

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The Korean alcoholic beverage, Takju, is made using a brewing process with the yeast Saccharomyces cerevisiae. The sugar necessary to produce alcohol is converted from rice starch to make Takju. Heavy alcohol intake places a heavy burden on the liver, whereas Takju does not because it contains about 6% alcohol and 1.5-1.9% protein, sugar, riboflavin, as well as other useful nutrients [1]. Hong et al. [2] reported that Takju possesses effective scavenging activity against free radicals, which is similar to the radical scavenging activity of wine. Takju also has functional effects on inflammation [3], cancer [4], hyperglycemia in diabetic rats [5,6], and hypertension [7].
Physical activity has well-known benefits for several chronic disorders including coronary artery disease, stroke, diabetes mellitus, and osteoporosis [8-10]. Furthermore, exercise is known to cause oxidative stress in the nervous system and increase lipid peroxidation, which may lead to alterations in neuronal membrane permeability. In clinical settings, the beneficial effects of physical fitness interventions on memory and other aspects of cognition have been documented in elderly individuals [11-15]. Regular physical exercise may promote neural health and function. Both animal and human studies have shown that exercise improves cognitive function [16,17]. Notably, exercise enhances learning and memory [16,18].
Ethanol is stressful to animals and exerts an oxidative stress response on the nervous system. After ingestion, ethanol is quickly absorbed from the gastrointestinal lumen into circulation and can readily cross the blood brain barrier entering areas of the brain. Due to its high lipid solubility, ethanol also changes the permeability and fluidity of synaptic membranes as well as the activities of membrane-bound enzymes [19].
Acetylcholinesterase (AChE) is a membrane-bound enzyme that degrades the neurotransmitter acetylcholine, producing choline and an acetate group, and has very high catalytic activity. AChE is mainly found at neuromuscular junctions and cholinergic synapses in the central nervous system, where its activity serves to terminate synaptic transmission. Acetylcholine is a neurotransmitter common to many synapses throughout mammalian nervous systems. AChE is bound to cellular membranes of excitable tissues at cholinergic synaptic junctions, which are usually associated with nerve impulse conduction. Progressive loss of cholinergic neurons in patients with Alzheimer's disease (AD) results in severe memory loss and impairment of cognitive function. AChE activity, a marker for degeneration of the central cholinergic system, is reduced in the cerebral cortex of patients with AD. A strategy for patients with AD is to treat with AChE inhibitors, which increase the availability of acetylcholine in the central synapses and contribute to modest improvements in memory, thinking, and reasoning skills [20]. Kwak et al. [21] reported that AChE inhibitory effects are influenced by concentrations of green tea extract and by water activity. Therefore, green tea, may serve as a potential dietary source of AChE inhibitors. Laurin et al. [22] suggested that physical activity is associated with lower risks of cognitive impairment, AD, and dementia of any type. Significant trends of increased protection with greater physical activity were observed. Several studies have found significant correlations between AChE levels in the cerebral cortex and spatial problem solving ability [23,24]. It appears that training alters the AChE activity of the cortex and informal enriched experiences lead to increased cortical AChE activity [25]. AChE activity is higher in the cerebral cortex of groups that have been trained and tested on more difficult problems than in those given easier problems. An experiment in which littermates were either trained on a difficult problem or were untrained reported that the trained rats developed significantly higher cortical AChE than their untrained littermates [26]. These data suggest that the intensity of physical and mental training may influence AChE expression and activity differently.
Because exercise and ethanol are effective for cholinergic system activity [27], we investigated the effects of exercise training and Takju and ethanol consumption on brain AChE activity and learning ability in rats. This study may prove useful to researchers who are addressing the functional compounds of Takju.
Materials and Methods
Animal care and selection
Thirty-six 6-week-old male Sprague-Dawley rats were obtained from Central Lab., Animal Inc., (Seoul, South Korea), and housed in individual wire cages in a controlled environment of 23 ± 1℃, 60 ± 10% relative humidity, and a 12-hour light-dark cycle (lights on at 7 a.m.). The cages were cleaned every second day. After 2 weeks of acclimation, the animals were divided by initial body weight into six blocks of six rats each, using a randomized complete block design. The rats were treated with exercise and alcohol factors for 4 weeks in separate groups as follows: non-exercised control (CC), exercised control (EC), non-exercised consuming ethanol (CA), exercised consuming ethanol (EA), non-exercised consuming Takju (CT), and exercised consuming Takju (ET).
