

INTRODUCTION
Hypoxia inducible factor (HIF) regulates the expression of proteins that increase oxygen delivery, which enables cells to survive in oxygen-deficient conditions. HIF-1 inhibitors could be useful as therapeutic agents for various diseases associated with the over-activation of HIF-1. Epithelial-mesenchymal transition (EMT) is involved in many crucial cell functions, including tissue reorganization, tumorigenesis, cancer recurrence, and metastasis [3]. Thus, many studies have examined the EMT process to understand the disease mechanism from a cancer-related perspective. Interestingly, EMT has been studied from many different perspectives revealing that it is not restricted to cancer, but is also implicated in upper airway sinonasal diseases such as chronic rhinosinusitis (CRS) and nasal polyp, and moreover, in lower airway diseases such as bronchial mucosa remodeling [4]. Recent evidence shows that hypoxia is associated with nasal polypogenesis and CRS, and that hypoxia drives the EMT via the HIF-1 and Smad3 signaling pathways [5]. In this study, hypoxia contributes to nasal polypogenesis by inducing the EMT not only in primary nasal polyp epithelial cells but also in the A549 airway epithelial cell line. Because a number of airway diseases involve lower (bronchial) and upper (sinonasal) airway mucosa, the airway epithelial cell line is used in sinonasal disease research.
CRS, which is one of the most prevalent diseases in the rhinologic field, has a relatively poor prognosis owing to its propensity for conjoined nasal polyp relapses and poorly known mechanisms. Sustained inflammation in sinonasal cavities caused by several etiologies leads to hypoxia. Hypoxia is one of the mechanisms that relates to the genesis of nasal polyps and CRS [12]. Although the role of hypoxia in CRS is not fully understood, the underlying mechanisms contributing to hypoxia-mediated CRS are partially dependent on the EMT of airway epithelial cells.
Wheatgrass refers to the young grass of the common monocot wheat plant Triticum aestivum. It contains vitamins, minerals, enzymes, amino acids, polysaccharides, and large amounts (70%) of chlorophyll. Several papers have indicated that wheatgrass has anti-tumor, anti-apoptotic, anti-proliferative, anti-oxidant, and therapeutic effects on distal ulcerative colitis, diabetes, and heart disease [67891011]. However, there are no reports on whether wheatgrass has an effect on human airway epithelial cells.
Considering that inhibition of the hypoxia-induced EMT can be a promising therapeutic target for the treatment of CRS, searching for new agents or compounds having such a function is an important alternative strategy. In this study, we evaluated the EMT-inhibiting effect of wheatgrass extract under hypoxic conditions in airway epithelial cells.
Go to : 
MATERIALS AND METHODS
Materials
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was obtained from Sigma-Aldrich (St. Louis, Missouri, USA). The other chemicals used were of the purest grade available from Sigma.
Preparation of wheat sprout sample
The wheatgrass sprout was harvested in 2015 from a commercial planting location in Gwangju, Korea.
Extract preparation
After grinding dried wheat sprouts in a mixer, they were extracted at 25℃ with 80% ethanol for three days in shaking incubators (200 rpm). They were concentrated in a rotary evaporator. The ethanol extracts were fractionated with ethyl acetate or water. The ethyl acetate layer was further separated with n-butanol and water. Then, the butanol fraction was separated with hexane and water. The n-ethanol fraction was utilized in all experiment.
Cell cultures and transfection
A549 cells were maintained in RPMI 1640 (Gibco BRL, Grand Island, NY, USA) media supplemented with 1% penicillin/streptomycin (Gibco/Invitrogen, Carlsbad, CA, USA) and 10% fetal calf serum. The cells were grown to 60% confluence in 100-mm culture plates, treated with different concentrations of wheatgrass extract (50-150 µg/mL) for 30 min, and then incubated in hypoxic conditions (5% CO2/1% O2) or in normoxic conditions (5% CO2/20% O2) for an additional 24 h.
