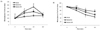INTRODUCTION
Diabetes mellitus and its complications are a cause for concern worldwide. The current global prevalence of diabetes mellitus is estimated to be 2.8 percent and is expected to reach about 7.7 percent by 2030 [1]. Thus, finding a treatment for this epidemic disease is crucial. Diabetes mellitus is an endocrine system disease that occurs metabolic disorders that induce to multiple organ damages. Causes of type 2 diabetes outbreak are complex and include insulin resistance in target tissues such as the skeletal muscle and adipose tissue [2], excessive hepatic glucose production, and decreased insulin secretion [3]. Although many oral antidiabetic drugs for type 2 diabetes such as metformin and thiazolidinediones [45] are available, they may have undesirable side effects [6]. Therefore, studies have been designed to identify natural products to treat diabetes.
Insulin regulates the metabolism of glucose by promoting glucose uptake from the blood to the skeletal muscles and adipose tissue and inhibits the production of glucose by the liver. Glucose uptake in the skeletal muscle is stimulated through activation of the AMPK-AS160-GLUT4 pathway. AMP-activated protein kinase (AMPK) is an important enhancer of insulin sensitivity. Phosphorylated AMPK activates AS160, thereby promoting glucose transporter 4 (GLUT4) translocation to the plasma membrane and glucose uptake in the cells [78]. The activation of this pathway enhances insulin sensitivity and is effective for the treatment of type 2 diabetes mellitus. Recent studies reported that some natural compounds such as flavonoids promote the activation of AMPK and glucose uptake in skeletal muscle [910]. The liver is also responsible for maintaining normal concentrations of blood glucose. In type 2 diabetes mellitus, increased gluconeogenesis was identified as the main cause of elevated glucose production and fasting hyperglycemia [11]. Therefore, regulation of gluconeogenic enzymes such as glucose-6-phosphatase (G6pase) and phosphoenolpyruvate carboxykinase (PEPCK) is important in hepatic glucose metabolism.
Gynura procumbens (G. procumbens) belongs to the family of Asteraceae. It recently received attention for its antidiabetic function as a folk remedy [12]. To date, studies showed that G. procumbens extract (GPE) presents anti-inflammatory, antioxidant, antibacterial, and antihypertensive effects [13141516]. The antidiabetic effect of G. procumbens was reported using streptozotocin-induced type 1 diabetic mice [17]. However, its effect on insulin secretion has been conflicting due to the differing response of various cell lines when treated with G. procumbens [18]. Especially, there is no report regarding the improvement of hyperglycemia and insulin sensitivity in type 2 diabetic mice. Therefore, we investigated the effect of GPE on hyperglycemia and insulin sensitivity in C57BL/Ksj-db/db mice, which show the characteristics of type 2 diabetes mellitus.
MATERIALS AND METHODS
Preparation of GPE
G. procumbens was collected from Uiwang Nae-Son province (Gyeonggi-do, Korea), washed in distilled water, freeze dried, and ground into a powder. G. procumbens powder was then extracted with water for 12 h three times at room temperature. GPE was concentrated in a rotary vacuum evaporator, freeze-dried to a powder, and then stored in a deep freezer (80℃). The yield of GPE was 5 ± 0.2%. GPE total flavonoid content was 10.33 ± 0.88 mg catechin equivalents (CE)/g dry weight (DW). It was reported that GPE contained 464.53 ± 1.81 µg/g DW of kaempferol, 251.10 ± 3.67 µg/g DW of myricetin, 135.87 ± 0.40 µg/g DW of quercetin [19].