Experimental procedures
The daily experimental schedule is shown in Fig. 1. All aspects of this study were conducted according to the standards of the Animal Ethics Committee of Kookmin University. Exercise training was performed at a speed of 10 m/min for 15 minutes per day. The rats in the CA and EA groups were administered 6% ethanol as a Takju control and those in the CT and ET groups were given the Korean rice wine, Takju. Deionized water, ethanol, or Takju were provided at 9 a.m. after 8 hours of deprivation to promote drinking. The ethanol and Takju were administered for 4-5 hours to achieve intakes of approximately 10 ml, and thereafter the animals were allowed free access to deionized water.
Experimental diets
When the animals were assigned to the experimental protocol, they were given free access to deionized water and a Purina pellet diet (Central Lab., Animal Inc.,) for the first 2 weeks. After 2 weeks of acclimation, the rats were switched to a powdered AIN-93M diet [28], which was provided ad libitum. The Takju (Seoul Jangsu, Seoul Takju Union Co, Seoul, South Korea) was purchased from a local liquor store and stored at 4℃ until use. It contained 6% alcohol, 10% isomaltooligosaccharides, and 0.01133% aspartame. Ethanol was used as a control and was diluted to 6% from absolute ethanol (Sigma Aldrich, St. Louis, MO, USA). The ingredients in the experimental diets were corn starch (Daesang Co., Seoul, South Korea), dextrose (Samchun Pure Chemical Co., Gyeonggi-do, South Korea), casein (Daejung Chemical and Metal Co., Gyeonggi-do, South Korea), soybean oil (Cheiljedang Co., Seoul, South Korea), sucrose (Samyang Co., Ulsan, South Korea), cellulose, tetrabutyric acid, choline bitrate (Sigma Aldrich), a vitamin mixture, and a mineral mixture (G-Bio Co., Gyeonggi-do, South Korea).
Exercise training protocol
All animals in the exercised groups were trained on a motor driven work wheels (120 cm in circumference × 15 cm × 5 tracks) (Dae-jong Instrument Industry Co., Seoul, South Korea) 6 days per week. The rats in the exercised groups were forced to run on a treadmill for 1,520 minutes, whereas those in the non-exercised groups were left in their cages. The experimental procedures were performed to expose the animals to a moderate intensity and to maintain a healthy status [29,30]. An electrical shock grid at the back of the treadmill provided a mild but aversive foot shock (50 V) to encourage the rats to run. Very few shocks were administered during the training sessions. Exercise was started at 15:00 each day in a dark chamber. During the 2 weeks of acclimation, training was performed by the training groups at a speed of 10 m/min for 15 minutes. After 2 weeks of acclimation, exercise training was performed for 20 minutes at 10 m/min for 3 weeks and increased to 15 m/min on the final fourth week of the experiment. This training protocol resulted in a training intensity of 20-30% Vo2 max [30]. This procedure was used to expose the animals to moderate speed running.
Maze test
We conducted a Y-shaped water maze test (Fig. 2), which is widely used to evaluate learning and memory in rodents [31]. The maze setup and test procedures were the same as used in our earlier study [32]. The maze was made of three arms measuring 20 × 70 cm and 45 cm in depth and positioned at equal angles; it was filled with clear tap water at a temperature of 25℃ and to a depth of 30 cm. Movable escape platforms were placed in the two arms of the maze and submerged 1 cm below the water surface. The experiments were performed between 17:00 and 18:00 h.
During the adaptation period, the animals were given a swim trial from the starting position to the platforms at the end of the Y-maze stem for 5 days. The side of the platforms that the animals stepped on was recorded, and the side with greater use was regarded as the preferred direction of the animal. The animals established a position preference for either the right or left arm and would consistently go to that arm. Next, two repeated phases of reversal training were performed during each of 5 days by removing the platform on the preferred side and forcing the animals to swim to the opposite arm. During the second phase, the platform was moved to the other side, so that the rats swam to reverse sides from the first phase. Errors were scored when a rat entered the arm that did not contain the platform. The number of errors and the time to reach the platform in seconds as a parameter of escape latency were measured daily to evaluate performance.
