

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Type 2 diabetes is characterized by a 2- to 4-fold increased risk of cardiovascular disease (CVD) [1]. This is generally attributed to the adverse effects of hyperglycemia and oxidative stress on vascular biology [2]. Endothelial progenitor cells (EPCs) play an important role in postnatal neovascularization and vascular repair [3,4]. As such, EPC dysfunction may be a cause of diabetic vascular complications [5]. In patients with type 2 diabetes, especially those with vascular complications, both the number and function of circulatory EPCs are known to be significantly reduced [6,7]. Furthermore, EPCs promote in vivo re-endothelization and are able to be incorporated into new vessels in animal models of hind limb ischemia [8]. EPCs are also involved in processes such as myocardial ischemia and infarction, wound healing, and endogenous endothelial repair [9,10]. Furthermore, in vivo animal model studies along with in vitro studies using EPCs from type 1 and type 2 diabetic patients have revealed a potential role for glucotoxicity in impairing EPC function [8,11,12]. In a clinical setting, high glucose (HG) conditions are usually accompanied by increased free fatty acid levels, which might have detrimental effects on endothelial cells [13]. More importantly, HG has been shown to inhibit both the colony-forming ability as well as proliferation of EPCs, whereas it enhances senescence of early and late EPCs; HG also was shown to impair the migration and vasculogenesis capacity of late EPCs [14,15]. P38 mitogen-activated protein kinases (p38 MAPKs) are critical regulators of cell differentiation [16]. In addition, activation of p38 MAPKs has been implicated in induction of cell apoptosis [17]. p38 MAPKs coordinately regulate cellular proliferation and differentiation processes induced by a variety of cellular stresses. Specifically, HG conditions can activate the p38 MAPK pathway in many cell types, including endothelial cells [18]. As glucose has been reported to activate p38, it could be a potential effector in signaling mechanisms impairing the proliferation and differentiation of EPCs.
Lycopene, which is a naturally present carotenoid in tomatoes and tomato products, is the most potent singlet oxygen quencher among all natural carotenoids [19]. Epidemiological studies have shown an inverse relationship between intake of tomatoes, serum lycopene levels, or adipose tissue lycopene levels and incidence of coronary heart disease [20]. An in vitro study indicated that lycopene appears to be the most effective in reducing both human aortic endothelial cell (HAEC) adhesion to monocytes as well as cell surface expression of adhesion molecules among five carotenoids, suggesting an important role for lycopene in attenuating atherogenesis [21]. Lycopene was also reported to attenuate the phenotypic and functional maturation of murine bone marrow-dendritic cells [22]. Additionally, lycopene was shown to down-regulate the expression of co-stimulatory molecules and major histocompatibility complex type II molecules, as it was able to inhibit MAPKs such as extracellular signal-regulated kinase 1/2 (ERK1/2), p38 and c-Jun N-terminal kinase (JNK), and the transcription factor Nuclear Factor kappa B (NF-kB) [22]. Lycopene may modulate the MAPK signaling cascade by regulating phosphorylation [23]. However, the protective effects of lycopene against HG-induced EPC injury as well as its underlying mechanism are not well established yet.
Based on these considerations, we tested the hypotheses that HG-induced EPC dysfunction is initiated by activation of p38 MAPK, resulting in apoptosis, and that the underlying protective effects of lycopene on EPCs involve modulation of p38 MAPK signaling pathways.
MATERIALS AND METHODS
Subjects
All subjects gave their informed consent, both verbally and in writing, after being informed about the study. Ethics approval for the trial was obtained from the ethical committee of the Guangzhou University of Traditional Chinese Medicine, China (Reference # KC08HISV0351), and the study was performed in accordance with the principles of the Declaration of Helsinki. The study enrolled 15 healthy medical personnel. Of these, eight were male and seven were female. The age range was from 29-52 years (mean 38.3 ± 12.1 years). All controls had no clinical history of diabetes or hyperlipidemia, and they showed normal fasting blood glucose (FBG) levels of < 7 mmol/L. They underwent a normal physical examination and had not received any medication. Exclusion criteria for participation were use of cigarettes, coffee, recent trauma, surgery, or dermal ulcers, as these all inhibit EPC proliferation, migration, vasculogenesis capacity, and survival rate [24,25,26].
