

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Obesity is a major risk factor for non-alcoholic fatty liver diseases (NAFLD). Macrophages/Kupffer cells are resident macrophages in the liver and are essential in both homeostasis and disease [1]. Excessive hepatic lipid accumulation in obesity promotes activation of hepatic macrophages/Kupffer cells, a major step in initiating hepatic inflammation, and steatohepatitis exacerbates insulin resistance and facilitates progression of non-alcoholic steatohepatitis (NASH) [23]. It is likely that the hepatic inflammatory responses largely depend on the macrophage phenotypes, which are characterized by their activation/polarization state into classically activated (M1) proinflammatory macrophages or alternatively-activated (M2) anti-inflammatory macrophages [45]. M1 macrophages are characterized by production of nitrogen oxide and other inflammatory cytokines such as tumor necrosis factor alpha (TNFα), interleukin-6 (IL-6); M2 macrophages are marked by upregulation of Mrc1, CD163, and IL-10 [6]. Macrophages/Kupffer cells in the liver are activated towards M1 phenotype in obese condition [7], whereas M2-polarized macrophages/Kupffer cells in the liver appear to protect against insulin resistance [5], alcoholic liver disease, and NAFLD [8]. Thus, strategies restraining M1 polarization and/or driving the alternative M2 activation of macrophages/Kupffer cells may be beneficial for protecting against hepatic inflammation and the progression of NASH.
Heme oxygenase-1 (HO-1), which is transcribed by activated nuclear factor erythroid 2-related factor 2 (Nrf2), a basic leucine zipper transcription factor [9], is the rate-limiting enzyme in the conversion of heme to biliverdin, releasing free iron and carbon monoxide (CO). It is critical in preventing oxidative stress, inflammation and metabolic dysregulation [91011]. HO-1 inducers such as hemin or substances that produce carbon monoxide such as cobalt protoporphyrin, reduce inflammatory cytokine expression in ob/ob mice and Zucker rats and improve insulin sensitivity in diabetic animals [1213]. Moreover, induction of HO-1 is likely to protect against macrophage-mediated inflammatory responses by preferentially promoting the M2 phenotype [1014]. With respect to this, we and others have shown that HO-1 induction by hemin reduces obesity-induced inflammation by polarizing macrophages in obese adipose tissue [15] and in alcohol-induced hepatic damage [16].
Some dietary components such as curcumin and resveratrol appear to control hepatic inflammation through regulation of HO-1 expression via the Nrf2 pathway [17], and these components affect phenotype switching of macrophages/Kupffer cells in liver [8]. Quercetin is a flavonoid that is abundant in fruits and vegetables such as apples, grapes, and onions. It possesses antioxidant and anti-inflammatory properties, and seems to inhibit obesity-induced inflammation and metabolic disorders such as insulin resistance in genetically and/or diet-induced obese rodents [181920]. We previously showed that it reduced obesity-induced inflammatory responses in both adipose tissue and skeletal muscle by inhibiting activation of inflammatory signaling pathways [2122]. It also protects against lipid accumulation, inflammation, fibrosis, and insulin resistance associated with NAFLD [232425]. However, the mechanism by which quercetin protects against obesity-induced steatohepatitis, in terms of its association with hepatic macrophage polarization, is unclear.
In this study, we demonstrate that quercetin reduces levels of inflammatory cytokines in the fatty liver of HFD-fed obese mice. This effect is associated with its action on HO-1 induction, and leads to polarization of hepatic macrophages toward the M2 phenotype. Furthermore, we show that the Nrf2/HO-1 pathway is involved in these effects. Quercetin may therefore be a useful dietary factor for protecting against obesity-induced steatohepatic inflammation.
MATERIALS AND METHODS
Animal experiment
Six-week-old (C57BL/6, male) mice were purchased from Orient Ltd. (Orient, Busan, Korea). After a one-week adaptation period, mice were randomly divided into three dietary groups (n = 6 per group). Mice fed on (1) a regular diet (RD) (13% of calories as fat from soybean oil; Harlan Teklad, Madison, WI, USA); (2) a high-fat diet (HFD) (60% of calories as fat from lard and soybean oil; Research Diets, New Brunswick, NJ, USA); and (3) an HFD supplemented with 0.05% (w/w) quercetin (HFD+0.05% Que) for 9 weeks. Quercetin was purchased from Jena Bioscience (Jena, Germany) with a purity of > 98%. All animal care and procedures were conducted according to the protocols and guidelines approved by the University of Ulsan Animal Care and Use Committee (2011-HA-003). The animals were killed by CO2 asphyxiation, and their livers were dissected.