Cells were transfected with control small interfering (si)-RNA or HIF-1α si-RNA using Lipofectamine RNAiMAX (Invitrogen) in 60-mm plates according to the manufacturer's protocol. The si-RNA was #1068432V for HIF-1α (Bioneer, Daejeon, Korea).
Cell proliferation assay
Cell growth inhibition was analyzed using the MTT assay. Briefly, cells cultured overnight on 96-wellplates were washed twice with PBS, administered media containing different concentrations of wheatgrass, and incubated for 24 h under hypoxic conditions. Ten microliters of MTT stock solution (5 mg/mL) was added to each well, followed by an additional 4 h incubation. Blue formazans were eluted from cells by the addition of 100 µL DMSO with gentle shaking for 10 min at room temperature. Absorbance was measured at 570 nm using an enzyme-linked immunosorbent assay reader (SpectraMAX; Molecular Devices, Sunnyvale, CA, USA).
Nuclear protein extract
The nuclear protein was extracted using NE-PER® Nuclear and Cytoplasmic Extraction Reagents (Pierce, Rockford, IL, USA) according to the manufacturer's instructions. After 24 h incubation in hypoxic conditions, the medium was removed and cells were washed with phosphate-buffered saline, pelleted by centrifugation at 13,000×g for 2 min, and resuspended in CER I buffer. Cells were allowed to swell on ice for 10 min and then ice-cold CER II was added to the cell pellet for 1 min on ice. Cells were vortexed to disrupt cell membranes and centrifuged at 13,000×g at 4℃ for 5 min. The supernatant was stored at −70℃ until used as the cytoplasmic extract. The pelleted nuclei were resuspended in the nuclear extraction buffer, placed on ice, and vortexed for 15 sec every 10 min for 40 min. The nuclear extract was collected by centrifugation at 13,000×g for 10 min at 4℃. Protein concentration of the nuclear extract was estimated using Bradford's reagent (BioRad, Hercules, CA, USA).
Immunoblot analysis
After 24 h incubation in hypoxic conditions, the medium was removed and cells were washed with phosphate-buffered saline and lysed with lysis buffer (50 mM Tris pH 7.7, 150 mM NaCl, 1% NP-40, 5 mM EGTA, 50 mM glycerophosphate, 20 mM NaF, 1 mM Na3VO4, 2 mM phenylmethylsulfonyl fluoride, 10 mg/mL leupeptin, and 10 mg/mL aprotinin). After centrifugation at 13,000×g for 10 min at 4℃, the supernatant containing the total cell lysate was collected. Equal amounts of protein were separated by electrophoresis on 10–12% Tris-HCl gels and were transferred to a PVDF membrane. After blocking with TBS-T (20 mM Tris, 500 nM NaCl, and 0.1% Tween-20) containing 5% (w/v) skim milk, the membrane was incubated with a specific primary antibody for E-cadherin (Santa Cruz Biotechnology, Dallas, TX, USA), N-cadherin, Snail, Phospho-Smad3, Smad3 , Lamin (Cell Signaling Technology, Danvers, MA, USA), HIF-1α (Novus, Littleton, CO, USA), or GAPDH (Santa Cruz Biotechnology) followed by peroxidase-conjugated anti-mouse immunoglobulin G (IgG) or anti-rabbit IgG (Jackson Immuno Research, West Grove, PA, USA). The membranes were developed using the enhanced chemiluminescent analysis system. Results were obtained from three independent experiments.
Immunofluorescence
Cells were incubated in hypoxic conditions for 24 h or transfected with si-HIF-1α mRNA and control si-mRNA, treated with different concentrations of wheatgrass (50–150 µg/mL), and then incubated in hypoxic conditions for 24 h on a cover slide in 12-well plates. Cells were rinsed three times with PBS, fixed with 4% paraformaldehyde for 10 min at room temperature, and rinsed again. Cells were then blocked with 1% bovine serum albumin, followed by the addition of the primary antibodies: anti-E-cadherin, anti-N-cadherin, anti-Snail, or anti-HIF-1α. After extensive washing with PBS, fluorescein isothiocyanate-conjugated IgG was added. DAPI was used to counterstain the nuclei. Following incubation, the slides were rinsed, mounted, and viewed at 488 nm on a confocal microscope (FV1000, Olympus, Tokyo, Japan).