Chemicals and materials
Lycopene (purity ≥ 90%, from tomato), human fibronectin (FN), FITC-labeled Ulex, 1,1-dioctadecyl-3,3,3,3-tetramethylindocarbocyanine (Dil), labeled acetylated low-density lipoprotein (acLDL), and SB203580 were purchased from Sigma-Aldrich (St. Louis, MO, USA). M199 medium and fetal bovine serum were obtained from Hyclone (USA). Lydroxypropylmethyl Cellulose was purchased from Tianjin Haoyang Biological Manufacture Co., Ltd. China (Haoyang, China). RrhVEGF, recombinant human basic fibroblast growth factor (rhbFGF), epidermal growth factor (EGF), and insulin-like growth factor 1 (IGF-1) were purchased from Pepro Tech (USA). Annexin V-FITC Apoptosis Detection Kit was purchased from Jingmei Biotech Co., Ltd. (Shanghai, China). Boyden chamber was purchased from Haimen City Qilin Medical Instrument Factory. Antibodies against p38 MAPK (product code 9212) and phosphorylated p38 MAPK (product code 9216) were purchased from Cell Signaling Technology (Danvers, MA, USA). Goat Anti-mouse IgG/HRP (product code ZDR-5307) was purchased from Zhongshan Golden Bridge Bio-technology Co., Ltd. (Beijing, China). All other chemicals were obtained from common commercial sources and were of reagent grade. Pure lycopene was dissolved in tetrahydrofuran (THF), stored at-70℃, protected from light [27], and diluted with medium to the desired concentration before use.
Isolation and cultivation of EPCs
Fasting peripheral venous blood sample (15 mL) was isolated from healthy medical personnel to obtain mononuclear cells by density gradient centrifugation following treatment with sodium heparin anticoagulant (within 1 h of delivery) [28,29].
The obtained mononuclear cells were cultured in fibronectin-coated 6-well culture plates. Then, 0.5 mL of M199 medium containing 20% fetal bovine serum (FBS), 10 µg/L of recombinant human vascular endothelial growth factor (rhVEGF), 2 µg/L of rhbFGF, 10 µg/L of EGF, 12 µg/L of IGF-1, 1×105 U/L of penicillin, and 1×105 U/L of streptomycin were added. The cell solution was cultured in a 5% CO2 incubator with saturated humidity at 37℃ for 4 days. Non-adherent cells were removed by washing with phosphate-buffered saline (PBS), after which attached early EPCs appeared. Adherent cells were incubated in fresh medium for another 4 days. After non-adherent cells were rinsed with PBS, the medium was changed for another 7 days of culture. Then, non-adherent cells were removed by PBS, and adherent cells were identified. Cultured cells were co-cultured with 2.4 mg/L of Dil-Ac-LDL at 37℃ for 1 h, followed by fixation with 20 g/L of paraformaldehyde for 10 min. After rinsing with PBS, 10 mg/L of FITC-labeled Ulex europaeus agglutinin I was added to the cell samples, which were cultured at 37℃ for 1 h. Under a laser scanning confocal microscope, Ulex lectin FITC-I and Dil-Ac-LDL double-stained positive cells were identified as differentiating EPCs [28,29]. Cell numbers were viewed with an inverted fluorescent microscope (Olympus).
