaweed powder (1kg) was extracted with 4 Ltr methanol successively three times after keeping at room temperature for 24 h. The organic extracts obtained were filtered and evaporated to dryness using a rotary evaporator (Roteva, India) in order to obtain a semisolid residue. The methanolic extracts obtained were further diluted in appropriate cell culture media in order to obtain the required concentrations needed for further analysis. The Stoechospermum marginatum extract was designated as SME for future reference.
Cell lines, cell culture and animals
HUVEC (Human Umbilical Vein Endothelial) cells were obtained from Cambrex Bioscience, Walkersville, USA and cultured in EGM-2 medium with 2% FBS, 0.04% hydrocortisone, 0.1% long R3-human Insulin like growth factor (IGF-1),0.1% ascorbic acid, 0.4% human fibroblast growth factor (bFGF), 0.1% VEGF, 0.05% gentamycin and 0.05% amphotericin-B according to the manufacturer's protocol. BeWo (Choriocarcinoma) cells were obtained from the National Center for Cell Science, Pune, India and cultured in DMEM Ham's F-12 medium (Supplemented with 10% FBS, 1% Penicillin-Streptomycin and Gentamycin). HEK 293 (Non-transformed Human embryonic kidney) cells were obtained from the National Center for Cell Science, Pune, India. EAT cells (Mouse mammary carcinoma) were maintained in the Laboratory of Molecular Oncology, Department of Studies in Biotechnology, University of Mysore, Mysore, India and used routinely for in-vivo transplantation. Both EAT and HEK 293 cells were maintained in DMEM with 10% FBS and 1% Penicillin- Streptomycin. All cells were grown in a humidified atmosphere, containing 5% CO2 at 37℃. When the cells reached confluency they were passaged by trypsinization using 0.025% trypsin/0.01% EDTA. Cells from passages 2-5 were used for the experiments.
Female swiss albino mice (8-10 weeks old) and male Wistar rats weighing 300-350 g were obtained from the animal house, Department of Zoology, University of Mysore, Mysore, India. Fertilized eggs were acquired from a government poultry farm in Bangalore, India. All experiments were approved by the committee for the purpose of control and supervision of experiments on animals (CSPCA), Government of India, and performed strictly according to the NIH guidelines for the care and use of laboratory animals (IACUC Registration No: 122/99/IACUC).
In-vitro assay of proliferation
3[H] Thymidine incorporation assay was performed as described previously [26]. In brief, the tumor cells were plated onto 12-well culture plates at 25 × 103 cells/well and incubated at 37℃ in 5% CO2 for 48 hrs in DMEM medium. Following incubation, seaweed extract at 100 µg/ml concentration was added to the wells in triplicate, leaving three wells as control prior to addition of 3[H] thymidine (1 µci/ mL) to all wells. After incubation for 48 hrs, the cells were washed with PBS and then fixed with 10% ice-cold trichloroacetic acid in order to precipitate the DNA. The Radioactivity was measured in scintillation solution using liquid scintillation spectrometry. The concentration of the sample was then plotted against the percentage cell survival.
Matrigel tube formation assay
The matrigel tube formation assay has been widely been used as an in-vitro measurement of endothelial cell differentiation and as such it was performed according to the manufactures instruction (In vitro angiogenesis assay kit, Millipore). In brief, matrigel (50 µl) was added to each well of a 96 well plate, which was then incubated at 37℃ for 1 hr allowing the gel to polymerize. Human Umbilical Vein Endothelial Cells (HUVECS) were suspended in endothelial growth medium and cells (1 × 104) were seeded in to each well containing 150 µl of EGM. The cells were incubated with or without the test sample (SME) at 37℃ and 5% CO2. All of the conditions were performed in triplicates. After incubation for 16 - 20 hrs, the tube formation was photographed at 40 × magnification using an inverted microscope (Olympus, Germany).
Migration of HUVECs in the wound-healing assay
A wound-healing assay was performed for measurement of the in vitro effect of S. marginatum extract on the unidirectional migration of HUVECs according to Liang et al. [27]. In brief, HUVECs were seeded at 2 × 105 cells per well in a 6-well plate, and incubated overnight with complete medium (EGM-2 with 1% FBS) at 37℃ in a humidified atmosphere of 5% CO2. The cells were then serum starved overnight and a wound was scratched on the monolayer using a sterile pipette tip. The plates were washed with PBS twice for removal of any detached cells. For stimulation, VEGF (10 ng ml-1) was added, with or without S. marginatum extract at 0.1mg ml-1, and incubation continued for 3, 6, 18, 24, 36 and 42 hrs. The images were taken at the time of the wounding and at different time intervals thereafter using a phase-contrast microscope (Olympus, Germany).
