

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Reactive oxygen species (ROS)-derived oxidative stress plays an important role in the pathogenesis of various diseases, including cancer, cardiovascular diseases, and inflammation [1]. Endogenous antioxidants have evolved to meet the very basic requirement of overcoming the oxidative stresses that occur during normal metabolic processes. However, when the endogenous antioxidant defense systems are overwhelmed by ROS, oxidative stress occurs and ultimately leads to development of various pathological conditions [2,3]. Antioxidant systems, such as antioxidant vitamins, superoxide dismutase (SOD), catalase, glutathione (GSH), and glutathione peroxidase (GSH-Px) protect cells against lipid peroxidation, which is the basis of many pathologic processes [4]. Erythrocytes and liver contain enzymes that contribute to the anti-oxidant defense mechanism. Erythrocytes are constantly exposed to both extracellular and intracellular sources of reactive oxygen species (ROS). In addition, there is a high probability that the liver will be subjected to oxidative stress-induced toxicity [5] because it is an important organ for the biotransformation and detoxification of toxic substances and oxidative agents [6]. Phase I and II biotransformation enzymes in the liver are involved in metabolic activation and detoxification of various toxic xenobiotics. Phase I enzymes convert xenobiotics to active intermediates, and phase II enzymes catalyze the conjugation of these active intermediates with endogenous cofactors and facilitate their excretion [7]. In particular, the induction of phase II detoxifying/antioxidant enzymes, such as glutathione S-transferase (GST), glutamate-cysteine ligase (GCL), which includes a catalytic subunit (GCLC) and a modifier subunit (GCLM), and heme oxygenase-1 (HO-1), results in detoxification of xenobiotics and inactivation of ROS [8].
Tert-butylhydroperoxide (t-BHP), a cytotoxin and oxidative agent, has often been used as a model in investigation of the mechanism of hepatocyte injury initiated by acute oxidative stress. T-BHP can be metabolized to free radical intermediates, such as peroxyl and alkoxyl radicals, by cytochrome P 450 in hepatocytes [9]. In addition, the detoxification of t-BHP can be achieved by GSH-mediated GSH-Px catalyzed reactions to yield t-butanol and glutathione disulfide (GSSG) [10]. GSSG is then reduced to GSH by glutathione reductase (GR). Free radical intermediates induced by t-BHP can subsequently initiate lipid peroxidation and GSH depletion and can lead to oxidative-induced hepatocyte damage [11]. Under oxidative stress, the intake of exogenous antioxidant compounds from a dietary source that enhances the biological antioxidant defense systems can prevent oxidative-induced liver damage [12].
Schisandra chinensis Baillon, a member of the Magnoliaceae family, grows wild in Korea, Russia, China, and Japan [13]. In a previous study, in vitro antioxidant activity of Schisandra chinensis Baillon extracts (SCE) was demonstrated through chemical assays [14]. In addition, its various dibenzocyclooctadiene lignans, including schisandrin, schisandrin B, and gomisin A, have been shown to exert antioxidant activities and hepatoprotective effects in vitro [15] and in vivo [16,17]. This study investigated the endogenous antioxidant capacity of SCE as an exogenous antioxidant dietary source against t-BHP-induced oxidative hepatic damage in rats.
MATERIALS AND METHODS
Plant materials
Schisandra chinensis Baillon (Mungyeong, Gyeongbuk, Korea) was provided by the Rural Development Administration (Suwon, Gyeonggi, Korea). The fruits of Schisandra chinensis Baillon (100 g) were extracted with 1 L of 60% ethanol at 60℃ for 6 h. After removal of the cake by filtration, the extract was freeze-dried under vacuum for five days. The manufacturing yield and the water contents were 32.4% and 10.75%, respectively. SCE was analyzed for schisandrin, Gomisin N, Gomisin A, and Deoxyschisandrin, and such. The content of schisandrin (8.3 mg per 100 g SCE) as a marker compound of the Schisandra chinensis Baillon extract was analyzed by HPLC (Waters Alliance HPLC Systems, USA) using a Luna C18 column (4.5 × 250 mm, 5 µm, Phenomenex Inc., Torrance, CA, USA).