Experimental grouping and procedures
Following culture in M199 medium without FBS, adherent cells were divided into six groups: normal control group (only M199 medium), mannitol group (30 mmol/L of mannitol and M199 medium, osmotic control), HG group (33 mmol/L of glucose) [30], and three lycopene groups according to dosage (each cultured with 33 mmol/L of glucose followed by lycopene administration at doses of 10, 30, and 50 µg/mL in M199 medium). Medium in each group was changed every 8 h, and the culture process lasted for 72 h. EPC proliferation and apoptosis were measured by MTT assay and flow cytometry at 12, 24, 48, and 72 h, respectively. Cells were incubated on the day of isolation with glucose/mannitol or lycopene; medium was not changed for the early EPC study, whereas cells under passage three were used for the late EPCs study. As above, EPCs were divided into six groups: normal control group, HG group, three lycopene groups according to dosage (each cultured with 33 mmol/L of glucose followed by lycopene administration at doses of 10, 30, and 50 µg/mL), and SB203580 (p38 MAPK inhibitor, 1 µmol/L) with 33 mmol/L of glucose group. The culture process lasted for 24 h. EPC migration, adhesion, vasculogenesis capacity, as well as phosphorylated and non-phosphorylated p38 MAPK protein expression levels were measured.
EPC proliferation and apoptosis
Freshly isolated early EPCs were used for treatment. The cells were treated with HG and/or different concentrations of lycopene throughout the 7-day experiment. Additional cells cultured with mannitol served as a control. The inert substance mannitol was used to maintain osmotic pressure. Cell proliferation and viability on day 7 were determined by MTT assay. After digestion with 0.25% trypsin and culture in M199 medium, the mononuclear cell suspension at a density of 1×104 cells per well was placed into fibronectin-coated 96-well plates, with each well containing a volume of 200 µL. Culture plates were placed into a 5% CO2 incubator with saturated humidity at 37℃ for 24 h. Then, 20 µL of a 5 g/L MTT solution was added to each well for continuous culture for 4 h. The supernatant was then removed, after which 150 µL of dimethyl sulfoxide (DMSO) was added to each well. A micro oscillator was used for 10 min to fully dissolve the crystals. The OD value was measured using an ELISA reader at 490 nm. The measurement was repeated three times, and the average values of absorbance were calculated.
Apoptotic EPCs were measured using an Annexin V-FITC Apoptosis Detection Kit. The cell concentration was adjusted to 109 cells/L following digestion and rinsing with PBS twice. The cell suspension (100 µL) was placed into a flow tube and mixed with 5 µL of AnnexinV/FITC and 10 µL of 20 mg/L propidium iodide (PI). The mixture was incubated at room temperature in the dark for 15 min and then diluted with 400 µL of PBS. According to flow cytometry, Annexin-V+PI- cells were considered to be early apoptotic cells, Annexin-V+PI+ were apoptotic and necrotic cells, Annexin-V-PI+ were damaged cells, and Annexin-V-PI- were living cells. Early apoptotic rate = number of apoptotic cells/total number of cells measured ×100%.
Adhesion assay of EPCs
Third generation EPCs as target cells were pretreated for 24 h with different concentrations of lycopene and HG (500 µL) and then counted. Identical cells (1×105) were placed into fibronectin-coated 96-well plates and incubated for 30 min at 37℃. Non-attached cells were gently removed with PBS, adherent EPCs were fixed with 4% paraformaldehyde, and adherent cells were counted by independent blinded investigators in five random high-powered (×200) microscopic fields in replicate wells. Three replicates were carried out, and the average was calculated.
EPC migration test
Migration of late EPCs was evaluated using a modified boyden chamber. In brief, isolated EPCs were detached using 0.25% trypsin, harvested by centrifugation, resuspended in 500 µL of M199 medium, and then counted. EPCs (2×104) were placed in the upper chamber of the modified boyden chamber. Vascular endothelial growth factor (VEGF) in M199 medium at 100 mg/L was then placed in the lower compartment of the chamber. After 24 h of incubation at 37℃, the lower side of the filter was washed with PBS and fixed with 2% paraformaldehyde. For quantification, cells were stained with Giemsa solution. Cells migrating into the lower chamber were counted manually in five random high-powered (×200) microscopic fields in replicate wells. Three replicates were carried out, and the average was calculated.