Chorioallantoic membrane assay (CAM)
CAM assay was performed according to the method of Gururaj et al. [28]. In brief, the fertilized eggs were incubated at 37℃ in a humidified and sterile atmosphere for 10 days. Under aseptic conditions, a window was made on the eggshell to check for proper development of the embryo. The window was resealed and further development was allowed. On the 12lh day, saline, recombinant VEGF (50 ng per egg) or the extract (100 µl/ egg) was air dried on sterile glass cover slips. The window was reopened and the cover slip was inverted over the CAM. The window was closed again, and the eggs were returned to the incubator for two more days. The windows were opened on the 14th day and inspected for changes in microvessel density in the area under the cover slip and photographed.
Corneal Neovascularization (CNV)
The rat corneal micro-pocket assay was performed as described previously [29]. In brief, hydron polymer (poly hydroxyl ethylmethacrylate was dissolved in ethanol to a final concentration of 12%. A 5 µl aliquot of this mixture was then pipetted onto Teflon pegs. Aliquots of 10 µl of 12% Hydron/Ethanol alone (group 1), with 1 µg of cytokine VEGF (group 2) and VEGF + 5 µg seaweed extract (group 3) was added to each pellet and allowed to dry under a laminar flow hood at room temperature for 2 hrs. The pellets were incubated at 4℃ overnight. All procedures were performed under sterile condition. Male Wistar rats weighing 300-350 g were anaesthetized with i.p with a combination of ketamine (87 mg kg-1) and xylazine (13 mg kg-1). A drop of 0.5% proparacaine was instilled into the eye and the globe was proptosed using a pair of 0.3 mm tissue forceps. Using a surgical microscope, a paracentral linear incision 1 mm from the center of the cornea, 1.5 nm in length and 50% of the corneal depth was made using a No. 11 surgical blade to create a corneal micropocket. The incision was bluntly dissected through the stroma to the limbal area using a curved iris spatula. A single pellet was advanced into the lamellar pocket to the limbus using corneal forceps. Postoperatively, gentamycin ointment was applied to the anterior surface of the operated eye. The rats were observed for 24-72 hrs for occurrence of non-specific inflammation and for localization of the pellets. On day 7, the rats were anaesthetized with ketamine and the corneas were observed under a stereo-binocular microscope with a CCD camera and photographed.
Record of body weight, ascites volume, cell number and peritoneal angiogenesis of EAT hearing mice treated with S. marginatum methanolic extract
The crude extract of the seaweed S. marginatum selected for the study was tested for its effect on EAT cell growth in-vivo [30]. EAT cells (5 × 106) were injected intraperitoneally (i.p) into mice (two groups of mice, six in each group) and growth was recorded each day from the day of transplantation. To determine whether the seaweed extract supressed tumor growth and angiogenesis mediated by EAT cells in-vivo, the extract (l00 µg of SME) was injected into the peritoneum of EAT bearing mice each day from the sixth day of transplantation. The body weight of the mice was monitored from the first day until the twelfth day. On the twelfth day, the animals were sacrificed and the volume of the ascites formed in both untreated and treated mice was recorded. The pelleted cells were counted using the trypan blue dye exclusion method using a hemocytometer. The animals were then dissected in order to observe the effect of the extract on peritoneal angiogenesis.
Immunohistological analysis (H & E staining)
To determine whether the SME inhibited the microvessel density, the effect of the extract was verified on the angiogenic response induced by cytokine VEGF in EAT bearing mice. EAT bearing mice were treated regularly with the extract from the sixth day of transplantation. On the twelfth day, the animals were sacrificed and the peritoneum from treated or untreated mice was fixed in 10% formalin. Sections (5 µm) were made from paraffin embedded peritoneum using an automatic microtome (SLEE Cryostat) and stained with hematoxylin and eosin. Microvessel counts and image photography were performed using a Leitz-DIAPLAN microscope with an attached CCD camera.
CD 31 Immunostaining
The effect of SME on proliferating endothelial cells was determined by staining the paraffin sections of the peritoneum of treated or untreated mice with anti-CD 31 anti-bodies [31]. Peritoneum sections were processed according to the protocol supplied by the manufacturer (Santa Cruz Biotechnology, CA, USA). In brief, sections were dewaxed in xylene three times for 5 minutes each. The sections were rehydrated in descending concentrations of ethanol (100% ethanol for 5 minutes, 95% for 2 minutes and 80% for 2 minutes) and washed in distilled water. For antigen retrieval the sections were heated at 95 ℃ for 15 minutes in a humidified atmosphere. The sections were treated with 3% H2O2 in PBS to block endogenous peroxidase activity. They were blocked in blocking serum for 30 minutes in order to reduce non specific binding and were then incubated with anti-CD 31 (PECAM-1) anti-bodies for 2 hrs. Following washing with PBS, slides were incubated with secondary anti-body (biotinylated rabbit anti- mouse IgG) for 1 hr at room temperature. The slides were washed in PBS for 5 minutes and incubated with the substrate (100 µl/ section) followed by ABC reagent for 45 minutes (2 ml histo buffer + 20 µl Avidin solution + 20 µl Biotin solution). After incubation, the slides were washed in PBS for 5 minutes. Antigen and antibody complexes were detected using substrate (DAB, 100 µl/ section) for 5 minutes. The sections were washed thee times for 2 minutes in tap water and twice for 2 minutes in distilled water. Subsequently, the slides were counter stained with 2% hematoxylin for 5-7 minutes and washed again in tap water three times for 5 minutes each. The slides were washed effectively for two minutes each in 50% ethanol, 80% ethanol and absolute alcohol. After xylene wash, the slides were mounted using entellan mountant solution and the sections were scored using DIAPLAN light microscope and photographed.