Animals and diets
Male C57BL/KsJ-db/db mice were purchased from Japan SLC (Hamamatsu, Japan). Five-week-old db/db mice were fed a pelletized commercial chow diet for 2 weeks after arrival. They were subsequently randomly divided into 3 groups (n = 7 per group): db/db mice in the control group (DMC) were fed a standard semi-synthetic diet (AIN-93G) for 6 weeks, while animals in the other 2 groups were fed the AIN-93G diet supplemented with either rosiglitazone (0.005% w/w) or GPE (0.5%, w/w). Rosiglitazone was purchased from Sigma Chemical Co. (St. Louis, MO, USA). All mice were individually caged in a light-(12 h on/12 h off) and temperature-controlled room with food and water available ad libitum. At the end of the experimental period, mice were anesthetized with ether after a 12-h fast and blood samples were collected from the inferior vena cava to assess plasma biomarker levels. After collection of the blood samples, the liver was dissected and rinsed with physiological saline solution. All procedures were approved by the animal ethics committee of our university (PNU-2014-0619).
Blood glucose and glycosylated hemoglobin levels
The glucose concentration in the venous blood drawn from the tail vein was measured using a glucometer (Roche Diagnostics GmbH, Mannheim, Germany) every week after a 12-h fast. Anticoagulated whole-blood samples were hemolyzed and the concentration of glycosylated hemoglobin (HbA1c) was measured. Hba1c levels were determined by immunoturbidimetry.
Plasma insulin level
Blood samples from the inferior vena cava were collected into heparin-coated tubes. After centrifugation at 1,000 g for 15 min at 4℃, plasma was carefully removed from the sample. The levels of plasma insulin were determined by using an ELISA kit (Linco Research Inc., Billerica, MA, USA).
Homeostatic index of insulin resistance and quantitative insulin sensitivity check index
Homeostatic index of insulin resistance (HOMA-IR) and quantitative insulin sensitivity check index (QUICKI) were determined as surrogates of insulin sensitivity. HOMA-IR was calculated using the homeostasis model assessment with the following equation (Eq. (1)):
QUICKI was calculated using the inverse of the sum of the logarithms of fasting insulin and fasting glucose by using the following equation (Eq. (2)).
Intraperitoneal glucose and insulin tolerance tests
An intraperitoneal glucose tolerance test (IPGTT) was performed at 5 weeks after the start of the experimental diets. Following an overnight fast, mice were injected intraperitoneally with glucose (0.5 g/kg of body weight [BW]) and their blood glucose levels were determined in tail blood samples 0, 30, 60, and 120 min after glucose administration. Additionally, an intraperitoneal insulin tolerance test (IPITT) was performed during the last week of the experimental period. After an overnight fast, human insulin (0.33 U/kg of BW) was administered by intraperitoneal injection to the mice and blood samples were collected to determine glucose levels 0, 30, 60, and 120 min later.
Hepatic glycogen assay
Glycogen concentration was determined as previously described [20] with some modifications. Liver tissue was homogenized in 5 volumes of 30% (w/v) ice-cold KOH solution and dissolved in a boiling water bath (100℃) for 30 min. Glycogen was precipitated with ethanol and then pelleted, washed, and solubilized in distilled water. The resulting solution was treated with an anthrone reagent (2 g anthrone/L of 95% (v/v) H2SO4) and its absorbance was measured at 620 nm.
Plasma membrane fraction of skeletal muscles
Muscle tissue was placed in a buffer (5 mM sodium azide, 0.25 M sucrose, 0.1 mM phenylmethylsulfonyl fluoride, 10 mM NaHCO3, pH 7.0) at 4℃. Sub-fractionation of the muscle membranes was performed as previously described [21], using procedures modified from those described by Klip et al. [2223]. Dissected skeletal muscle was homogenized and centrifuged at 1,000 g for 10 min and the supernatant was collected and stored. The resulting pellet was resuspended in the buffer and re-homogenized in a glass homogenization tube. The supernatant was collected, combined with the first supernatant, and the combination was centrifuged at 9,000 g for 10 min. The resulting supernatant was then centrifuged at 190,000 g for 60 min. The membranes were subsequently applied to a discontinuous sucrose gradient containing 25, 30, and 35% sucrose (wt/vol) solutions and were centrifuged at 190,000 g for 16 h. Membranes were collected from the top of each sucrose gradient, resuspended in the buffer, pelleted by centrifugation at 190,000 g for 60 min, and resuspended in buffer.