Statistical analysis
The results were analyzed statistically using SPSS 19.0 software (SPSS Inc., Chicago, IL, USA). Student's t-tests were used for pairs of data. A P value < 0.05 was considered to be statistically significant.
Go to :
RESULTS
Wheatgrass inhibits the hypoxia-mediated EMT in airway epithelial cells
Wheatgrass extract per se did not affect the viability of airway epithelial cells (data not shown). In an effort to identify new EMT inhibitors, we screened natural extracts from wheatgrass for their ability to alter the fibroblastic morphological changes induced by hypoxia in airway epithelial cells. Incubation in hypoxic conditions (O2 1%) induced the expected morphological changes in epithelial cells at 24 h (data not shown). Consistent with the observed morphological changes in hypoxia-exposed airway epithelial cells, reduced expression of the epithelial marker E-cadherin was observed, whereas the expression of the mesenchymal marker N-cadherin increased. When the cells were pre-treated with wheatgrass for 30 min and then incubated for 24 h in hypoxic conditions, E-cadherin expression was restored (Fig. 1A, B, and C, P < 0.01), whereas that of N-cadherin gradually decreased as wheatgrass concentration increased (Fig. 1D and E, P < 0.01). These data suggest that wheatgrass inhibits the EMT in airway epithelial cells.