Co-cultured hepatocytes and macrophages
Mouse primary hepatocytes were isolated and cultured using Life Technologies liver media and reagents (Grand Island, NY, USA) as described. The livers of anesthetized 6-8 week-old C57BL/6 male mice were perfused with liver perfusion and digestion medium at a flow rate of 5 ml/min. Digested livers were dissected and dissociated in liver wash medium. After filtration through a 100 µm nylon mesh filter, hepatocytes were purified by repeated centrifugation (three times) at 50 × g for 2 min. Mouse primary hepatocytes on 12-well plates (2 × 105 cells) were incubated with 200 µM free fatty acid, a palmitate coupled to fatty acid-free BSA (molar ratio, 5:1) in DMEM medium (Life technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS, Life technologies), and 1% penicillin-streptomycin (Life technologies) for 12 h. Murine macrophage Raw 264.7 cells were obtained from the Korean Cell Line Bank (KCLB40071, Seoul, Korea) and cultured in RPMI-1640 medium (Life Technologies) supplemented with 10% FBS, 1% penicillin-streptomycin and 0.2% gentamicin (Life Technologies) at 37℃ and 5% CO2. Raw 264.7 macrophages (8 × 104 cells) were plated with the lipid-laden hepatocytes, and the co-cultures were incubated in serum-free DMEM for 12 h in the presence of 20 µM of quercetin and/or 0.1 µM ZnPP, an HO-1 inhibitor.
Isolation and culture of peritoneal macrophages
Nrf2-deficient mice (B6/SJL background) were injected i.p. with 2 ml of 3% thioglycollate four days prior to being killed. Peritoneal macrophages were collected by lavaging the peritoneal cavity with 7 ml of HBSS. The cells were collected by centrifugation, washed, and suspended in EMEM medium supplemented with 10% FBS, 1% penicillin-streptomycin and 0.2% gentamicin. The cells were purified by adherence to tissue culture plates for 2 h. Peritoneal macrophages at 2 × 106 cells/ml were treated with 100 ng/ml LPS, 20 µM of quercetin for 24 h in serum-free medium.
Measurement of cytokines
Liver tissues were homogenized in lysis buffer (100 mM Tris-HCl and 250 mM sucrose, pH 7.4) with protease inhibitors (Sigma, St. Louis, MO, USA) and centrifuged at 10,000 g for 10 min. The levels of cytokines in the homogenates were evaluated according to the manufacture's instruction with a TNFα, MCP-1 ELISA set (BD Biosciences, San Diego, CA, USA) and an IL-6, IL-10 ELISA kit (R&D Systems, Minneapolis, MN, USA). Amounts of cytokine were adjusted for protein content, which was determined by using a BCA protein assay kit (Pierce, Rockford, IL, USA).
RT-PCR
Total RNA was extracted from liver tissue and cultured cells by using a Trizol reagent (Life technologies, Carlsbad, CA, USA) and then reverse-transcribed into cDNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA). For target genes expression analysis, cDNA was amplified using a SYBR premix Ex Taq kit (TaKaRa Bio Inc., Foster, CA, USA) using a Thermal Cycler Dice (TaKaRa Bio Inc., Japan). Results were analyzed with real-time system TP800 software (Takara Bio Inc.) and all values for genes were normalized to values for a housekeeping gene. The primers used in the analysis are listed in Table 1.
Western blot analysis
Samples of equal amounts were separated on SDS-PAGE and transferred onto nitrocellulose membranes (Millipore, Tullagreen, Carrigtwohill, Ireland). After incubation with antibodies against HO-1 (Enzo Life Sciences, Farmingdale, NY, USA) and β-actin (Sigma), membrane proteins were detected by HRP-conjugated secondary antibody (Cell Signaling, Beverly, MA, USA), and enhanced chemiluminescent substrate kit (PerkinElmer, Waltham, MA, USA). Intensity of protein bands was quantified by densitometry using Image J program.
Statistical analyses
Results are presented as means ± SEMs. Statistical comparisons were performed using one-way analysis of variance (ANOVA) with Dunnett and/or Student's t test using GraphPad Prism 5 (San Deiego, CA, USA). Null hypotheses of no difference were rejected if P-values were less than 0.05.
RESULTS
Quercetin supplementation reduces inflammatory responses in HFD-fed obese mice
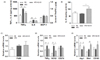
Our previous study demonstrated that quercetin did not alter food intake, and the HFD-fed mice supplemented with quercetin tended to gain less weight than the control HFD-fed mice [26]. In this study, we first examined whether quercetin supplementation reduced hepatic inflammation in HFD-fed obese mice. The obese mice supplemented with quercetin had lower levels of inflammatory cytokines (TNFα, and IL-6) in their livers than the HFD-fed controls (Fig. 1A) and higher levels of the anti-inflammatory cytokine, IL-10 (Fig. 1B). Quercetin had no effect on the expression of the macrophage marker F4/80 (Fig. 1C). To examine the subsets of macrophages/Kupffer cells, we further analyzed levels of the transcripts of macrophage phenotype specific markers. Dietary quercetin suppressed TNFα, and NOS2 transcripts, which are markers of M1 macrophages, (Fig. 1D) and increased those of the M2 macrophage markers Arg-1, and Mrc1 (Fig. 1E).