Statistical analysis
All experiments were performed in triplicate (n = 3) at least and values are expressed as mean ± SD. STATISTICA software (Statsoft, 1999) was utilized for comparison of the mean values of each treatment). Data were analyzed by one way analysis of variance (ANOVA) for determination of significant differences. P-values < 0.05 were considered significant and P-values < 0.001 highly significant.
Go to :
RESULTS
S. marginatum extract inhibits in vitro proliferation of EAT/ BeWo cells
Preliminary screening of SME demonstrated potent antiproliferative ability against EAT cells (Fig. 1); therefore, it was selected for further anti-angiogenic studies. BeWo cells and HEK-293 cells were used to determine whether SME inhibits proliferation of tumor/normal cells in-vitro. SME extract inhibited proliferation of BeWo cells in a dose-dependent manner. However, no potent effect was seen in untransformed normal HEK-293 cells. As shown in Fig. 2A, inhibition of proliferation was 80.55, 83.54, 90.79, 95.43, and 96.36% on the proliferation in BeWo cells and 4.62, 5.76, 5.85, 6.71, and 7.40% was recorded, while in HEK 293 cells at 0.005, 0.025, 0.050, 0.075, and 0.1 mg ml-1 concentrations of SME with P < 0.001 (Fig. 2B).
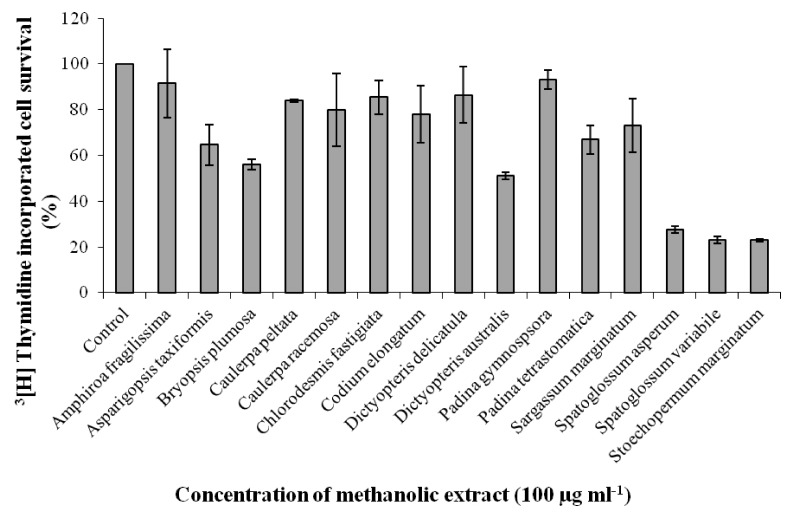 | Fig. 1
Effect of 15 seaweed methanolic extracts on proliferation of EAT cells in-vitro. EAT cells were plated in 12 well plates and incubated for 48 h. Seaweed extracts of 100 µg ml-1 concentration were added to the wells in triplicate prior to addition of 3[H] thymidine followed by incubation for another 48 h. The cells were trypsinized after two days and processed for scintillation counting. Values are presented as mean ± SD (n = 3).