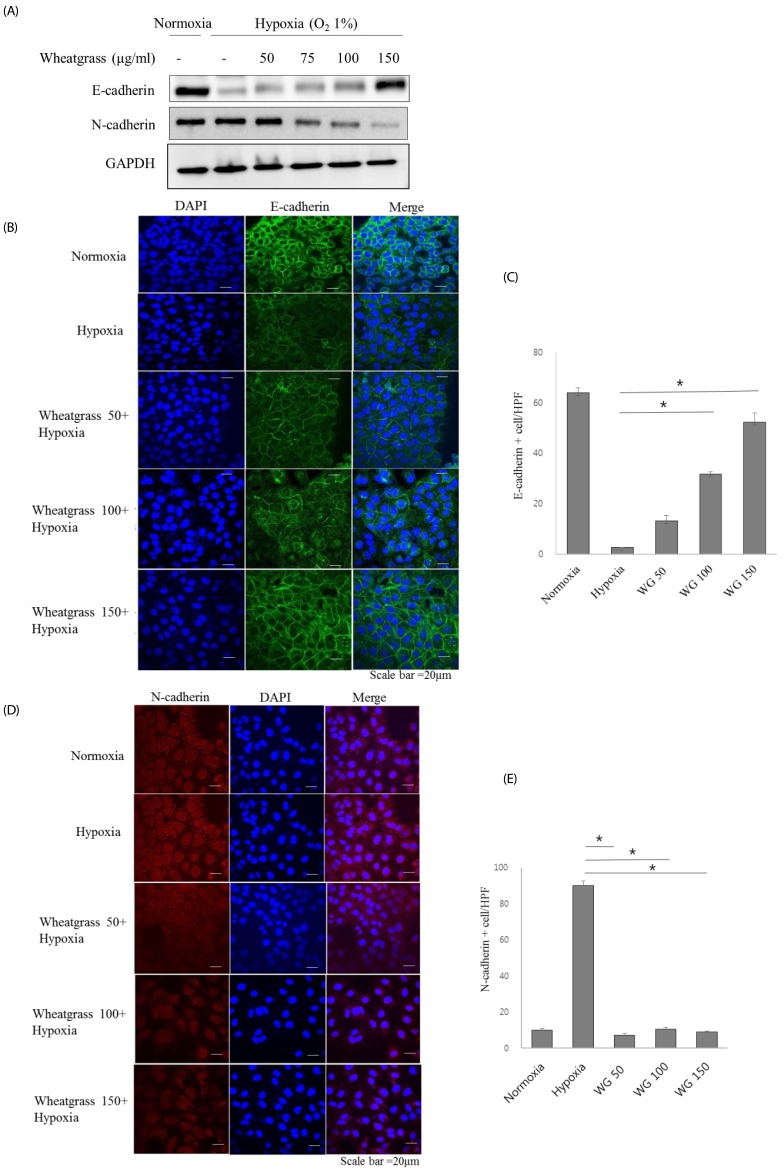 | Fig. 1Wheatgrass inhibits the hypoxia-mediated EMT process in airway epithelial cells.(A) A549 cells were treated with wheatgrass at the indicated concentrations for 24 h and total cell lysates were analyzed by immunoblotting with anti-E-cadherin or N-cadherin antibodies. (B and C) E-cadherin levels were visualized by immunofluorescence analysis. Green fluorescence indicates E-cadherin expression, blue denotes DAPI staining of the nucleus, and the right panel is a merged image of the two panels. Scale bar represents 20 µm. (D and E) N-cadherin expression levels were visualized by immunofluorescence analysis. Red fluorescence indicates E-cadherin expression (*P < 0.01). WG, wheatgrass; EMT, epithelial mesenchymal transition; HPF, high power field.
|
Wheatgrass negatively regulates HIF-1α expression in airway epithelial cells
Various signaling pathways contribute to inducing an EMT. We investigated the relationship between HIF-1α and EMT, and the effect of wheatgrass on HIF-1α expression. We found that wheatgrass down-regulated HIF-1α expression (Fig. 2A). To further evaluate the role of HIF-1α, we transiently knocked down HIF-1α expression using si-HIF-1α mRNA. Loss of the E-cadherin marker due to hypoxia was recovered following si-HIF-1α mRNA transfection and in cells treated with wheatgrass (Fig. 2B and C, P < 0.01). Hypoxia-induced HIF-1α expression was decreased by wheatgrass treatment or si-HIF-1α mRNA transfection to a similar extent (Fig. 2D and E, P < 0.01). Together, these results suggest that wheatgrass suppresses the EMT by inhibiting the activation of HIF-1α.
