

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Adipose tissue is a crucial organ that regulates whole-body insulin sensitivity and energy homeostasis [1]. It is well-accepted that adipocytes not only store excessive fat but also have endocrine functions by secreting hormones and inflammation-related adipokines [23]. One of the pathologies of insulin resistance (IR) and other obesity complications is the accumulation of circulating bone marrow-derived inflammatory macrophages (BMDM) in adipose tissue, resulting in adipocyte death that appears to increase in obese mice and humans [456]. Obesity-associated adipocyte death is linked to recruitment of BMDM, aggregation of crown-like structures (CLS), and pro-inflammatory gene expression [7]. Thus, chronic inflammation in adipose tissue plays an important role in development of obesity-related IR [8].
Due to the high costs and potentially adverse side effects associated with anti-obesity and anti-inflammatory drugs as long-term treatments, screening of natural sources with reducing effects on inflammation as well as minimal side effects has drawn much attention. Numerous natural products such as crude extracts and isolated compounds from plants appear to be applicable as obesity-induced inflammation treatments, including genistein [9], kaempferol [10], quercetin [11], daidzein [10], fermented soybean paste (Doenjang) [12], green tea [13], garlic compounds [14], and conjugated linoleic acid (CLA) [15].
Seaweeds, especially brown seaweeds, are a common food ingredient in Eastern Asian countries such as Korea and are a rich source of proteins, vitamins, essential minerals, fiber, polyunsaturated fatty acids, and various bioactive components [16]. Due to their potential beneficial activities such as anti-inflammatory [17] and anti-diabetic effects [1819], seaweeds have been intensively studied. The compounds included in seaweed extracts can vary depending on the solvent used for preparation. In general, ethanol extracts of seaweeds are comprised of 70-80% polyphenol compounds and 20-30% polysaccharides, whereas water extracts of seaweeds contain very little amounts of polyphenols and > 90% polysaccharides [20]. Most previous studies on the anti-inflammatory and anti-diabetic effects of seaweeds have been carried out by extraction using various solvents rather than consumption of seaweeds themselves. Furthermore, there is not much information about the long-term beneficial effects of seaweeds supplemented with high-fat diet (HFD), especially their regulatory effects on inflammation in white adipose tissue and systemic IR. In the current study, we hypothesized that brown seaweed supplementation alters complications induced by long-term HFD feeding. To test this hypothesis, we evaluated whether or not supplementation with brown seaweeds such as Undaria Pinnatifida (UP), Laminaria Japonica (LJ), Sargassum Fulvellum (SF), and Hizikia Fusiforme (HF) inhibits obesity-associated inflammation in adipose tissue, bone-marrow derived immune cells, and systemic IR in a HFD-induced obese mouse model for 16 weeks.
MATERIAL AND METHODS
Animals and diets
Male C57BL/6N mice at 5 weeks of age were purchased from Orient Bio Co. (Korea) and housed individually at the Natural Science Animal Facility of Jeju National University. Animals were maintained in a temperature (21 ± 2℃) and humidity (50 ± 20%) controlled room with a 12 h dark-light cycle. Ethical treatment of animals was assured by the Jeju National University Institutional Animal Care and Use Committee (#2014-0004). After 1 week of acclimation, mice were randomly divided into 6 groups (n = 6 mice/group) that received one of six pelleted diets for 16 weeks (Research Diets, NJ, USA) as follows: low-fat diet (LFD, 10% kcal% fat (D12450B)), high-fat diet (60% kcal% fat (D12492), HFD), or modified high-fat diet supplemented with 5% freeze-dried UP, LJ, SF, or HF (Table 1). Four types of brown seaweeds were purchased from a traditional market in Jeju from December 2014 to March 2015. The powder of freeze-dried seaweeds (UP, LJ, SF, and HF) were prepared as described elsewhere [21] and sent to Research Diets to produce the modified HFDs. Fresh diet was provided at least twice per week to minimize oxidization of fats and stored at -20℃.
IR measured by insulin tolerance test (ITT)
ITT were performed to measure IR induced by HFDs on food-deprived (6 h), non-anesthetized mice as described previously [22]. Glucose measurements were obtained from whole tail vein blood using an automated glucometer (OneTouch Select simple, Switzerland) at baseline as well as 30, 45, 60, and 90 min following intraperitoneal injection of insulin (0.75 mU/kg, form of bovine pancreas, Cat#: I1882, Sigma, USA). Glucose concentrations following insulin injection were calculated as a proportion of the value at baseline. Glucose area under the curve (AUC) was calculated using these values and analyzed by ANOVA.