Western blot
Western blot analysis was performed on tissue extracts from the liver and skeletal muscles. Liver and skeletal tissues were homogenized in ice-cold lysis buffer, centrifuged at 14,000 rpm, 4℃, for 15 min, and the supernatant were collected. Protein concentrations in the supernatants were measured using a protein assay kit (Bio-Rad, Hercules, CA, USA). Then, 30 g protein samples were separated on 12% resolving Tris-HCl gels. Separated proteins were transferred to nitrocellulose membranes. The membranes were blocked in 5% skim milk in Tris-buffered saline, 0.1% Tween-2 for 1 h at room temperature. Blocked membranes were incubated with antibodies overnight at 4C. Antibodies against phospho-AMPK (pAMPK), AMPK phospho-AS160 (Thr172), AS160, plasma membrane GLUT4 (PM-GLUT4), GLUT4, G6Pase, and PEPCK were purchased from Cell Signaling Technology (Beverly, MA, USA). The membranes were then washed and probed with a secondary antibody for 1 h at room temperature. Each antigen-antibody complex was visualized using ECL western blotting detection reagent and detected by chemiluminescence with LAS-1000 plus (FUJIFLIM, Tokyo, Japan). Band densities were determined by an image analyzer (Multi Gauge V3.1, FUJIFLIM Corporation, Valhalla, NY, USA) and normalized to -actin for total protein.
RESULTS
Body weight, food intake, and water intake
Body weights as well as food and water intake of all mice were observed during the experiment. Table 1 presents the monitored body weights as well as food and water intake of the mice during the 6 weeks of the experiment. Body weights of mice in db/db-control, db/db-RG, and db/db-GPE groups did not differ significantly at the beginning of the experiment. Body weights increased gradually during the 6-week period. At the end of the experiment, mice in the db/db-RG group exhibited significantly higher body weights than mice in the db/dbcontrol and db/db-GPE groups. The body weight was 46.18 ± 4.89, 45.80 ± 4.90, and 57.00 ± 2.12 g in the db/db-control, db/db-GPE, and db/db-RG group, respectively. The daily food intake was not significantly different between the db/dbcontrol, db/db-RG, and db/db-GPE groups. However, water intake was significantly higher in the db/db-control group mice than in the db/db-GPE group mice. Daily water intake was 28.57 ± 3.02 and 18.62 ± 1.97 mL/day in the db/db-control and db/db-GPE group mice, respectively.
Blood glucose and HbA1c levels
Fasting blood glucose levels in db/db-GPE group mice are shown in Fig. 1. At the beginning of the study, blood glucose levels were not significantly different between groups. However, fasting blood glucose levels of db/db-GPE mice after 4 weeks were significantly lower than those of the db/db-control mice. Blood glucose levels measured in db/db-control mice were elevated throughout the experiment, likely reflecting the progress of diabetes mellitus. As shown in Table 2, HbA1c values were 12.92 ± 0.31, 8.87 ± 1.09 and 7.45 ± 0.2 % in db/db-control, db/db-GPE and db/db-RG group, respectively. HbA1c values in the db/db-GPE mice were significantly lower than those measured in the db/db-control mice (P < 0.05).
Plasma insulin levels, HOMA-IR, and QUICKI
Plasma insulin levels measured in experimental animals are shown in Table 2. Plasma insulin levels were markedly lowered in the mice of the db/db-RG group (139.06 ± 21.96 pmol/L) compared to those of the mice in the db/db-control group (251.04 ± 25.24 pmol/L). Similarly, db/db-GPE mice exhibited significantly lowered plasma insulin levels than mice in the db/db-control group. HOMA-IR values were significantly lower in the db/db-GPE mice than in the db/db-control mice (P<0.05). Additionally, QUICKI values were significantly higher in the db/db-GPE mice than in the db/db control mice.