Effect of HO-1 inhibitor on quercetin actions in co-cultured lipid-laden hepatocytes/macrophages
To test for involvement of HO-1 in the effect of quercetin on the phenotypes of hepatic macrophages, we first confirmed that quercetin treatment upregulated HO-1 protein in macrophages (Fig. 2A), as previously reported [27]. Next, we co-cultured lipid-laden hepatocytes and macrophages to mimic steatohepatitis, and found that co-cultures resulted in increased inflammatory responses. Quercetin markedly reduced levels of MCP-1 release and expression of the M1 markers TNFα and NOS2 in the co-cultures (Fig. 2B-C), and these effects were antagonized by the HO-1 inhibitor ZnPP (Fig. 2B-C).
Effect of Nrf2 deficiency on the quercetin actions in hepatic inflammation
The Nrf2, a transcriptional regulator of HO-1, is important in the regulation of oxidative stress and the inflammatory state. We examined whether the HO-1 induction by quercetin occurred via the Nrf2 pathway. When we isolated peritoneal macrophages from Nrf2-deficient and wild-type mice, and stimulated them with LPS to induce macrophage M1 polarization and inflammation, we found that quercetin markedly increased HO-1 expression in the wild type and much less so in Nrf2-deficient mice (Fig. 3A). Along with this, we observed that quercetin suppressed the transcript levels of M1 macrophage marker CD274, and enhanced M2 macrophages marker CD163 in the LPS-stimulated WT peritoneal macrophages (Fig. 3B-C), but not in those from the Nrf2-deficient mice (Fig. 3B-C).
DISCUSSION
Obesity-induced steatohepatitis is characterized by increased activation of hepatic macrophages/Kupffer cells and infiltration of circulating monocytes/macrophages [2829]. In this situation, modulation of the inflammatory responses of hepatic macrophages might be beneficial for protection against inflammatory liver disease progression. In this study, we found that quercetin reduced levels of the inflammatory cytokines TNFα and IL-6, and enhanced the anti-inflammatory cytokine IL-10 in the livers of HFD-fed obese mice, indicating that quercetin protects against obesity-induced hepatic inflammation. Because hepatic inflammatory cytokine production is largely responsible for the activation/polarization state of hepatic macrophages in terms of M1 and M2 phenotypes [4530], we inquired whether quercetin promoted phenotype switching of M1/M2 macrophages. We indeed found that quercetin supplementation markedly reduced transcripts of M1 marker genes such as TNFα, and NOS2 and enhanced those of M2 markers such as Arg-1, and Mrc1. This indicates that the beneficial effect of quercetin on hepatic inflammatory responses in the HFD-fed obese mice may be attributed to polarizing macrophages towards the M2 phenotype. Alternatively, hepatic lipid accumulation promotes macrophages/Kupffer cells activation, which initiates and perpetuates the inflammatory responses [2831]. Given that quercetin supplementation reduces hepatic lipid accumulation in the HFD-fed obese mice by enhancing mitochondrial oxidative capacity [32], the beneficial effect of quercetin may be partly associated with reduction of hepatosteatosis.
Quercetin reduces the production of lipopolysaccharide-stimulated inflammatory mediators such as nitric oxide, TNFα, IL-1β, and IL-6 in Raw 264.7 macrophages [2733], and does so by inducing HO-1 and disrupting activation of components of inflammatory signaling pathways such as phosphatidylinositol 3-kinase [2733]. Consistent with these findings, we found that quercetin enhanced expression of HO-1 in macrophages. Because HO-1 preferentially promotes the M2 phenotype [1014], we thought that quercetin might promote macrophage polarization by inducing HO-1. We confirmed this in experiments on co-cultures of lipid-laden hepatocytes/macrophages mimicking steatohepatitis. Furthermore, given that HO-1 expression is regulated by the activation of transcription factor Nrf2 [9], we inquired whether the quercetin action is mediated by Nrf2/HO-1 pathway. Indeed, we found that quercetin-induced alteration in M1/M2 phenotype switching: reduction of M1 marker and increase of M2 marker was completely blunted by Nrf2-deficient macrophages. More importantly, the effect of quercetin on the induction of HO-1 was completely disrupted in the LPS-treated peritoneal macrophages of the Nrf2-deficient mice. Our findings suggest that quercetin promotes macrophage M2 polarization and this effect is mediated, at least in part, by HO-1 induction, which is dependent on activation of the transcription factor Nrf2.
In conclusion, quercetin reduces obesity-induced steatohepatic inflammation and this is accompanied by macrophage phenotype switching towards the M2 phenotype dependent on the induction of HO-1 by the transcription factor Nrf2. Quercetin may be a useful dietary component to reduce obesity-induced hepatic inflammation.



 XML Download
XML Download