|
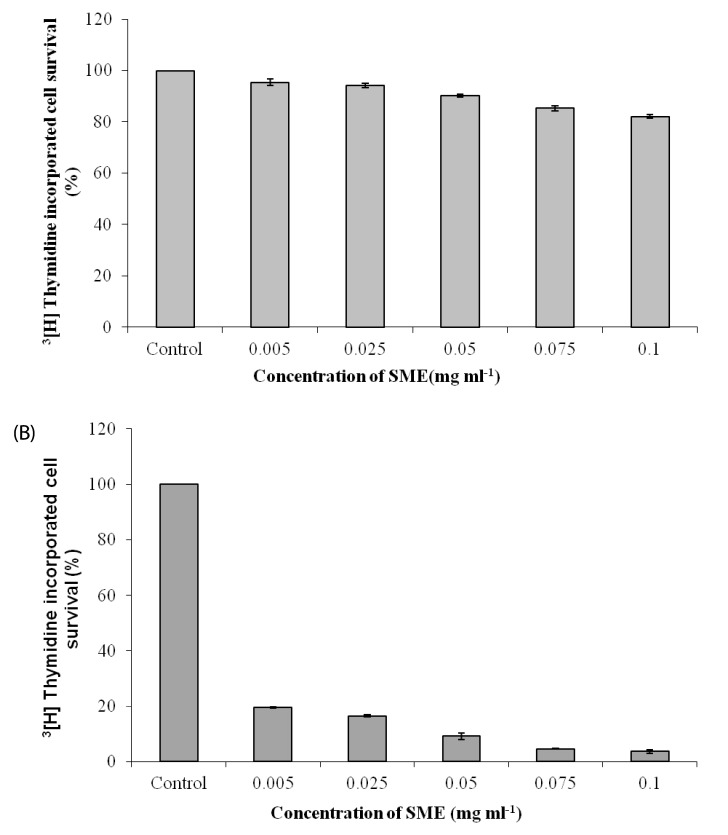 | Fig. 2
Effect of S. marginatum extract (SME) on proliferation of HEK-293 and BeWo cells in-vitro. (A) HEK-293 and (B) BeWo cells were plated in 12 well plates and incubated for 48h. SME in concentrations of 0.005, 0.025, 0.05, 0.075 and 0.1 mg ml-1 was added to the wells in triplicate prior to addition of 3[H] thymidine followed by incubation for another 48 h. The cells were trypsinized after two days and processed for scintillation counting. Values are presented as mean ± SD (n = 3).
|
SME inhibits VEGF induced in-vitro tube formation of HUVEC's
Tube formation assay was performed in-vitro in order to substantiate the effect of SME on formation of blood vessels by HUVEC's. In the positive control group stimulated with VEGF (10 ng), the HUVEC's adhered to the matrigel surface within 20 to 24 hrs and formed a branching anastomising network of capillary like tubes with multi-centric junctions over 24 hrs. However, treatment with SME prevented VEGF stimulated tube formation of HUVEC's, in a dose dependent manner (Fig. 3). Complete inhibition was achieved at 5 mg ml-1of SME. At the latter concentration, most of the cells appeared as unorganized cell aggregates.
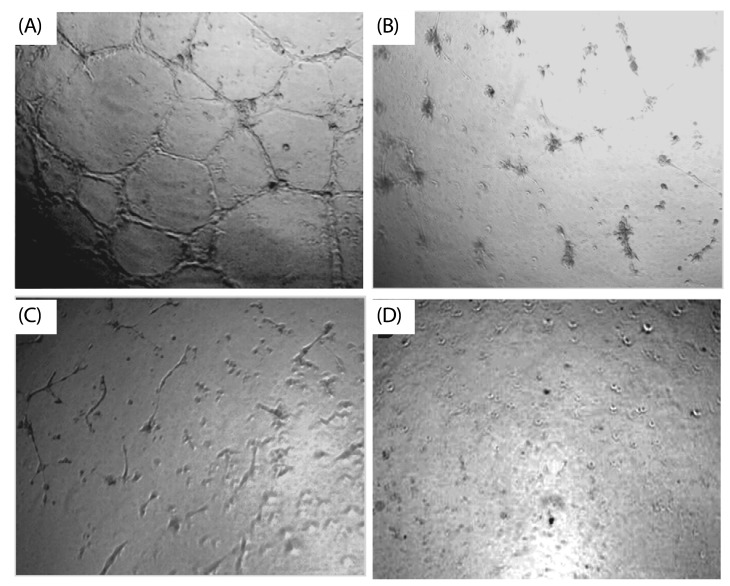 | Fig. 3
Inhibitory effect of S. marginatum extract (SME) on VEGF induced tube formation in-vitro. HUVEC's were seeded into the matrigel layer in a 96-well plate. (A) VEGF alone (+ ve control), (B) without VEGF (-ve control), (C) VEGF + SME (5 µg/ well), (D) VEGF + SME (10 µg/ well). The experiment was repeated three times with similar results (values are presented as mean ± S.D; n = 3). Three replicate fields of triplicate wells were digitally photographed.
|
Effects of SME on HUVEC migration in the wound healing assay
To determine whether cell migration was inhibited by SME, in-vitro wound healing assays was performed using HUVEC cells, as described in the Materials and Methods section. Cell migration of HUVECs treated with VEGF increased in control wells while, migration in cells treated with SME at 100 µg ml-1 was effectively inhibited in the wound (A) healing assay (Fig. 4).