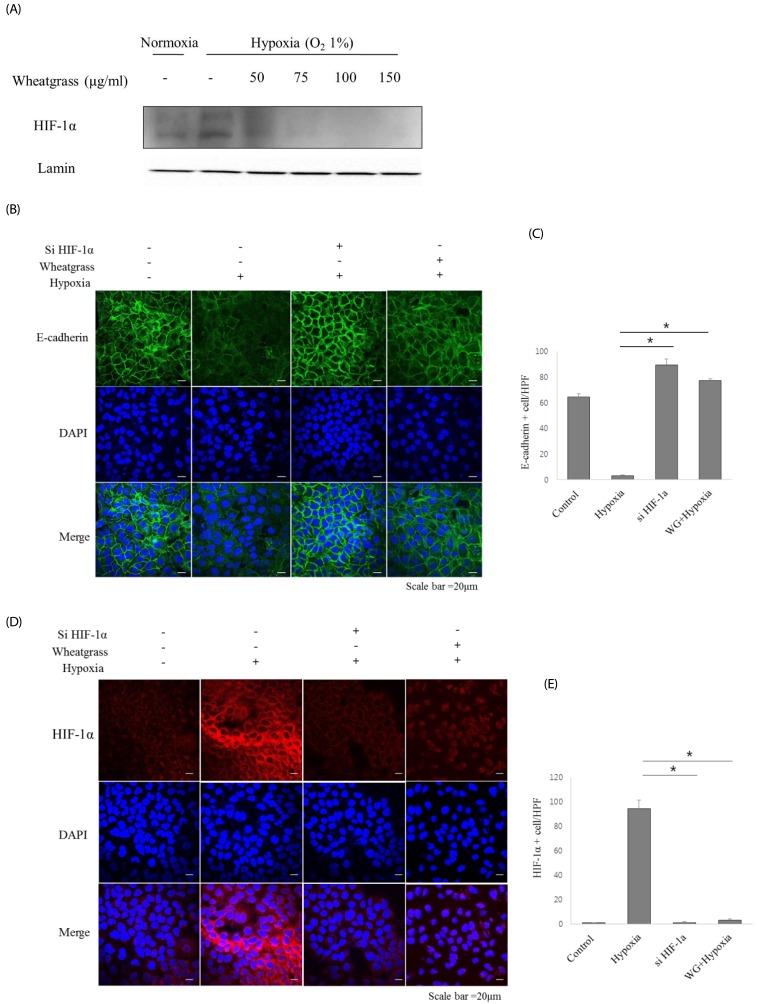 | Fig. 2Wheatgrass inhibits the hypoxia-mediated EMT process by inhibiting the activation of HIF-1α in airway epithelial cells.(A) A549 cells were treated with wheatgrass at the indicated concentrations for 24 h, and nuclear protein fractions were analyzed by immunoblotting with an anti-HIF-1α antibody. (B and C) Cells were transfected with either si-HIF-1α, si-negative control (NC), wheatgrass (150 µg/mL), or hypoxia only as indicated. E-cadherin expression levels (green) were analyzed by immunofluorescence analysis. (D and E) HIF-1α expression levels (red) were visualized by immunofluorescence analysis (*P < 0.01). WG, wheatgrass; EMT, epithelial mesenchymal transition; HIF, hypoxia inducible factor; HPF, high power field.
|
Wheatgrass suppresses EMT by reducing Smad signaling
The Smad signaling pathway facilitates the induction of hypoxia-mediated EMT. We found that wheatgrass affected the phosphorylation of Smad3. The phosphorylated Smad3 signal intensity was maintained for 6 h after hypoxia exposure. However, it decreased in an extract dose-dependent manner (Fig. 3A and B). Immunofluorescence analyses showed that Smad3 localized to the cell nucleus following hypoxia exposure, whereas it localized in the cytosol in the normoxic state. Hypoxia-induced nuclear localization of Smad3 expression was reduced by the wheatgrass treatment (Fig. 3C and D, P < 0.01). These data suggest that wheatgrass suppresses the EMT by inhibiting the activation of Smad.