Animal sacrifice and tissue collection
At the end of the experimental period, unanesthetized animals were decapitated after 12 hours of fasting. The brain was rapidly removed from the cranium and cut in half on an ice-cold plate. The right brain was immersed immediately in dry ice. The tissues were stored at -70℃ until analysis and were analyzed within 1 month of sacrifice.
AChE activity
AChE activity was assayed by spectrophotometrically measuring the rate of thiocholine production as acetylcholine was hydrolyzed [33]. Enzyme activity was measured by following the increase in yellow color produced from thiocholine when it reacted with dithiobisnitrobenzoate ions. Brain tissue was homogenized in 18 ml of 0.1 M phosphate buffer (pH 8.0) in a homogenizer (OMNI International Inc. Kennesaw GA, USA) and centrifuged at 7,600 × g at 4℃ for 10 minutes [34]. A 0.2 ml aliquot of the supernatant was added to a cuvette containing 2.8 ml of 0.1 M phosphate buffer. Of 100 ml of the DTNB reagent (0.01 M 5:5-dithiobis-2-nitrobenzoic acid in 0.1 M phosphate buffer), 100 µl was added to the cuvette. The absorbance was measured at 412 nm. Then, 20 µl of the substrate (0.075 M acetylthiocholine iodide substrate), was added. After 2 min, absorbance was recorded, and change in absorbance per minute was calculated. The rates were calculated as follows
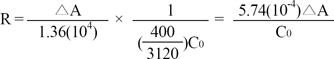
Where
R = rate, in moles of substrate hydrolyzed per min per g of tissue;
ΔA = change in absorbance per min;
C0 = original concentration of tissue (g/ml)
Statistical analysis
All data were analyzed using SPSS Ver. 18.0 (SPSS, Inc., Chicago, IL, USA). A one-way analysis of variance (ANOVA) with Duncan's multiple range test was conducted to analyze body weight, feed intake, AChE activity, and the water maze data, and the data are expressed as means or means ± standard error of the mean. A 2 × 3 ANOVA was performed on the two factors of exercise training and ethanol consumption. The Y-shaped water maze data were further analyzed via two-way repeated measures ANOVA and the nonparametric independent Mann-Whitney U and Wilcoxon signed rank tests for each testing phase. All significance levels were set at P < 0.05.
Results
Feed intake and weight gain
Final body weight, weight gain, and total caloric intake are presented in Table 1. Caloric intake was calculated as the sum of calories from the diet and alcohol. The number of calories from the Takju and ethanol was 0.42 kcal/ml. Total caloric intake was not significantly different among the groups, as the consumption of Takju and ethanol did not affect caloric intake. Final body weights and weight gain were also not significantly different among the groups. Exercise had no effect on weight gain, even though weight gains were slightly less in the exercised groups than in the non-exercised groups.
AChE activity
The brain AChE activities are presented in Table 2. The highest AChE activity was observed in the EC group, and the lowest activity was observed in the CC group. However, no significant differences in AChE activity were observed among the groups. The exercised groups tended to have increased AChE activity when compared to the non-exercised groups, which agreed with earlier data showing that exercise significantly increases AChE activity [35].
Learning ability
The results of the position reversal test in the maze are given in Table 3. The number of errors was significantly lower in the EA group than that in the CA and ET groups, indicating that exercise decreased the number of errors in the animals that drank ethanol, and that ethanol caused fewer errors than that of Takju. However, exercise training and alcohol intake, were not significantly different according to the two-way ANOVA. In the case of swim time to escape from the water by stepping on a platform, latency showed a proportional trend with the number of errors because the animals spent more time in the water by entering the wrong side. Therefore, the latency of the EA group was significantly shorter than that of the CC, EC, CT, and ET groups. Latency and the number of errors during the second phase showed a similar trend to those in the first phase, but the difference was not statistically significant.
Fig. 3 shows the time difference from latency on the first day of position reversal training to latency on the fifth day. The time difference of the ET group was significantly larger than that of the CC, CA, EA, and CT groups.
