

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Persistent hypertension increases the risk of cardiovascular disease, including stroke, myocardial infarction, and heart failure, renal disease, and mortality. In particular, in individuals older than 50 years, systolic blood pressure (SBP) greater than 140 mmHg rather than diastolic blood pressure is a major risk factor for cardiovascular disease [1,2].
Reactive oxygen species (ROS) are associated with many vascular risk factors, including hypertension. Antioxidant properties of vegetables reduce the risk of cardiovascular disease [3]. ROS are scavenged by enzymes such as glutathione peroxidase (GPx) and catalase, which convert hydrogen peroxide into water [4]. The L-arginine-NO pathway also plays an important role in hypertension. Endothelial nitric oxide synthase catalyzes the production of nitric oxide (NO) from L-arginine [5]. In addition to its role in protein synthesis, L-arginine is essential in the synthesis of creatinine, urea, and NO. NO diffuses across the endothelial cell into neighboring smooth muscle and induces vasodilation. Impaired endothelium-dependent vasodilation due to alteration in the L-arginine-NO pathway is a common characteristic of hypertension [6]. While a loss of NO bioactivity in the vessel wall impairs endothelium-dependent vasorelaxation, an increased serum NO level helps ameliorate hypertension [7]. The role of oxidative stress in the pathogenesis of hypertension, at least in part, is related to endothelial dysfunction due to NO degradation by oxygen free radicals [8,9].
A fundamental pathogenic feature of hypertension is angiotensin-converting enzyme (ACE) activity. In the renin-angiotensin-aldosterone system (RAAS), ACE converts the inactive decapeptide angiotensin-I by cleaving a dipeptide from the C-terminus to form the potent vasoconstricting octapeptide angiotensin II. This potent vasopressor is also involved in the release of a sodium-retaining steroid aldosterone, which suppresses the excretion of Na+ in urine, from the adrenal cortex. In the kallikrein-kinin system, ACE inactivates bradykinin, a vasodilator [10]. Several studies have reported that the restoration of endothelial function in hypertensive patients by the administration of ACE inhibitors [11,12]. The treatement of ACE inhibitor reduced 24 hour ambulatory blood pressure. Wing et al. [13] reported that the initiation of antihypertensive treatment with ACE inhibitors in elderly led to better outcomes than the treatment with diuretic agent. Therefore, ACE inhibitors are widely used for the treatment of hypertension and congestive heart failure [14].
Recently, many reports regarding the antihypertensive effects of polyphenol-rich foods have been published. Several epidemiological studies have shown an association between high consumption of chocolate, berries, and grape seeds and low blood pressure and low risk of cardiovascular disease [15-19]. Other studies carried out in humans have also shown improved endothelial function after consumption of cocoa, wine, and tea [20,21]. These results indicate that polyphenol-rich foods decrease arterial pressure probably by an antioxidant mechanism.
Demand for medicinal foods for alleviating hypertension and cardiovascular disease has increased. Readily available alternative potency with minimal side effects is preferred. Radish (Raphanus sativus L.) belongs to the cruciferae family, which includes white and red cabbage, broccoli, brussels sprouts, and cauliflower [22]. It contains glucosinolates, their derivatives, including isothiocyanates and nitriles, essential oils, flavonoids, and other polyphenol compounds [22]. In particular, glucosinolates in the family Brassicaceae (syn. Cruciferae) have been found to have antioxidant effects and anticancer properties [23]. Glucosinolates suppress lipid peroxide production and protect against DNA damage [24]. In addition, they might exert an antihypertensive effect by mechanisms associated with antioxidation.
Hypertension in SHRs was suppressed after 4 weeks of feeding with radish leaf powder. We also confirmed the inhibitory activity of radish leaf powder for ACE measured in a previous study [25]. The identification of the presence of potential antihypertensive agents in routinely consumed radish leaf generated much scientific interest because it fulfilled the demand for an ideal and efficacious antihypertensive agent. However, a detailed study of the antihypertensive activity of radish leaf extract is yet to be conducted. Therefore, the objective of this study was to investigate the antihypertensive activity of an ethyl acetate fraction of an ethanol-based extract obtained from radish leaves in SHRs.