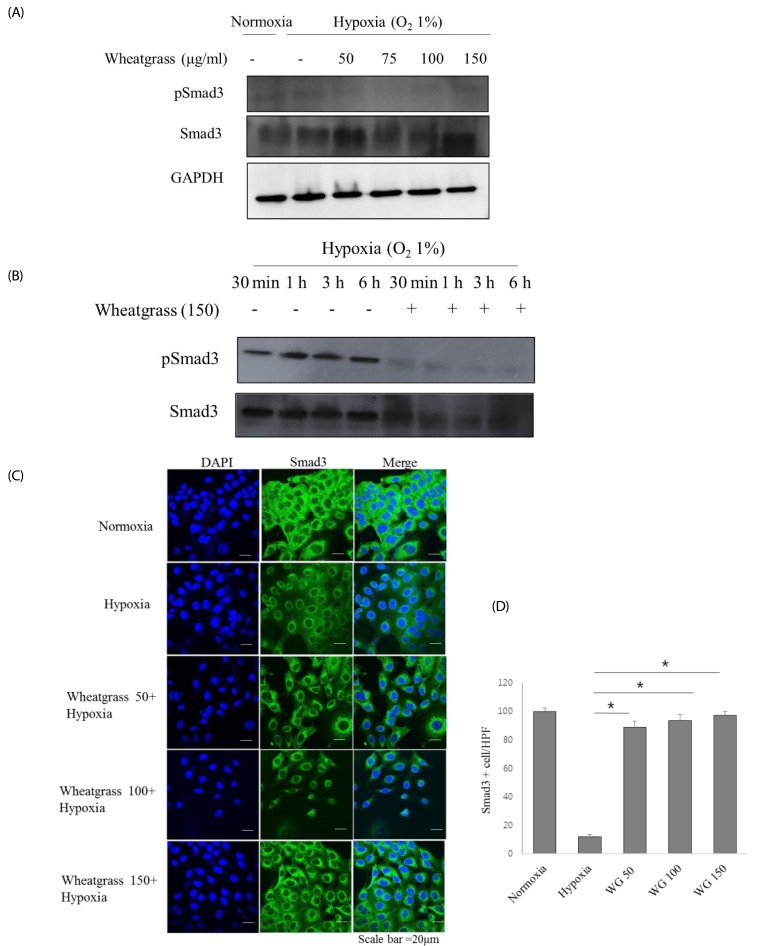 | Fig. 3Wheatgrass inhibits the hypoxia-mediated EMT process by reducing Smad activation.(A and B) Cells were treated and cultured with wheatgrass at the indicated concentrations and times. (C and D) Smad3 expression levels (green) were analyzed by immunofluorescence (*P < 0.01). WG, wheatgrass; EMT, epithelial mesenchymal transition; HPF, high power field.
|
Wheatgrass negatively regulates Snail expression in airway epithelial cells
Snail, Slug, and ZEB 1/2 are well known EMT-related transcriptional factors that suppress epithelial marker expression and enhance mesenchymal marker expression. Pre-treatment with increasing concentrations of wheatgrass down-regulated Snail expression in parallel with increased E-cadherin expression (Fig. 4A). Immunofluorescence analyses demonstrated that the hypoxia-mediated nuclear localization of Snail was reduced by wheatgrass treatment (Fig. 4B and D, P < 0.01). Collectively, these results suggest that wheatgrass negatively regulates the EMT by inhibiting the expression of Snail that is responsible for repressing E-cadherin expression, and that this process is governed by hypoxia-mediated Smad activation.
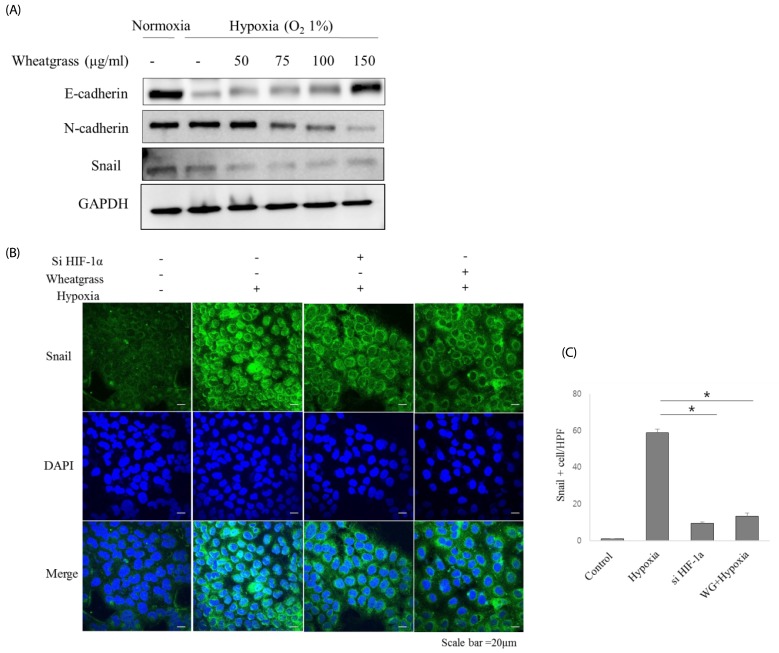 | Fig. 4Wheatgrass negatively regulates Snail expression in airway epithelial cells.(A) A549 cells were treated with wheatgrass at the indicated concentrations for 24 h, and total cell lysates were analyzed by immunoblotting with anti-E-cadherin, N-cadherin, and Snail antibodies. (B and C) Snail expression levels (green) were analyzed by immunofluorescence (*P < 0.01). WG; wheatgrass; HPF, high power field.