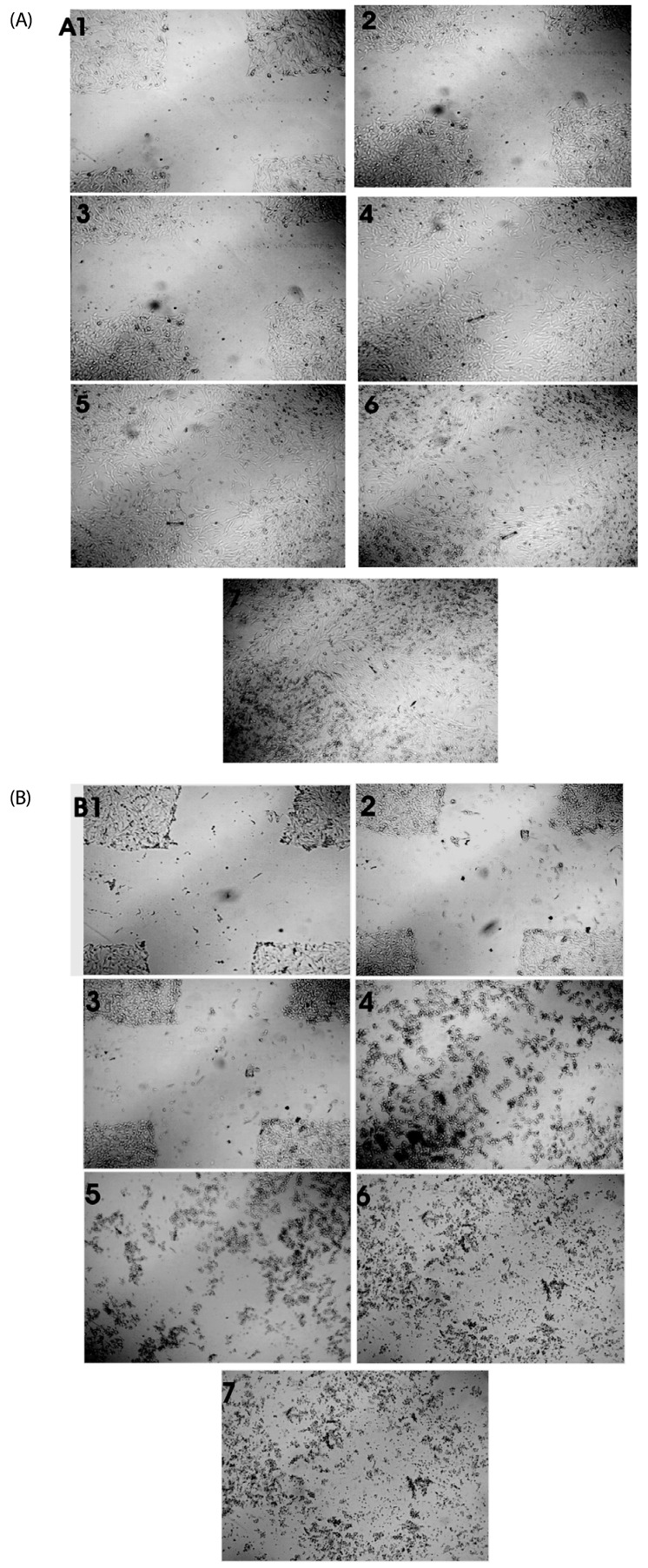 | Fig. 4
Effect of S. marginatum extract (SME) on HUVEC cell migration in an in-vitro scratch wound healing assay. The representative phase-contrast images show migration of cells into the wounded area. (A) Wound closure in control wells at 1) 0h, 2) 3h, 3) 6h, 4) 18h, 5) 24h, 6) 36h and 7) 42h. (B) Wound closure in SME (100 µg/ well) treated wells at 1) 0h, 2) 3h, 3) 6h, 4) 18h, 5) 24h, 6) 36h and 7) 42h.
|
Angio-suppressive effect of SME on CAM and CNV
The CAM and rat cornea assays form a part of the conventional angiogenesis assays commonly used for in-vivo validation of the angio-suppressive efficacy of anti-angiogenic molecules. Results shown in Fig. 5A and Fig. 5B indicated that SME had a direct effect on inhibition of angiogenesis in an in-vivo model system. When compared to the extensive angiogenesis seen in VEGF treated CAM and rat cornea, angiogenesis at the site of application of SME was significantly reduced.