Intraperitoneal glucose and insulin tolerance tests
We performed IPGTT in db/db mice receiving GPE to evaluate GPE's ability to improve glucose tolerance. The results are presented as percentages of the measurement performed at the time of glucose injection (t = 0) and significant differences among db/db-control, db/db-RG and db/db-GPE group. As shown in Fig. 2 (A), blood glucose levels in the db/db-GPE mice peaked at 60 minutes after injection and recovered to levels close to the basal value at 120 min. A similar pattern was observed in mice from the db/db-RG group. IPITT was also performed to determine the effects of GPE supplementation on insulin tolerance. The results expressed as percentages of the measurement at the time of insulin injection (t = 0) and significant differences among db/db-control, db/db-RG, and db/db-GPE group are presented. As shown in Fig. 2 (B), db/db-GPE and db/db-RG mice exhibited a rapid reduction in blood glucose levels compared with the db/db-control mice, with blood glucose levels decreasing within 120 min of the insulin injection.
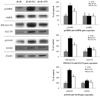
Expression of PM-GLUT4, GLUT4, pAMPK, AMPK, pAS160, and AS160 in the skeletal muscle
In this study, we studied the expression of PM-GLUT4, GLUT4, pAMPK, AMPK, pAS160, and AS160 in the skeletal muscle. RG supplementation, positive control, showed the highest PM-GLUT4, pAMPK, and pAS160 levels in the skeletal muscle. GPE supplementation also increased PM-GLUT4, pAMPK, and pAS160 levels in the skeletal muscle. As shown in Fig. 3, the expression of GLUT4 in the plasma membrane of the skeletal muscle significantly increased, reaching levels that were 2.2 and 3.3-fold higher in db/db-GPE and db/db-RG mice than those measured in db/db-control mice, respectively. No significant difference was observed in AMPK expression among db/db-RG, db/db-GPE and db/db-control mice. However, the phosphorylation levels of AMPK and AS160, two upstream regulators of GLUT4 translocation to the plasma membrane, were significantly increased by GPE supplementation. This means that GPE has the ability to stimulate glucose uptake through activation of the AMPK-AS160-GLUT4 pathway in the skeletal muscle.
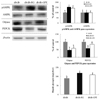
Expression of pAMPK and gluconeogenic enzymes, G6Pase and PEPCK, and glycogen levels in the liver
As shown in Fig. 4, pAMPK activation in the liver of db/db-GPE mice was significantly higher than that in the db/db-control mice. The expression of G6Pase in the liver significantly decreased in db/db-GPE mice, with 0.5-fold lower levels than those observed in the db/db-control mice. Hepatic glycogen levels in the db/db-RG mice were the highest with values of 119.33 ± 4.04 mg/g of liver. db/db-GPE mice were significantly higher than the levels in the db/db-control mice, with values of 109.67 ± 4.16 and 90.25 ± 4.75 mg/g of liver measured in db/db-GPE and db/db-control mice, respectively (Fig. 4).
DISCUSSION
Common characteristics of type 2 diabetes are chronic hyperglycemia caused by insulin resistance and increased gluconeogenesis in the liver [24]. Treatment for type 2 diabetes, aim at increasing insulin sensitivity and decreasing hyperglycemia. However, pharmacological treatments for type 2 diabetes exhibit a number of limitations, including side effects. Thus, in this study, we investigated the effect of a medicinal plant extract, GPE on insulin sensitivity and glucose metabolism using an animal model of type 2 diabetes, C57BL/KsJ-db/db mice.
The daily food intake was not significantly different among db/db-control, db/db-RG and db/db-GPE groups. The body weight was also not significantly different between db/dbcontrol and db/db-GPE groups. But it was significantly more increased in db/db-RG group than those measured in the db/db-control and db/db-GPE groups. Weight gain of db/db-RG mice was attributable to an increased in the adipose tissue mass. Some studies attributed weight gain to rosiglitazone treatment. RG may affect the differentiation of preadipocytes in humans [25]. This effect is markedly enhanced in subcutaneous fat [26].