In vitro vascular tube formation
Tube formation assay was performed on late EPCs to assess vasculogenesis capacity, which is believed to be important in new vessel formation [31]. In brief, ECMatrix gel solution was thawed at 4℃ overnight, mixed with ECMatrix diluent buffer, and then placed in a 24-well plate at 37℃ for 1 h to allow the matrix solution to solidify. EPCs were harvested as described above with trypsin/EDTA, after which 1×104 EPCs were placed on matrix solution containing M199 medium with HG or different concentrations of lycopene, followed by incubation at 37℃ for 24 h. Tubule formation was inspected under five random inverted light microscope (×100) fields in replicate wells. Three replicates were carried out, and the average was calculated. Tubule formation was defined as a structure exhibiting a length four times its width. Five representative fields were taken, and the average of the total area of complete tubes formed by cells was compared using the computer software Image-Pro Plus.
Western blot analysis
Following EPC pretreatment for 24 h with different concentrations of lycopene or HG, cells were washed with ice-cold PBS and lysed. The protein concentration was determined by the Bradford method. In brief, 30 µg of total proteins was loaded into each lane of a 9% sodium dodecyl sulfate-polyacrylamide gel, followed by electrophoresis and then transfer to a polyvinylidene difluoride membrane. The blots were probed with antibodies against phospho-p38 MAPK. Horseradish peroxidase-conjugated secondary antibodies were used in conjunction with an ECL chemiluminescence detection system (Amersham, UK). Staining was quantified by scanning densitometry.
Statistical analysis
Data were analyzed using SPSS16.0 software (SPSS Inc., Chicago, IL, USA), and all measurement data were expressed as mean ± standard deviation (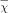 ± SD). Analysis of variance was used for group comparisons. Statistical analysis was performed using analysis of variance for mean comparison of multiple samples, LSD tests for paired comparison, and variance analysis of replicate measurements data for comparison of different experiment intervals. Absolute values, which were equal to differences in functional parameters between the lycopene plus HG group, were subjected to curve fitting analysis. The level of statistical significance for all analyses was set at α = 0.05.
± SD). Analysis of variance was used for group comparisons. Statistical analysis was performed using analysis of variance for mean comparison of multiple samples, LSD tests for paired comparison, and variance analysis of replicate measurements data for comparison of different experiment intervals. Absolute values, which were equal to differences in functional parameters between the lycopene plus HG group, were subjected to curve fitting analysis. The level of statistical significance for all analyses was set at α = 0.05.
± SD). Analysis of variance was used for group comparisons. Statistical analysis was performed using analysis of variance for mean comparison of multiple samples, LSD tests for paired comparison, and variance analysis of replicate measurements data for comparison of different experiment intervals. Absolute values, which were equal to differences in functional parameters between the lycopene plus HG group, were subjected to curve fitting analysis. The level of statistical significance for all analyses was set at α = 0.05.RESULTS
Identification of EPCs
The isolated mononuclear cells formed several cell masses from days 3-4, and some spindle cells sprouted from the edge of the cell mass from days 5-6. By day 7, adherent cells had increased in number, and some aggregated to form a "Blood Island". Laser confocal microscopy showed that Dil-Ac-LDL (red, excitation wavelength 543 nm) and Ulex lectin FITC-I (green, excitation wavelength 477 nm) double-stained positive cells were differentiating EPCs [10] (Fig. 1).
Effects of lycopene on EPC proliferation
As shown in Table 1, dramatic reduction of proliferation was observed following exposure of EPCs to HG for 12, 24, 48, and 72 h compared to the control group (P < 0.05) at all time points during the experiment. In contrast, osmotic control with mannitol had no effect on proliferation of EPCs (P > 0.05). Lycopene significantly increased the proliferative capacity of EPCs compared to that of the HG group. Absorbance values of the three lycopene groups were higher than that of the HG group (P < 0.05). Compared with the control group, the absorbance value of EPCs from the 30 µg/mL lycopene group was the highest, and differences were statistically significant at three time points (12, 24, and 48 h; except 72 h). In the repeated measures analysis of variance, Ftime = 129.241, P = 0.000; F time* group = 24.425, P = 0.000; and Fgroups = 920.137, P = 0.000. Lycopene was the main effect factor responsible for increasing the absorbance value and protecting the proliferative capacity of EPCs cultivated with HG (Table 1).