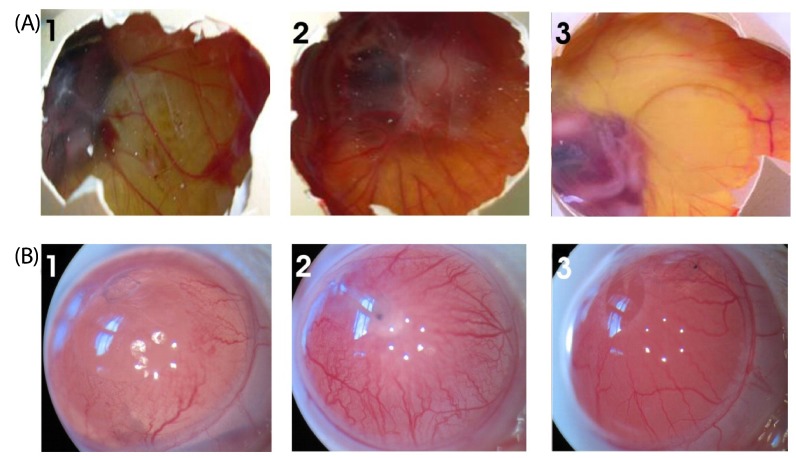 | Fig. 5
Effect of SME on blood vessel regression in the chick CAM and rat cornea assays in-vivo. (A) Photographs of VEGF-induced neovascularization observed in CAM: (1) Saline (- control), (2) VEGF alone (+ control), (3) VEGF + SME (100 µg) was applied to the CAM of 11-day-old chicken embryos. After incubation for 48h, the treated area was inspected for changes in neovascularization. The arrows indicate the treated area. The data shown represent the result of an experiment performed using a maximum of six eggs in each group. All photographs were taken at 40 × magnification. (B) Photographs of VEGF-induced neovascularization observed in rat corneas: (1) hydron polymer + VEGF (1 µg) (+ control), (2) hydron polymer alone (- control), and (3) hydron polymer + VEGF + SME (100 µg). After incubation for seven days, the corneas were photographed at 40 × magnification.
|
In-vitro treatment with SME extract inhibits growth of EAT cells in the peritoneal angiogenesis assay
The treated as well as control animals were observed for the degree of vascularization occurring in the peritoneal cavity. The results shown in Fig. 6A demonstrate that control EAT bearing mice showed a gradual increase in body weight of 8 to 10 g when 5 × 106 EAT cells were injected on day zero. In mice treated with/ without SME, a significant decrease was observed in groups treated with SME when compared to the control, indicating the effect of SME in preventing the growth of tumor cells (P < 0.05). In a fully grown ascites tumor, a volume of 6.25 ± 0.22 ml of ascites was generated during the tumor growth period of 12 days. In SME treated mice, the volume of ascites was approximately 2.6 ± 0.32 ml with P < 0.001 (Fig. 6B). The numbers of viable cells in fully grown EAT bearing mice was 3.25 ± 0.24 × 108/ mouse, while this number was reduced to 0.64 ± 0.06 × 108/ mouse with P < 0.001 in SME treated mice (Fig. 6C), indicating a considerable reduction when compared with the control. These results indicate the potential anti-tumor activity of SME. In a fully grown ascites tumor in-vivo, there is extensive peritoneal angiogenesis. However, in SME treated mice, a significant decrease in peritoneal angiogenesis was also observed in-vivo (Fig. 6D).
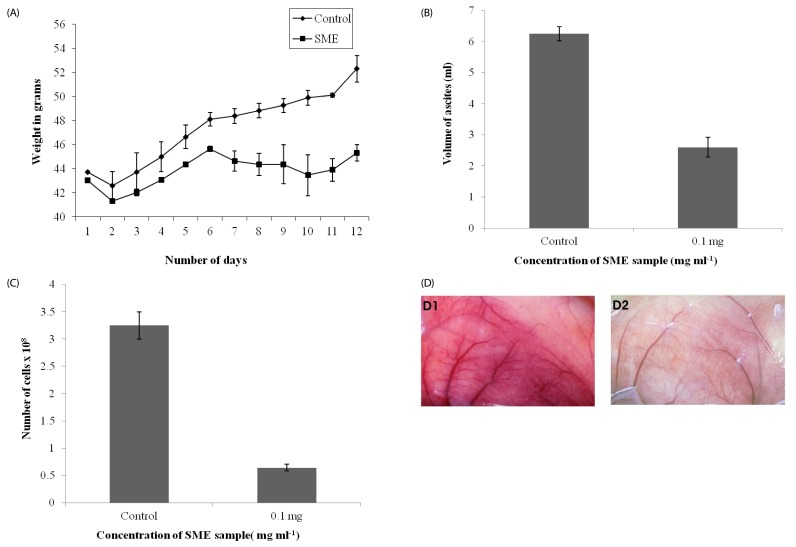 | Fig. 6
In-vivo inhibition of tumor growth and angiogenesis by S. marginatum extract (SME). (A) Body weights of EAT-bearing untreated mice or mice treated with SME were recorded. From the sixth day onward, SME (4 mg kg-1 body weight) was administered (i.p) every day for six days; the animals were sacrificed on the 12th day. (B) EAT cells were collected along with ascites fluid and measured, (C) Cells were counted using a haemocytometer, (D) The peritoneum of the animal was photographed: (1) Control, (2) SME treated. At least six mice were used in each group and the results obtained are an average of three individual experiments and mean of ± SD (n = 6 per group).
|
In addition, the effect of SME on survival of EAT bearing animals was tested. Upon intraperitoneal transplantation of 5 × 106 cells/ mice, the EAT-bearing mice succumbed to the tumor burden 13 days after tumor transplantation, due to increase in the number of tumor cells with passage of time, whereas the animals treated with SME (4 mg kg-1 body weight/dose, every day) extended the survival time of EAT-bearing mice from 13 days up to one month (data not shown).