Materials and Methods
Preparation of the leaf extract by solvent
Radish leaves were collected in September 2010 from Dongsu Farm, Yangpeong-gun, Korea. The leaves were completely dried at 50℃ and were first extracted with 80% ethyl alcohol at room temperature for 24 h. The alcohol extract was filtered using Whatman #1 filter paper, and the remaining material was extracted twice. The alcohol extract was evaporated at 45℃ by using Speed Vac concentrator (Savant Instruments, Farmingdale, NY) and suspended in distilled water and n-hexane (1:1, v/v). It was further extracted sequentially with methylene chloride and ethyl acetate. The resulting extracts were evaporated to dryness in vacuum to obtain fractions. We obtained 18.0 g of ethyl acetate extract from 1 kg of radish leaves. The soluble ethyl acetate extract was selected for use as a supplement for SHRs, because it showed the highest inhibitory activity in the subsequently performed ACE assay in vitro (data not shown).
Experimental animals and diets
Eighteen 11-week-old male SHRs with SBP greater than 200 mmHg, and 6 age-matched normotensive male Wistar rats were purchased from Central Lab. Animal Inc. (Seoul, South Korea). The rats were fed a non-purified chew diet, which was obtained from Cargill Agri Purina (Minnesota, USA). The rats were housed in individual wire cages in a controlled environment with a temperature of 22 ± 1℃, relative humidity of 65%± 5%, and a 12-h light-dark cycle (the lights were switched on at 6 pm). After a week of acclimation, the SHRs were randomly divided into 3 groups of 6 rats each on the basis of their initial blood pressure. The rats were treated with oral administration of ethyl acetate extract of radish leaves for 5 weeks as follows: normotensive control Wistar rats administered 0.3 mL of the vehicle per os (p.o.) (WC), hypertensive control SHRs administered 0.3 mL of the vehicle (SC), hypertensive rats with a low dose of radish leaf extract at 30 mg/kg bw (SL), and hypertensive rats with a high dose of radish leaf extract at 90 mg/kg bw (SH). The powdered extract was dissolved with DMSO, and the concentration of DMSO was 50%, v/v. The lower p.o. dose level was chosen because it is approximately equivalent to a daily dose of radish leaf when received as a part of the diet (1%) and this concentration has been used frequently in intervention studies [26]. All aspects of this study were conducted according to the standards of the Animal Ethics Committee of Kookmin University.
Measurement of blood pressure
Blood pressure was measured indirectly and non-invasively every week by using the volume-based tail-cuff method involving the use of a sphygmomanometer (LE5001, Panlab, Spain). To get accurate blood pressure, at least 5 consecutive determinations were recorded, then, the maximum and minimum values were eliminated, and blood pressure was calculated as the average of the remaining 3 values.
Collection of urinary and fecal excretions, blood, and tissues
In the 5th week, to collect excretion samples from the rats, the rats were kept in metabolic cages for 48 h. After 16 h of fasting, animals were killed by decapitation. Blood samples were collected and centrifuged at 3,000 rpm at 4℃ for 15 min. The supernatants of blood and red blood cells (RBCs) were stored at -70℃ until analysis. The RBCs were separated from platelet-rich plasma by centrifugation (180 g, 20 min, 22℃) and washed thrice with a saline buffer. Then, the liver, kidney, and lung were removed, weighed, and stored at -70℃ for various analyses.
Analysis of ACE activity
Serum ACE level was determined according to the method of Hurst and Lovell-Smith [27]. A mixture of 0.2 mL of incubation buffer (containing 0.2 M boric acid and 2 M NaCl, pH 8.3), 0.1 mL of distilled water, and 15 µL of serum sample were placed and incubated at 37℃ for 5 min. The above mixture was added with 0.1 mL of 12.5 mM hippuryl-L-histidyl-L-leucine (HHL; Sigma Chemical Co, USA) into borate buffer (containing 0.05 M borax and 0.2 M boric acid, pH 8.3) as the substrate and incubated at 37℃ for 15 min. The reaction was stopped by adding 0.5 mL of 1 M HCl and neutralized by adding 0.5 mL of 1 M NaOH. We added 2 mL of diluent buffer (0.2 M KH2PO4, pH 8.3), followed by 1.5 mL of color reagent (containing 30 g of cyanuric chloride in 1 L of 1,4-dioxan). The mixture was centrifuged at 3,000 rpm for 10 min. The absorbance of the supernatant was measured at 382 nm with a spectrophotometer (Ultrospec 2100 pro, Amersham, USA). For determining the ACE activity in the tissues, 100 mg of lung were homogenized with 5 mL of 100 mM borate buffer (containing borax, pH 8.3), and 500 mg of kidney were homogenized with 2 mL of 100 mM borate buffer.