Effects of lycopene on EPC apoptosis
As shown in Table 2, dramatic elevation of apoptosis was observed following exposure of EPCs to HG for 12, 24, 48, and 72 h compared to the control and mannitol groups (P < 0.05) at all time points during the experiment. Mannitol had no effect on EPC apoptosis (P > 0.05). Lycopene significantly inhibited apoptosis of EPC compared to that of the HG group. Apoptotic rates of EPCs from the three lycopene groups were lower than that of the lycopene plus HG group (P < 0.05). Compared with the control group, the apoptotic rate of EPCs from the 30 µg/mL lycopene group was the lowest, but differences were not statistically significant at any time point. In the repeated measures analysis of variance, Ftime = 189.810, P = 0.000; Ftime * group = 54.118, P = 0.000; and Fgroups = 1741.180, P = 0.000. Lycopene was the main effect factor responsible for inhibiting apoptosis of EPCs cultivated with HG (Table 2).
Effects of lycopene on EPCs migration, adhesion, and in vitro vasculogenesis capacity
As shown in Table 3, dramatic inhibition of migration, adhesion, and in vitro vasculogenesis capacity was observed following exposure of EPCs to HG for 24 h compared to the other groups (P < 0.05). The detrimental effects of HG on EPC function were significantly attenuated by treatment of EPCs with lycopene (10, 30, and 50 µg/mL) for 24 h compared with the 33 mmol/L glucose group (P < 0.05). EPC migration and adhesion were higher in the 30 µg/mL lycopene group compared to the 10 and 50 µg/mL lycopene groups (P < 0.05). In comparing the 30 µg/mL lycopene group with the signal inhibitor group (SB203580), there was no difference in migration, whereas adhesive capacity following lycopene treatment was higher than that of the signal inhibitor group (P < 0.05) (Table 3).
For the Matrigel network formation assay, EPCs were cultured on Matrigel for 24 h to form an extensive and enclosed microvascular network structure. HG impaired the ability of EPCs to form the microvascular network structure. Specifically, the total length of the tube network decreased while the tube network was severely disrupted under HG, resulting in an incomplete and sparse microvascular network (P < 0.05) (Table 3). Lycopene and signal inhibitor restored the ability of EPCs to form the microvascular network in vitro. There were also significant differences in vascular network capacity among the EPC groups according to the dosage level of lycopene. As shown in Table 1, lycopene improved EPC vasculogenesis capacity in the presence of 33 mmol/L of glucose in a dose-dependent manner, and maximal effect was observed at 30 µg/mL of lycopene. Compared with the other lycopene groups (10, 50 µg/mL) and signal inhibitor group, microvascular structures were more complex and complete in EPCs treated with 30 µg/mL of lycopene. Representative pictures of tube formation in EPCs subjected to different treatments are shown (Fig. 2).
Effects of lycopene on MAPK activation in EPCs
To further characterize the mechanism underlying the protective effects of lycopene, activation of p38 MAPK was examined by Western blotting. As showed in Fig. 3, HG sharply increased the phosphorylation of p38 MAPK, and the increase was inhibited by lycopene. Total amounts of p38 MAPK were unaltered by either HG or lycopene. The protective effect observed in the 30 µg/mL lycopene group was greater than that in either the 10 or 50 µg/mL lycopene group. To confirm the causal relationship between MAPK inhibition and the protective effects of lycopene against HG-induced cellular injury, the effects of the MAPK inhibitor SB203580 on HG were further examined. The inhibitor similarly reversed the detrimental effects of HG (Fig. 3). These results indicate that inhibition of MAPKs is responsible for the protective effects of lycopene against HG-induced EPC injury.
Curve fitting analysis of effects of lycopene on EPC function
To explore the effects of lycopene to EPC functional variables, we carried out curve fitting analyses. Dose of lycopene was the independent variable, whereas the absolute number of differences in the various functional parameters (HG group mean-lycopene groups value) was the dependent variable. The regression equation is shown in Fig. 4. Each regression equation's coefficient of determination R2 was satisfactory, P = 0.000. Differences in EPC functional variables were detected within the range of lycopene doses (10-50 µg/mL) (Fig. 4).