Water intake was significantly lower in db/db-GPE mice than in db/db-control mice. The increase in water intake shows the state of polydipsia, one of the typical characteristics of diabetes [27]. In this study, GPE supplementation in db/db mice ameliorated the symptoms of diabetes, reducing water intake.
After 4 weeks of experimental diet, fasting blood glucose levels in the db/db-GPE mice were significantly lowered when compared with those of the db/db-control mice. Hyperglycemia is associated with the development of type 2 diabetes. Therefore, controlling blood glucose levels is important for preventing diabetes mellitus and diabetic complications [28]. In addition, GPE treatment significantly lowered HbA1c levels. HbA1c is generally used as an accurate index of long-term blood glucose regulation and is a simple way to investigate the glycemic control in diabetes [29]. Lowering HbA1c levels in diabetes mellitus can effectively prevent the risk of complications such as retinopathy and nephropathy [30]. The flavonoids present in GPE may be responsible for the reduction of hyperglycemia. Components of GPE such as quercetin, kaempferol, and myricetin have antidiabetic effects by lowering blood glucose levels [3132]. This suggests that GPE supplementation has positive effects in controlling hyperglycemia in type 2 diabetes.
This study showed that GPE supplementation could improve plasma insulin levels. The levels of plasma insulin in db/dbcontrol mice were higher than those in db/db-GPE mice, suggesting that db/db-control mice still exhibited hyperinsulinemia. db/db mice generally show an initial phase of hyperinsulinemia, while insulinopenia could develop with age [33]. It is a common feature exhibited in the late stage of type 2 diabetes. In this study, the plasma insulin levels in db/db-GPE mice were significantly lower than those measured in db/dbcontrol mice. The mice in the db/db-control group still displayed hyperinsulinemia. On the other hand, db/db-RG and db/db-GPE mice showed reduced hyperinsulinemia. Hyperinsulinemia is caused by insulin resistance, which is a characteristic of the early stage of type 2 diabetes [34]. When the insulin reaction does not function properly in target tissues, more insulin is needed, resulting in hyperinsulinemia. Some studies reported that flavonoids have the ability to improve insulin resistance, thereby, reducing hyperinsulinemia and preventing metabolic symptom [932]. HOMA-IR and QUICKI calculated from fasting insulin and glucose levels are the commonly used measures of insulin sensitivity. HOMA-IR is an index of insulin resistance, which increases with increasing insulin resistance [35]. QUICKI is an index of insulin sensitivity, with values that decrease with increasing insulin resistance [36]. In this study, HOMA-IR was significantly lowered in mice of the db/db-GPE group compared with the values measured in db/db-control mice. These results suggest that GPE supplementation might improve insulin sensitivity.
In type 2 diabetes mellitus, hyperglycemia presents as a reduction in the insulin-mediated blood glucose disposal. Our results showed that GPE supplementation may improve both glucose and insulin tolerance. The results of IPGTT showed improved glucose levels in db/db-GPE and db/db-RG mice by lowering blood glucose levels to the basal level, compared to the levels measured in db/db-control mice. IPITT was also improved in mice supplemented with GPE and RG compared to that of mice in the db/db-control group. These results suggest that GPE and RG might act as insulin sensitizers. RG is an antidiabetic drug and was used as the positive control for GPE in this study. RG works as an insulin sensitizer by binding to the PPAR receptor in fat cells making the cells more responsive to insulin [26]. db/db-GPE mice also showed improved insulin sensitivity, which is important for lowering blood glucose levels, even if the effect of GPE was lower than that of RG.