Analysis of nitric oxide (NO)
Because of the short half-life and low concentrations of NO in vivo [28], we evaluated serum NO levels by measuring its stable metabolites, nitrite (NO2-) and nitrate (NO3-), by the modified Griess reaction method [29]. The principle of the assay was reduction of nitrate by vanadium (III) combined with detection by the acidic Griess reaction. After loading a 96-well plate with 50 µL of serum and standard, respectively, 80 µL of vanadium (III) chloride (VCl3; containing 400 mg of VCI3 in 50 mL of 1 M HCL) was added to each well and rapidly followed by addition of 20 µL of Griess reagents, sulfanilamide (SULF; 2% in 5% HCl), and N-(L-naphthyl) ethylenediamine dihydrochloride (NEDD; 0.1%). Nitrite was measured in a similar manner except that samples and nitrite standards were exposed to Griess reagents only. Samples were allowed to react for 45 min at 37℃, and then measured at 540 nm with an enzyme-linked immunosorbent assay reader (UVM 340 & MicroWin S/W, Seoul in Bioscience, Korea).
Analysis of glutathione peroxidase activity
Glutathione peroxidase (GPx) activity in red blood cells (RBCs) and tissue homogenates was analyzed using Glutathione Peroxidase Cellular Activity Assay Kit (CGP1, Sigma, USA). RBCs were washed with 0.9% NaCl and diluted 11 times with 400 mM phosphate buffer (containing 340 mM sucrose, 900 mM NaCl, and 10 µL of protease inhibitor cocktail per 1 mL of homogenate, pH 7.2). Liver was homogenized with the phosphate buffer, and the homogenates were centrifuged at 3,000 rpm for 10 min, and the supernatants were used for analysis.
Analysis of catalase activity
Catalase activity was analyzed spectrophotometrically as described by Beers and Sizer [30]. From the liver, kidney, and RBCs each, 0.4 g was homogenized with 1.6 mL of 50 mM phosphate buffer (pH 7.0). The homogenates were centrifuged at 10,000 rpm for 30 min, and then 2 mL of the supernatant was allowed to react with 1 mL of 30% hydrogen peroxide (H2O2). The decrease of 30% H2O2 in the presence of the tissue homogenate was measured at 240 nm after 2 min. Reaction mixtures with 1 mL of 50 mM phosphate buffer (pH 7.0) instead of H2O2 were used as tissue blanks. Tissue catalase activity was calculated as follows:
CAT activity (U/mg) = ΔA/43.6 × 3/min/mg protein
ΔA = absorbance of the test-the blank
3 = total assay volume
43.6 = millimolar absorptivity of the chromogen
Analysis of lipid peroxide
Malondialdehyde (MDA) is a reactive substance of thiobarbituric acid (TBA). The level of lipid peroxide in the liver, kidney, and RBCs was determined by measuring concentrations of TBA-MDA complex. Tissues homogenates diluted 5-fold with 0.1 mM phosphate buffer (pH 7.4), 1.5 mL of 0.8% TBA, 0.2 mL of 8% sodium dodecyl sulfate (SDS), 1.5 mL of 20% acetic acid, and 0.7 mL of distilled water were combined, and the suspension was boiled for 1 h. After cooling, 4 mL of the mixture of butyl alcohol and pyridine (14:1) was added. This mixture was mixed and centrifuged at 3,000 rpm for 10 min. Then, absorbance was measured at 532 nm [31]. The protein content of the tissues was quantified using the SMART BCA Protein Assay Kit (21071, iNtRON, Korea).