DISCUSSION
EPCs are a kind of precursor cell that can differentiate into vascular endothelial cells. EPCs are not only involved in human embryonic angiogenesis but also in postnatal angiogenesis and endothelial repair, which is a major factor in maintaining dynamic equilibrium between endothelial injury and repair. Important risk factors for the occurrence and development of vascular complications due to diabetes mellitus are the number of EPCs and vascular endothelial dysfunction [32]. EPCs from patients with type 2 diabetes mellitus are characterized by reduced proliferative capacity as well as lower adhesiveness and ability to form capillary tubes in vitro [32]. It was recently shown that HG impairs the proliferation and function of early and late EPCs, including migration and capacity for vasculogenesis [14,33].
Previous epidemiological studies have established that lycopene is strongly associated with reduced carotid intima-media thickness and lower incidences of myocardial infarction, ischemic heart disease, stroke, cancer, and diabetes. Intake of lycopene-rich foods can prevent the occurrence and development of these chronic diseases [20,34,35,36,37,38]. Among carotenoids, only lycopene has been shown to modulate adhesion molecule expression in human aortic endothelial cell cultures, which suggests a role in mitigating atherogenesis [21]. Some studies indicated that lycopene exerts a protective vascular endothelial function effect on a variety of endothelial dysfunctional processes [39,40,41]. EPCs can differentiate into vascular endothelial cells. In the present study, we observed the effects of HG on EPC proliferation, apoptosis, and function, as well as the potential mechanisms responsible for these effects. Lycopene has been shown to act as a pro-oxidant or antioxidant depending on its administrated dose [42,43]. In Lowe GM's study [42], lycopene was shown to only provide protection against DNA damage (induced by xanthine/xanthine oxidase) at relatively low concentrations (1-3 µM). However, at higher concentrations (4-10 µM), its ability to protect cells against such oxidative damage was rapidly lost. Another study using Hs68 cells demonstrated that lycopene can be either an antioxidant or pro-oxidant depending on the oxidants used, but it is unclear whether or not lycopene has pro-oxidant activity in vivo [43]. Our study suggests that apoptotic rate and phosphorylation of p38 MAPK are increased in EPCs cultivated with HG, whereas EPC proliferation, migration, adhesion, and in vitro vasculogenesis capacity are attenuated. On the other hand, we found that lycopene inhibits the excessive phosphorylation of p38 MAPK, promotes proliferation, migration, adhesion, and in vitro vasculogenesis capacity, and reduces apoptosis of EPCs induced by HG. The results in Table 3 indicate that the proliferation, migration, adhesion, and in vitro vasculogenesis capacity of EPCs from the 30 µg/mL lycopene group were significantly elevated compared to the HG group and 10 and 50 µg/mL lycopene groups, whereas the apoptotic rate and phosphorylation of p38 MAPK were significantly reduced. Therefore, whether or not the pro-oxidation phenomenon in the above experiments is observed with a higher dose of lycopene must be studied further. This study showed that the proliferation, migration, adhesion, and in vitro vasculogenesis capacity of EPCs were significantly higher in the three lycopene groups compared to the HG group, whereas apoptotic rate and phosphorylation of p38 MAPK were significantly lower. Our results indicate that lycopene can protect proliferative capacity and inhibit apoptosis of EPCs cultivated with HG. Further, lycopene not only can increase the number of EPCs cultivated with HG but also improve their adhesion, migration, and in vitro vasculogenesis capacity. Previous studies have shown that lycopene plays a role in the protection of endothelial cells [39,40,41]. In support of this, our study indicates that lycopene may promote endothelial repair by protecting the proliferative capacity and function of EPCs. HG-induced apoptosis of EPCs has been confirmed by many scholars, and its mechanism of action may promote oxidative stress [44], mitochondria-dependent and non-dependent caspase pathways [45], MAPK pathway, and p53 pathway [46,47].