SME inhibits formation of MVD and proliferation of endothelial cells
Results of H & E staining indicated that there was a reduction in the number of newly formed microvessels in the peritoneum of SME treated peritoneum of EAT bearing mice, compared to the control (Fig. 7A). The proliferation of endothelial cells can be confirmed using CD31 marker. Our results of CD31 staining, showed a marked reduction in the number of proliferating endothelial cells in the peritoneum of SME treated EAT-bearing mice (Fig. 7B), corroborating the results shown in the inhibition of peritoneal angiogenesis in-vivo.
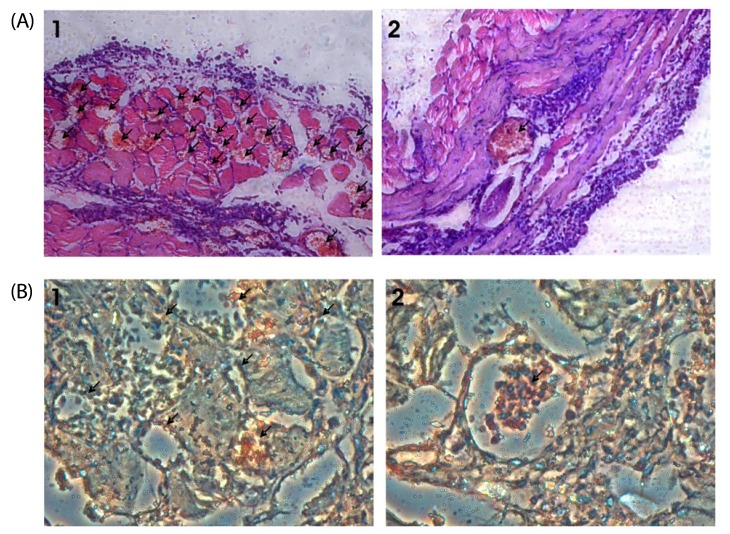 | Fig. 7
S. marginatum extract (SME) inhibits MVD and proliferation of endothelial cells in mouse peritoneum. (A) The peritoneums of control (1) as well as SME-treated (2) EAT-bearing mice were embedded in paraffin and 5 µm sections were made using a microtome. The sections were stained with hematoxylin and eosin and observed for microvessel density (40×). Arrows indicate the microvessels. (B) Paraffin sections (5 µm) of peritoneum of control (1) and SME (2) mice were immunostained with anti-CD31 (PECAM) anti-bodies. Arrows indicate the stained activated endothelial cells.
|
Go to :
DISCUSSION
The hypothesis that anti-angiogenic compounds can be used in combination with cytotoxic drugs for tumor therapy has recently been proposed [32]. And, since anti-angiogenesis has become a major issue in cancer treatment, the search for chemicals expressing anti-invasive and angio-suppressive properties rather than direct cytotoxic effect on cancer cells has gained significant momentum in recent years of modern drug development. To date, several angiogenesis inhibitors have already been developed, whose efficacy has been evaluated by means of different in-vitro and in-vivo assays and their clinical evaluations are in progress [33]. Additionally, chemotherapeutic agents have shown anti-angiogenic properties both in-vitro and in-vivo [34,35], leading to the idea of chemotherapy with an anti-angiogenic setup [35,36].
Keeping all of these facts in mind, we attempted to screen for potent anti-proliferative compounds in marine seaweed extracts in-vitro and evaluated the angio-suppressive effect of a promising seaweed S. marginatum both in-vitro and in-vivo, for the first time.
The antiproliferative effect of crude SME was evaluated using EAT and BeWo and HEK 293 cell lines. The methanolic extract showed strong inhibition of proliferation of the tumor cell lines. Data on the antiproliferative effect of SME (100 µg/ ml) on EAT is shown in the Fig. 1, while data on its effect on BeWo and HEK 293 cells are shown in Fig. 2A and 2B. Our results demonstrate that SME powerfully inhibits cell proliferation of EAT and BeWo cells in-vitro, while having less effect on non transformed HEK 293 cells. Earlier, phytochemical investigations have reported the presence of phenols, tannins, saponins, cardiac glycosides, alkaloids, anthraquinones, and flavonoides in the seaweed extracts tested in this study [37]. Therefore, the antiproliferative activities of the seaweeds may be attributed to the presence of these compounds in their extracts.