Animals
Seven-week-old male Sprague-Dawley rats (Orientbio Inc., Seongnam, Gyeonggi, Korea) were individually housed in stainless steel wire-mesh cages under laboratory conditions of 23 ± 1℃ with a 12 h light / 12 h dark cycle and 45 ± 5% humidity. Rats (200-220 g, n = 60) were fed a commercial pellet diet (Samyang, Seoul, Korea) for seven days for acclimatization and randomized into six groups: NC, normal control; TC, t-BHP control; SCE-L, 300 mg/kg BW SCE; SCE-M, 600 mg/kg BW SCE; and SCE-H, 1,200 mg/kg BW SCE. Prior to t-BHP injection, the animals received daily administration of SCE by gavage for 14 days. On day 14, t-BHP was injected intraperitoneally (i.p., 0.8 mmol/kg B.W.) into each animal, except for animals in the NC group. After 16 hours, the animals were anesthetized, and their blood was collected by cardiac puncture. The blood samples were centrifuged (3000 × g, 4℃, 30 min) in order to separate the serum. Erythrocytes were separated from the pellet after separation of the plasma layer and stored at -80℃. The liver, kidney, and spleen were removed, weighed, snap-frozen in liquid nitrogen, and stored at -80℃. The experimental protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of the Ewha Womans University of Korea (Approval No. 2012-01-033).
Serum AST and ALT levels
Serum aspartate aminotransferase (AST) and alanine aminotransferase (ALT) activities were measured using commercial kits (Asan Pharmaceutical, Seoul, Korea) based on the method developed by Reitman-Frankel. Absorbance was determined using a microplate reader (Eon Microplate Spectrophotometer, BioTek Instruments, Inc, Winooski, Vermont, USA) at 505 nm.
Histological analysis
The formalin-fixed liver tissue samples were dehydrated through a gradient series (70-100%) of ethyl alcohol, dealcoholized in xylene, embedded in paraffin, and sectioned (5 µm thickness). For histological analysis, the slide-mounted embedded tissues were deparaffinized in xylene, rehydrated in a reverse-gradient series of ethyl alcohol, and stained with hematoxylin and eosin (H&E). The dehydrated and dealcoholized formalin-fixed tissue sections were mounted on slides with Canada balsam, and tissue damage was observed using an Olympus 51 microscope (Olympus Optical Co., Tokyo, Japan).
Quantitative reverse transcription polymerase chain reaction analysis
Total RNA was extracted from the liver samples using TRIzol (Invitrogen Co., San Diego, CA, USA). The RNA concentration and quality were determined using a BioSpec-nano spectrophotometer (Shimadzu Corp., Kyoto, Japan). cDNA was synthesized using a High-Capacity RNA-to-cDNA kit (Applied Biosystems, Foster City, CA, USA). Quantitative RT-PCR was performed using the TaqMan method with the Step-One-Plus RT-PCR System (Applied Biosystems). The primer sets for the target genes were Heme Oxygenase-1 (HO-1, Rn01536933_m1), glutathione S-transferase (GST, Rn01446656_m1), glutamate-cysteine ligase catalytic subunit (GCLC, Rn00689046_m1), glutamate-cysteine ligase modifier subunit (GCLM, Rn00568900_m1), and β-actin (Rn0066 7869_m1). Amplifications were performed starting with a 10-min template denaturation step at 95℃, which was followed by 40 cycles at 95℃ for 15 s and 60℃ for 1 min. The relative amounts of these mRNAs were normalized to the amount of β-actin, and the relative amounts of the RNAs were calculated using the comparative CT method.
Erythrocyte antioxidant enzyme activity
The superoxide dismutase (SOD) activities of the erythrocytes were determined using the method developed by Miquel [18]. One unit of SOD was defined as the amount of enzyme that inhibits 50% of cytochrome c in a xanthine-xanthine oxidase system. Erythrocyte catalase (CAT) activity was measured by production of formaldehyde from the decomposition of H2O2, as described by Johnson et al. [19].
Hepatic glutathione level and glutathione reductase activity
Total glutathione (GSH) concentrations were quantified using the glutathione reductase recycling assay as described by Akerboom et al. [20]. Glutathione Reductase (GR) activity was assayed as described by Foyer et al. with some modifications [21]. GR activity was expressed as nmoles of NADPH per minute per milligram of protein using the following equation, with a molar extinction coefficient for NADPH at 340 nm. Glutathione peroxidase (GSH-Px) activity was determined using the modified method of Mohandas et al. [22]. GSH-Px activity was expressed as nmoles of NADPH oxidized to NADP per minute per milligram of protein using the following equation, with a molar extinction coefficient for NADPH at 340 nm.
Plasma TBARS levels
Plasma malondialdehyde (MDA) was estimated using the method of thiobarbituric acid reactive substances (TBARS), which primarily reflects malondialdehyde (MDA), as previously described [23]. The absorbance of the organic layer was measured at 535 and 572 nm using a microplate reader (Eon Microplate Spectrophotometer, BioTek® Instruments, Inc, Winooski, Vermont, USA).