|
Go to :
DISCUSSION
We sought to identify whether wheatgrass extract has an EMT inhibiting capability. A previous study demonstrated that hypoxia contributes to nasal polypogenesis in chronic rhinosinusitis by inducing EMT not only in primary nasal polyp epithelial cells but also in A549 airway epithelial cell lines [5]. Using human A549 airway epithelial cells, we directly evaluated the effect of wheatgrass extract on several hypoxia-mediated EMT parameters, including expression levels of epithelial and mesenchymal markers. We found that wheatgrass inhibits the hypoxia-mediated EMT in airway epithelial cells and rescues the hypoxia-mediated downregulation of E-cadherin.
It has increasingly been recognized that hypoxia can result in the failure of transepithelial oxygenation, non-vascularized exudates, or the tendency of inflammatory hyperplasia to exceed neovascularization [12]. Moreover, in CRS, oxygen levels have been found to be lower in inflamed sinus cavities, supported by the up-regulated HIF-1α and osteopontin levels in the sinonasal tissue of patients with CRS [21314]. HIF is a transcription factor that activates genes required for adaptation to hypoxia. HIF-1α and 2α are expressed in nasal polyps and nasal epithelium and they induce EMT by altering the expression of EMT markers [125]. Thus, inhibition of HIF activity or the EMT might be a new target for innovative mechanism-based drug discovery for sinonasal disease.
Wheatgrass is a medicinal plant used in traditional medicine to cure various diseases. Because it contains chlorophyll, minerals, phenolic compounds, antioxidants, and flavonoids such as apigenin, quercetin, and luteonin, wheatgrass is primarily studied as a potent anti-carcinogenic agent. In one study, methanol extracts of wheatgrass inhibited human laryngeal cancer cell proliferation via cell cycle G1 arrest and p53 induction [15]. Another study demonstrated that chlorophyllin has the capability to reduce the risk of breast cancer by inducing cell cycle arrest and apoptosis in human breast cancer MCF-7 cells through ERK deactivation and cyclin D1 depletion [16]. In a study with a chronic myeloid leukemia cell line, aqueous and ethanol wheatgrass extracts inhibited the growth of cells in a time-dependent manner, induced apoptosis, increased malondialdehyde (oxidant) levels, and increased catalase and superoxide dismutase (antioxidant) activities [17]. Wheatgrass has beneficial effects on disease states such as beta-thalassemia, ulcerative colitis, skin wound healing, and diabetes mellitus [10181920]. However, few scientific or clinical studies exist for the use of wheatgrass in sinonasal disease.
In our study, immunofluorescence staining revealed that HIF-1α expression is closely related to the EMT process and both HIF-1α levels and the EMT are downregulated by wheatgrass treatment. In addition, the transfection of si-HIF-1α mRNA reduced the expression level of Snail and rescued E-cadherin expression in airway epithelial cells. Finally, wheatgrass inhibited the EMT by reducing HIF-1α activation. Wheatgrass appears to inhibit the hypoxia-mediated EMT by reducing the expression level of the EMT-related transcription factor, Snail, via the HIF-1α-Smad3 pathway. This suggests that wheatgrass may help to treat sinonasal disease, and could therefore have potential therapeutic relevance.
In conclusion, to the best of our knowledge, this is the first study to demonstrate that wheatgrass effectively inhibits the EMT processes in a concentration-dependent manner in airway epithelial cells. Although further studies are needed to examine the effects of wheatgrass, wheatgrass might have the potential to be developed into an anti-HIF-1-related disease agent.
Go to :