Analysis of Na+ and K+ concentrations
After 25 µL of serum was diluted to 1.5 mL by adding 0.01 N HCl, the sample was measured with inductively coupled plasma-mass spectrometry (ICP-MS; Waters, USA). Urine was filtered using ashless Whatman #40 filter paper and diluted with distilled water depending on the respective volume. The concentration of the diluted urine was adjusted by adding 0.01 N HCl, and then the sample was measured with inductively coupled plasma-mass spectrometry. Feces were desiccated at 10 5℃ until a constant weight was achieved; then, the feces (0.3 g) were burned in a furnace (temperature, 550-600℃) for 15 h. The ash was dissolved in HCl for 15 h and filtered. After 300-fold dilution with 0.01 N HCl, the sample was analyzed with inductively coupled plasma-mass spectrometry.
Statistical analysis
The data are expressed as means ± standard error of the means. All data were analyzed using SPSS ver. 18.0 (SPSS, Inc., Chicago, IL, USA). One-way analysis of variance (ANOVA) was performed, and Duncan's multiple range test was used to determine which mean values were significantly different (P < 0.05).
Results
Body weight gain
Body weight gain of the groups is presented in Table 1. There was no significant difference among the groups. The daily food intake in the metabolic cages was also not significantly different among the groups. Ethyl acetate extract of radish leaves had no effect on food intake and body weight.
Systolic blood pressure
Changes in SBP during the experimental period are shown in Fig. 1. The SBP of the normotensive WC group was constant while that of all the spontaneously hypertensive groups showed a decreasing tendency. However, all of the spontaneously hypertensive groups-SC, SL, and SH-showed significantly higher SBP than the normotensive WC group for the 5 weeks. The SL group presented a decreasing slope (SBP = -8.399x + 224.6, r2 = 0.89), which is meaningful. In the third week, the SBP of the SH group fed with 90 mg extract/kg bw reduced from 214 mmHg to 166 mmHg and was significantly lower than that of the WC and SC groups.
Angiotensin-converting enzyme activity
The results of ACE activity analysis after feeding for 5 weeks are shown in Fig. 2. Ethyl acetate extract of radish leaves did not show a significant effect on the ACE activity in the serum, kidney, or lung, although the activity in the kidney tended to be enhanced and that in the lung tended to be reduced.
Nitric oxide
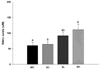
The serum NO measurements are presented in Fig. 3. The serum NO concentration in the SH group was significantly higher than that in the WC and SC groups. This indicated that the oral administration of ethyl acetate extract of radish leaves increased the serum NO concentration.
Antioxidant enzyme activity and lipid peroxide content
Antioxidant enzyme activities and lipid peroxide content are shown in Table 2. We examined the activities of glutathione peroxidase (GPx) and catalase as antioxidant enzymes and lipid peroxide on the basis of malondialdehyde (MDA) level. Intergroup differences in GPx activity in the liver were not significant, and the GPx activity in the RBCs was higher in the groups treated with the ethyl acetate extract of radish leaves than in the normotensive and hypertensive controls. There were no intergroup differences in catalase activity in the liver and kidney, however, intergroup differences in catalase activity in the RBCs were significant. The catalase activity in the normotensive WC group was significantly higher than that in the spontaneously hypertensive SC group. In addition, the catalase activity in the hypertensive SL group that was administered a low dose of 30 mg/kg bw radish leaf extract was significantly higher than that in the SC group. The activity in the SL group was almost as high as that in the normotensive control SH group. No intergroup differences were noted in MDA levels in the liver, kidney, and RBCs.
Na+ and K+ concentration in serum, urine, and feces
Na+ and K+ concentrations in serum, urine, and feces are given in Table 3. Fecal concentrations of Na+ and K+ were significantly higher in the SL and SH groups than in the WC and SC groups. This indicates that the ethyl acetate extract of radish leaves increased the fecal concentration of Na+ and K+. Intergroup differences in serum Na+ and K+ concentrations were not significant. However, urinary concentration of Na+ in the normotensive Wistar rats was higher than that in the SHRs, while that of K+ was not significantly different.
Discussion
We reported that radish leaf powder suppressed hypertension in SHRs and inhibited ACE activity in previous study [25]. In the present study, we intended to examine whether an ethyl acetate extract of radish leaves can decrease blood pressure in SHRs and whether it has antioxidant properties.