Statistical analysis
All statistical analyses were performed using the SAS 9.2 program (SAS Institute, Cary, NY, USA). The values are presented as the mean and standard error (S.E.). The significance of the differences between the NC and TC groups was analyzed using the Student's t-test (**P ≤ 0.01). The significance of the differences between the TC and SCEs groups was analyzed using one-way analysis of variance (ANOVA) with post-hoc Dunnett's multiple-comparison test (#P ≤ 0.05 and ##P ≤ 0.01).
RESULTS
Effects of SCE on serum ALT and AST levels in rats with t-BHP-induced hepatic damage
Significantly higher serum ALT activity was observed in the TC group compared with that in the NC group (68.94 ± 12.71 vs. 10.44 ± 0.57, P = 0.0013, Fig. 1A). No significant differences in the levels of ALT were observed in the SCE groups compared with the TC group, however, that of the SCE-H group was lowered compared to the TC group. A significant increase in serum AST activity was observed in the TC group compared to the NC group (206.15 ± 41.03 vs. 29.86 ± 1.83, P = 0.002, Fig. 1B). Although no significant differences were observed among the TC and SCE groups, that of the SCE-H group was lowered compared to the TC group.
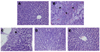
Effects of SCE on hepatic morphology in rats with t-BHP-induced hepatic damage
Livers of the NC group showed a regular arrangement of hepatocytes with clearly visible nuclei (Fig. 2A). However, those of the TC group showed moderate neutrophil infiltration (sharp arrow), swelling (arrow), and necrosis (arrowhead, Fig. 2B). Swelling and necrosis of hepatocytes in the SCE-L group were decreased compared with the TC group (Fig. 2C). The SCE-M and SCE-H groups showed only swelling and almost normal morphology (Fig. 2D and 2E). Pretreatment with SCE resulted in reduced incidence of lesions, such as neutrophil infiltration, swelling of hepatocytes, and necrosis, induced by injection of t-BHP.
Effects of SCE on GST, GCLC, and GCLM mRNA expression levels in rats with t-BHP-induced hepatic damage
Hepatic HO-1, GST, GCLC, and GCLM mRNA levels were measured using quantitative RT-PCR in order to determine the effects of SCE on phase II detoxification and antioxidant enzymes (Fig. 3). HO-1 expression in the TC group tended to be higher than that observed in the NC group. However, a marked decrease was observed in the SCE-L and SCE-M groups compared with the TC group, although no significant differences were observed among the groups (Fig. 3A). The mRNA levels of GST, GCLC, and GCLM were lowered significantly by t-BHP injection compared to the NC group (P = 0.0006, P < 0.0001, and P = 0.046, respectively).
Conversely, the GST, GCLC, and GCLM mRNA levels in the SCE-pretreated groups showed a dose-dependent increase with SCE (Fig. 3B, 3C and 3D). In particular, significantly higher mRNA levels of GST and GCLC were observed in the SCE-H group than in the TC group (P = 0.0177 and P = 0.0097, respectively). No significant difference in the GCLM mRNA level was observed between the TC and SCEs groups, although the SCE-H group showed an increasing tendency compared to the levels observed in the TC group.
Effects of SCE on antioxidant enzyme activities in rats with t-BHP-induced hepatic damage
In erythrocytes, the SOD activity was lowered significantly in the TC group compared with the NC group (15.60 ± 2.40 vs. 9.97 ± 0.83, P = 0.0483, Table 1). The SCE-M group tended to exhibit higher SOD activity than the TC group, and the SCE-L group showed a significant increase in SOD activity compared with the TC group (9.97 ± 0.83 vs. 16.82 ± 1.77, P < 0.0001). Nonetheless, no significant difference in catalase activity was observed among the groups. In the liver, the total GSH and GR activities did not differ significantly among the groups. In addition, no significant difference in plasma TBARS level and hepatic GSH-Px activity was observed among the groups.
DISCUSSION
We evaluated the endogenous antioxidant capacity and hepatoprotective potential of SCE against t-BHP-induced oxidative liver damage in rats. T-BHP induces lipid peroxidation, damages the membrane of liver cells and organelles, causes swelling and necrosis of hepatocytes, and results in release of cytosolic enzymes, such as ALT and AST, into the circulating blood [24]. In the current study, the serum ALT and AST activities tended to decrease after pretreatment of rats with a high dose of SCE. In addition, SCE caused attenuation of liver lesions in rats with t-BHP-induced oxidative damage. The hepatic histopathological evaluation revealed that pretreatment with SCE resulted in reduced t-BHP-induced incidence of lesions, such as neutrophil infiltration, hepatocyte swelling, and necrosis.