Previous studies have demonstrated that HG accelerates the onset of EPC senescence via activation of p38 MAPKs [46] and increases oxidative stress [44]. It has been reported that glucose can activate the p38 MAPK pathway in EPCs [35]. Several studies demonstrated that blockage of p38 MAPKs is associated with increased angiogenesis [36,37,38]. Protein phosphorylation, induced by different kinases, plays a crucial role in triggering signal transduction pathways. MAPKs are a group of serine/threonine protein kinases comprising three subfamilies: p42/p44 ERKs, JNKs, and p38 MAPKs. MAPKs can be activated by various extracellular molecules and can induce downstream phosphorylation of many key signaling molecules related to cell proliferation, inflammation, and apoptosis [48]. Studies have shown that p38 MAPK is a critical regulator of EPC differentiation [47]. SB203580 can inhibit HG-induced senescence of EPCs, as well as improve proliferation, differentiation, and in vivo angiogenesis capacity [47]. In our results, proliferation and functional variables of EPCs exposed to HG were significantly reduced compared with the control group, whereas the apoptotic rate significantly increased. This can likely be attributed to the increased phosphorylation of p38 MAPK.
Studies have shown that lycopene has a protective effect against a variety of exogenously-induced endothelial dysfunctional processes [39,40]. Recent studies also showed that lycopene significantly inhibits the effects of lipopolysaccharide (LPS) by suppressing a key inflammatory pathway related to MAPK, NF-κB, NO formation, and IL-6 expression in macrophages, and it has an anti-inflammatory effect [22]. Many independent lines of evidence suggest that different MAPKs can be activated in EPCs in response to HG [46,14]. Such results were confirmed in the present study. Specifically, lycopene significantly inhibited the phosphorylation or activation of p38 MAPK by HG, and total protein levels of MAPK remained unchanged. Therefore, lycopene blocked the activation but not the biosynthesis of MAPKs. Furthermore, we studied the effects of SB203580 on EPC functional variables. EPCs were treated with SB203580, a specific p38 kinase inhibitor. SB203580 reversed the inhibitory effects of HG on EPC function. These results suggest that the inhibitory effect of HG on EPCs is partially mediated through the p38 pathway.
To confirm the causal relationship between MAPK inhibition and the protective effects of lycopene against HG-induced cellular injury, we treated cells with SB203580. The inhibitor significantly inhibited the toxic effects of HG, and EPC adhesion, migration, and angiogenesis in vitro were markedly elevated. Therefore, p38 MAPK activation plays an important role in HG-induced EPC dysfunction, and lycopene can protect EPC function by blocking the activation of p38 MAPK.
In our experiment, there was no significant difference between migration of EPCs from the 30 µg/mL lycopene group and the SB203580 group (P > 0.05). Adhesion and in vitro angiogenesis capabilities of EPCs from the 30 µg/mL lycopene group were superior compared to the SB203580 group. Moreover, the functional variables of EPCs from the 30 µg/mL lycopene group were higher than those of the 10 and 50 µg/mL groups. The exact molecular mechanisms responsible for the effects of lycopene on EPCs remain to be elucidated. Curve fitting (Fig. 4) and the regression equation demonstrated the effects of lycopene on the function of human peripheral blood EPCs cultivated with 10-50 µg/mL of HG, and the data provide a scientific basis for future experimental research.
In conclusion, a high concentration of glucose may attenuate the proliferation, migration, adhesion, and in vitro vasculogenesis capacity of EPCs while increasing the apoptotic rate. On the other hand, lycopene may promote proliferation, migratory, adhesion, and in vitro vasculogenesis capacity as well as reduce apoptosis of EPCs cultivated with HG. These findings suggest that lycopene can improve the function of EPCs incubated with a high concentration of glucose by inhibiting the p38 MAPK pathway, which is one of the mechanisms responsible for protecting EPCs. The underlying molecular mechanisms of the p38 MAPK signal transduction pathway may involve the protective effects of lycopene against HG-induced EPC injury. Further, lycopene may be considered as a new strategy for preventing the macrovascular complications of type 2 diabetes.







 XML Download
XML Download