Endothelial cells differentiate and form capillary-like structures when seeded on matrigel and forms an essential step of angiogenesis involving cell-matrix interaction, intercellular communication and cell mobility like in-vivo tumor angiogenesis. As shown in Fig. 3, untreated control wells containing HUVEC's showed formation of well structured capillary tubes within 9 h. While in the presence of SME (at 0.005 and 0.010 mg/well), the HUVECs rounded up and rendered partial network structures, indicating that it inhibited endothelial cell proliferation and consequently angiogenesis in-vitro.
VEGF promotes many of the events necessary for angiogenesis including endothelial cell migration through a variety of signaling pathways [38]. Therefore, the effects of SME on migration of HUVECs was evaluated using the wound healing assay and it was found to inhibit VEGF induced HUVEC migration (Fig. 4). The inhibitory effects of SME on VEGF induced migration may be attributed to inhibitions of endothelial proliferation.
The CAM assay was applied in order to verify the angio-suppressive effect of SME ex vivo. The CAM assay is based on angiogenesis taking place on the chick embryo chorioallantoic membrane of fertilized eggs and is frequently used in the study of both angiogenesis and anti-angiogenesis [39]. In the current study, the methanolic extract of S. marginatum (SME) was studied for its angio-suppressive activity by induction of an angiogenic response subsequent to stimulation of CAM with VEGF. After incubation for 48 hr, angiogenesis in the form of increased vessel density and a normal vascular pattern with numerous branching was observed in the control group around the implant, with the vessels radially converging toward the center. Conversely, when SME (100 µg/ egg concentration) was tested, the vessels became less intense and presented avascular zones around the insert showing suppression of new vessel development on the CAMs (Fig. 5A).
Topical application of angiogenic inhibitors is a beneficial route for prevention and treatment of CNV because it is non-invasive and has minimal systemic adverse effects. The current study established the effectiveness of SME by means of topical application in CNV. In fact, topical application of 5 µg/eye significantly prevented VEGF induced CNV as confirmed by morphological observation (Fig. 5B).
Our preliminary results clearly demonstrated that SME has a potent angiosuppressive effect on Ehrlich ascites tumor (EAT) cells in-vivo. Treatment of EAT bearing mice with SME resulted in a decrease in body weight, ascites volume and cell number in-vivo (Fig. 6) and also increased their survivability (Data not shown). VEGF in malignant ascites formation is known to play a pivotal role in accumulation of ascites fluid, tumor growth, and microvessel density [40,41,42]. Therefore, the antitumor effects of SME may be correlated with its angio-suppressive activity and this assumption is also supported by the reports of Folkman et al. [43]. Several research reports have already illustrated the anti-tumor potential of seaweeds in a variety of tumor cell types [44]. As a result, our results undeniably provide the proof that inhibition of angiogenesis could be at least one of the many mechanisms of action triggered by SME and may be a cause for repression of VEGF-like factors or secretion of such factors in the process.
The Hematoxylin-eosin staining method is usually used for examination of vascular invasion and is characterized by the presence of at least one tumor cell cluster, which is clearly evident in decorated vascular spaces, where endothelial cells are stained brown as described by Birner et al. [45] In the current study, peritoneum from mice with and without SME treatment was fixed in formalin. Sections (5 µm) were made from these paraffin embedded peritoneum and stained with Hematoxylineosin. Results were expressed as the highest number of microvessels in any single 40 × field. Final MVD was the mean score obtained from the hot spot areas counted using Leitz Diaplan light microscope, Germany, attached to CCD camera. In this study, the average MVD was significantly higher in control with vascular invasion than in test SME, suggesting a close association of angiogenesis with microvessel density of tissue and clinical aggressiveness of tumor (Fig. 7A).
CD31staining was performed in order to evaluate the inhibition of proliferating endothelial cells in the peritoneal lining of tumor-bearing mice. Under microscopic analysis, a marked decrease in peritoneal angiogenesis and levels of microvessels in CD31 stained sections of peritoneal wall of the treated group confirmed the angiosuppressive activity of SME when compared with that of the control (Fig. 7B). An earlier study has established that the density of microvessels is almost doubled in tumors from patients with metastasis [46]. While the presence of vascular invasion is not a consistent finding, it is still linked to increased incidence of lymph node, distant metastasis, and a corresponding decrease in survival [47].
In the current study, for the first time, we have shown that methanolic extract of S. marginatum contains strong angio-suppressive and anti-proliferative activities. The overall activity of SME may be ascribed to the abundant presence of several cytotoxic compounds, including fucoidans, laminarins and terpenoids in seaweeds, stated to possess anti-cancer properties. Taken together, our results highlight the distinctive antiangiogenic properties of SME and its promising application in cancer chemoprevention, while providing a solid groundwork for clinical implementation in humans. However, further investigations are essential in order to evaluate the comprehensive mechanism, the likely constituent(s) and to establish its favorable role in the clinical background.
Go to :
 XML Download
XML Download