The body has an effective mechanism for prevention and neutralization of the damage induced by free radicals. This mechanism is accomplished by a set of endogenous antioxidant enzymes, including SOD, catalase, GSH, and GSH-Px. SOD neutralizes the superoxide anion that plays a pivotal role in oxidative stress-induced cellular damage by transforming it into hydrogen peroxide and thereby preventing formation of highly aggressive compounds, such as peroxynitrite and hydroxyl radical [26]. Erythrocytes are extremely vulnerable to these oxidative challenges [25]. SOD and CAT are considered to be a primary defense that protects biological macromolecules from oxidative damage. Intake of exogenous antioxidant compounds from a dietary source that enhances the biological antioxidant systems can prevent oxidative damage [12]. In this study, pretreatment with a low dose of SCE prevented the decrease in the erythrocyte SOD activity induced by t-BHP. However, catalase activities did not differ significantly among the groups. Next, we focused on hepatic phase II antioxidant/detoxifying enzymes because we used a t-BHP-induced oxidative hepatic damage model. Liver is known to undergo free radical-mediated injury, and increased ROS is related to damage of hepatic anti-oxidant defense. Antioxidants are of direct and indirect types. Direct antioxidants, such as GSH and ascorbic acid, are substances that can participate in physiological, biochemical, and cellular processes involved in inactivation of free radicals and/or that prevent free radical-initiated chemical reactions. In contrast, indirect antioxidants boost the antioxidant capacity of cells and thereby afford protection against oxidative stress. There is substantial evidence indicating that phase II enzymes, such as GSTs, NAD(P)H, and heme oxygenase, play important roles in detoxification of electrophiles and that their induction protects animals and their cells against carcinogenesis and mutagenesis [27,28]. Induction of phase II detoxifying/antioxidant enzymes, such as GST, GCLC, GCLM, and HO-1, results in detoxification of xenobiotics and inactivation of ROS [8]. In particular, GST catalyzes the conjugation of electrophilic molecules with GSH and thereby protects cellular macromolecules against toxic foreign chemicals and oxidative stress [7]. In addition, GCL is the rate-limiting enzyme in the overall glutathione synthesis pathway. In this study, no significant differences in hepatic total glutathione (GSH) level, glutathione reductase (GR), and glutathione peroxidase (GSH-Px) activities were observed among the TC and SCE groups. On the other hand, treatment with a high dose of SCE resulted in induction of phase II antioxidant/detoxifying enzyme expression, such as glutathione S-transferase (GST) and glutamate-cysteine ligase catalytic subunit (GCLC). The major bioactive components of SCE include schisandrin, Gomisin N, Gomisin A, and Deoxyschisandrin, and such. Among them, schisandrin is the highest, at 8.3 mg per 100 g SCE. Previous studies have demonstrated that schisandrin exhibits anti-oxidative effects in vivo [16]. In particular, the methylenedioxy group of the schisandrin molecule is an important structural determinant for enhancing the hepatic mitochondrial glutathione and hence protecting against CCl4 hepatotoxicity in mice [29]. LD50 value of ethanol extract of Schisandra chinensis was estimated to be 35.63 ± 6.46 g/kg [30]. In addition, Ryu et al. investigated acute toxicity of the fruit figment of SCE in mice following oral administration at the doses of 1, 2, and 4 g/kg, without abnormal behaviors, body weight, or other toxic symptoms for 14 days [31]. Furthermore, the activities of AST and ALT were decreased by 92% and 87%, respectively, by administration of 5 g SCE/kg BW in rats with CCl4-induced liver injury [32]. In addition, 1.2 g/kg as an animal dose is applicable to 0.72 g/day as a human dose, depending on the safety factor. Therefore, in this study, medium or high-doses of SCE were free from undesirable side effects.
In conclusion, we suggest that SCE is a potent inducer of phase II detoxification enzymes and exerts protective effects against t-BHP-induced oxidative hepatic damage. In particular, treatment with SCE resulted in reduced neutrophil infiltration, swelling of liver cells, and necrosis in liver. Induction of phase II antioxidant and detoxifying enzymes such as GST, GCLC, and GCLM mRNA expression might contribute to antioxidant potential of SCE via detoxification of xenobiotics and inactivation of ROS. However, SCE regulates gene expression of phase II antioxidant/detoxifying enzymes independent of hepatic antioxidant enzyme activity. Conduct of further studies will be needed in order to provide a logical explanation for interaction between these parameters.



 XML Download
XML Download