Enhancing insulin sensitivity is very important in the treatment of diabetes. We investigated the expression of genes related to insulin sensitivity using western blotting. Glucose uptake is regulated by insulin binding to the insulin receptor on the cell surface via GLUT4 [37]. GLUT4 is the key glucose transporter in response to insulin in the skeletal muscle [38]. The action of insulin on glucose transport in the skeletal muscle is also affected by AMPK, suggesting that AMPK can be a therapeutic target for the treatment of type 2 diabetes, especially, with insulin resistance [39]. Through an insulin-dependent mechanism, increased activation of AMPK increases GLUT4 translocation to the plasma membrane [40]. Dietary GPE supplementation in db/db mice significantly increased AMPK activation. This resulted in the increased expression of GLUT4 in the plasma membrane, which contributed to the improvement in insulin sensitivity. GLUT4 expression in the plasma membrane was 2.2 and 3.3-fold higher in db/db-GPE and db/db-RG mice than those measured in db/db-control mice, respectively. Although GLUT4 expression in the plasma membrane of db/db-GPE mice was lower than that in db/db-RG mice, GLUT4 expression in the plasma membrane in db/db-GPE mice was significantly higher than that in db/db-control mice. GPE supplementation also increased the expression of phosphorylated AS160, suggesting that it may stimulate glucose uptake in the skeletal muscles and reduce hyperglycemia. AS160 is a major mediator of GLUT4 translocation and glucose uptake in the skeletal muscle cells [40]. When AS160 is activated, GLUT4 translocation from the intracellular pool to the plasma membrane is promoted [6]. Through the upregulation of the AMPK-AS160-GLUT4 pathway, glucose uptake and utilization in the skeletal muscle are enhanced. GPE contains flavonoids such as quercetin and 3-O-glycosides of kaempferol. These flavonoids stimulate AMPK activation and GLUT4 translocation in the skeletal muscle, thereby, increasing basal glucose uptake in the skeletal muscle [41].
The liver plays a significant role in regulating glucose production from gluconeogenesis or glycogenolysis. In the postprandial state, insulin stimulates the liver to take up more glucose from the blood and synthesize glycogen. In the starving state, the liver produces more glucose to maintain blood glucose levels. However, under the diabetic state, hepatic gluconeogenesis is abnormally enhanced and glycogen synthesis is inhibited [42]. In this study, the expressions of G6pase and PEPCK, the key gluconeogenic enzymes, was significantly decreased by GPE supplementation, while hepatic glycogen levels were significantly increased. These results show that GPE might decrease blood glucose levels by inhibiting gluconeogenesis and stimulating hepatic glycogen synthesis. In the liver, AMPK also plays an important role in the control of hepatic glucose metabolism. AMPK activation in the liver results in the downregulation of the expression of gluconeogenic enzymes [43]. Therefore, increased AMPK level in the liver is associated with an amelioration of glucose production [44]. Suppressing hepatic glucose output and several enzymes that regulate gluconeogenic pathways in the liver might be useful for the treatment of diabetes [45]. The main enzymes that control gluconeogenesis and glucose metabolism in the liver are G6Pase and PEPCK [46]. By downregulating the activity of these enzymes, glucose production can be decreased, resulting lowered blood glucose level. Our results indicate that pAMPK expression was increased in GPE-supplemented mice, while the expression of gluconeogenic enzymes was downregulated. This means that GPE may have the ability to reduce hepatic gluconeogenesis. These effects may be attributed to the flavonoids present in GPE. Quercetin is a known nonspecific inhibitor of glycogen phosphorylase, the enzyme that catalyzes the rate-limiting step of glycogenolysis by releasing glucose-1-phosphate from the terminal α-1,4-glycosidic bond [4748]. Flavonoids such as quercetin and kaempferol activate AMPK and significantly reduce G6Pase and PEPCK in hepatocytes and may reduce hepatic glucose production [3249].
In conclusion, the present study showed that GPE improves hyperglycemia by enhancing insulin sensitivity in the skeletal muscle and suppressing glucose production in the liver of type 2 diabetic db/db mice. GPE increased the expression of PM-GLUT4 in the skeletal muscle through pAMPK and pAS160 activation and reduced hepatic gluconeogenesis. These results suggest that GPE might be beneficial in type 2 diabetes and useful as an antidiabetic supplement.