The results of exercise training and alcohol intake are shown in Fig. 4 and 5, respectively. In Fig. 4, latency time and the number of errors decreased rapidly from days 1 to 5 in all groups. On the first day, latency and the number of errors were significantly higher in the exercised groups than in the non-exercised groups (Fig. 4).
Discussion
The purpose of this study was to investigate whether moderate exercise training and Takju ingestion would influence AChE activity in rat brains. We was also intended to determine whether exercise and Takju are effective for memory and learning ability.
Approximately 10 ml of 6% alcohol intake and moderate exercise training at a speed of 10 m/min for 15 minutes did not affect weight gain as was intended. The animals obtained 4.2 kcal/d by consuming 10 ml alcohol and were provided 1.1 g of experimental food with the alcohol. Hencno significant differences were observed in energy intake among the groups. Our results agree with studies in which rats trained using moderate intensity exercise showed no differences in weight gain [30,36]. Monteiro et al. [37] reported that the consumption of red wine containing 13% ethanol caused no difference in energy intake compared to a control.
Ethanol (1.0 or 1.5 g/kg) administered intraperitoneally evokes behavioral changes characteristic of intoxication and depressed acetylcholine release from the cerebral cortex [27]. These doses of 1.0 and 1.5 g/kg were similar to our study, although the intake method was different. Therefore, our 10 ml of 6% alcohol consumed was considered effective enough to influence acetylcholine.
Husain and Somani [38] studied the effects of exercise and ethanol on AChE activity in the brain of rats, and reported selective striatal AChE inhibition by acute exercise and 1.6 g/kg ethanol intake. The dose of 1.6 g/kg is almost the same as in our experiment. Exercise training for 6.5 weeks decreases AChE activity significantly (64% of control) in the hypothalamus but increases AChE activity in the cerebral cortex (149% of control). Chronic ethanol ingestion of 2.0 gm/kg decreases AChE activity in the hypothalamus (63% of control). Distribution of AChE varies in different brain regions [39-41]. Ethanol likely influences enzyme activity differently.
Significant correlations are observed between AChE enzyme levels in the cerebral cortex and the ability to solve spatial problems [42], and informal enriched experiences lead to increased cortical AChE activity [43]. In contrast, exercise can decrease AChE activity [35]. In the current study, the results show that the exercised EC, EA, and ET groups tended to have increased brain AChE activity, but that the differences were not significant. Similarly, alcohol ingestion revealed a tendency for elevated brain AChE. These results disagree with Wen et al. [35] in which AChE activity decreased transiently by exercise-induced fatigue and then gradually increased over 24 hr. The various controversial results on exercise and AChE activity levels are due to various exercise intensities and alcohol levels.
We chose a left-right discrimination paradigm in the Y-maze, rather than the more often applied Morris water maze. The water maze paradigm involves both handling and swim stress, and causes a considerable elevation in plasma corticosterone levels [44]. We attempted to design our experiment so that stress levels were minimized. Although Y-maze learning can be stressful to rats due to the novelty [45], we reduced stress associated with this task by habituating the rats to the maze and by maintaining the water temperature at 25℃. When we conducted the Y-maze test, the animals escaped from the water very quickly. Thus, the number of errors and latency time emerged as good indicators to measure learning ability. Our results show that the number of errors was significantly lower in the EA group than that in the CA group, indicating that exercise training decreased the number of errors on the water maze. Similarly, latency during the first phase in the EA group was significantly shorter compared to that in the CC, EC, CT, and ET groups. Latency and the number of errors during the second phase exhibited a similar trend as those in the first phase, but the differences were not statistically significant. The latency difference from days 1 to 5 was significantly larger in the ET group than that in the CT group. In addition, the latency from days 1 to 5 was larger in the alcohol groups and the Takju groups (EA, CT, and ET groups) than that in the control CC and CE groups. These results suggest that exercise and Takju helped to increase learning ability. We cannot exclude the possibility that higher doses and/or a longer duration of Takju ingestion would result in significant alterations in learning ability. In summary, the results indicate that moderate exercise increased memory and learning without neurochemical changes in AChE activity and that the combination of exercise and Takju ingestion may enhance learning ability during a short and initial period.







 XML Download
XML Download