The administration of the ethyl acetate extract of radish leaves did not affect weight gain or food intake. The dose of 30 mg/kg bw was equivalent to general consumption of radish leaves in the diet (1%) and was deemed reasonable.
It has been reported that antioxidant effects of the extract delay the development of hypertension. Both superoxide anion and hydrogen peroxide production by polymorphonuclear leukocytes has been observed, and the plasma levels of lipid peroxides are higher in individuals with uncontrolled essential hypertension than in normal controls [32]. The concentrations of antioxidants such as vitamin E and superoxide dismutase were found to be decreased in patients with uncontrolled hypertension [33]. These results suggest that hypertension is characterized by an increase in free radical generation and a simultaneous decrease in the production of nitric oxide and anti-oxidants. This increase in free radical generation can inactivate prostacyclin and nitric oxide and decrease their half-life, which can lead to an increase in peripheral vascular resistance and hypertension.
Several experimental and clinical trials have linked enhanced production of ROS to certain diseases of the cardiovascular system, including hypertension. ROS include superoxide anion radical (O2-), hydrogen peroxide (H2O2), hydroxyl radical (OH), and nitric oxide (NO) is involved in the control of vascular function [3]. In recent studies, production of excessive O2- in the vascular wall has been observed in animal models of hypertension, including SHRs [34]. As O2- scavenges NO, bioactive NO is reduced [35]. Enzymes such as GPx and catalase protect from an oxidative attack, reducing H2O2 to H2O. The present study showed that the administration of the ethyl acetate extract of radish leaves significantly increased GPx and catalase activities in RBCs.
It is interesting how ACE activity of the RAAS is inhibited and suppressed by hypertension treatment [35]. With regard to RAAS, ACE activity should be lowered after administration of ACE inhibitors. However, no significant effect of the radish leaf extract on the ACE activity in the lung, kidney, or serum was observed in the groups treated with the extract. The effect of the radish leaf extract on ACE activity was found only in the pulmonary tissue, and this effect was dose dependent, but not significant. Zhang and Tan reported that ACE activity in the plasma, but not the lungs, was significantly inhibited by aqueous extract of Andrographis paniculata [36].
NO concentration in the blood is associated with antiatherogenic effects and may help explain the vascular pathology of hypertension [37]. NO is produced from L-arginine by NO synthase (NOS) [38,39]. NO dilates blood vessel and inhibits platelet aggregation and proliferation of vascular smooth muscle cells [39]. It has been reported that supplementation of L-arginine, the precursor of NO, lessens the extent of atherosclerosis in diet-induced hypercholesterolemic rabbits. It is recognized that NO levels are generally low in hypertensive patients, but there was no difference in NO levels between normotensive and hypertensive rats in the present study. NO also suppresses the synthesis of ACE in the endothelial cells [40]. In our study, we found that the serum NO levels were higher in the groups fed with the radish leaf extract. The radish leaf extract increased the serum concentration of NO and showed a significant effect on spontaneous hypertension.
The RAAS regulates sodium and water contents and arterial blood pressure via angiotensin and aldosterone regulation [41]. The excretion of Na+ and K+ is associated with hypertension. Healthy people take in moderate amounts of electrolytes, and only adequate amounts of electrolytes are excreted to excrete adequately for homeostasis. In pathological conditions, the RAAS does not function properly, and the restriction of Na+ intake is needed [42]. Khaw et al. [43] have shown that a negative interaction between K+ and hypertension. In our study, the fecal concentrations of Na+ and K+ significantly increased with increase in the quantity of the radish extract. Urinary excretion of Na+ is about 80-90% of the intake amount and that of K+ is about 68% of the intake amount [29,31]. In the present study, the urinary concentration of Na+ in the normotensive controls was significantly higher than that in the hypertensive controls. There was no difference in the urinary concentration of Na+ among groups fed with the radish leaf extracts. Therefore, our results indicate that the radish leaf extract might inhibit Na+ and K+ absorption.
In summary, we observed that consumption of an ethyl acetate extract of radish leaves may have antihypertensive effects in SHRs since it increases the serum concentration of NO, and enhance antioxidant activities. Our results suggest a possible beneficial effect of radish leaf on antihypertensive function and provide a basis for its use as a medicinal food.





 XML Download
XML Download