Adipose tissue histology and CLS
Mice were killed by cervical dislocation following CO2 narcosis. Subcutaneous (SQ) adipose tissue and gonadal adipose tissue were fixed in 10% formaldehyde overnight, embedded in paraffin, sectioned, and stained with hematoxylin and eosin (H&E) [47]. Digital images were acquired with an Olympus BX51 light microscope (Tokyo, Japan). Adipocyte death was quantified by identification of CLS within the sections of gonadal adipose tissue. Adipocyte size was calculated by Image J software (Sun microsystems, Santa Clara, CA) [723].
Isolation of BMD cells
After 16 weeks of feeding, a representative weight-matched mouse from each experimental group was chosen, and bone marrow cells were isolated from femoral and tibial bone marrow as described previously [2324]. Briefly, L929 cells were purchased from Korea Cell Line Bank (Seoul, Korea) and cultured in DMEM (Gibco BRL, MD, USA) containing 10% fetal bovine serum (FBS, Gibco BRL, MD, USA) and 1% penicillin-streptomycin (Penn/Strep). L929 cells were plated in a 75-cm flask (5×105 cells) containing 55 mL of L929 medium for 7 days to generate M-CSF (macrophage colony-stimulating factor (M-CSF)). The isolated bone marrow cells were differentiated into macrophages in BMDM medium consisting of 50% DMEM, 30% L929-conditioned medium, 20% FBS, and 5 mg/mL of Penn/Strep in a 10-cm bacterial dish. BMDM cells were plated in 6-well plates and ready for LPS stimulation (100 ng/mL) on the following day.
Inflammation cytokine detection by ELISA
Mouse adiponectin, IL-1β, IL-6, leptin, and TNFα were measured by a Quantikine system from R&D systems (MN, USA) according to the manufacturer's instructions. BMDM culture media was collected at the end of 24 h of LPS stimulation, and serum of mice fed the assigned experimental diet was collected by cardiac puncture. All samples and standards were measured in duplicate.
Statistical analysis
Data are presented as mean ± standard error of the mean (SEM). Statistical differences were determined by t-test or one-way ANOVA followed by Duncan's multiple range tests was employed to assist statistical significance by using SPSS software (ver. 21.0, SPSS Inc., USA). Significance was set at P ≤ 0.05.
RESULTS
Effects of seaweeds on body weight, adipose tissue weight, and inflammatory cytokines in C57BL/6N mice fed HFD for 16 weeks
The 6-week-old male C57BL/6N mice were fed LFD, HFD, HFD + UP, HFD + LJ, HFD + SF, or HFD + HF for 16 weeks. Body weight (BW) of the LFD group showed significant difference from HFD groups starting at week 5 (Fig. 1). At week 8, BWs of the HFD + LF and HFD + HF groups were slightly lighter than that of the HFD group (P = 0.1), although this trend disappeared as feeding progressed. Interestingly, BW of the HFD + UP group (48.32 ± 2.06 g) was heavier than the rest of the HFD groups (44.67 ± 1.47 g) partially due to increased subcutaneous adipose tissue weight (HFD vs. HFD + UP; 3.49 ± 0.42 g vs. 4.07 ± 0.54 g, P = 0.02) (Table 2). Eventually, all HFD groups became obese at the end of 16 weeks of feeding with no statistical difference between the HFD only and HFD plus seaweed groups. Additionally, food intake among the experimental groups was not significantly different (data not shown). Consistent with a lack of difference in body weight among the HFD groups, gonadal fat pad masses were similar among the HFD groups. Plasma concentrations of adiponectin in the HFD + UP group were significantly lower than those of the HFD group, whereas plasma concentrations of leptin were significantly increased in the HFD group compared to the LFD group. Supplementation of LJ, SF, or HF with HFD reduced plasma concentrations of leptin compared to the effects of HFD alone. However, plasma concentrations of IL-6 were not altered by supplementation of HFD with seaweeds in general. Thus, obese mice fed HFD + seaweed showed reduced levels of circulating cytokines compared with mice fed HFD.
Effects of seaweed consumption on HFD-induced IR
ITT were performed after 14 weeks of experimental diets to assess the effects of seaweeds on HFD-induced IR (Fig. 2A). The AUC of the HFD group was significantly greater than that of the LFD group. Notably, mice fed HFD + LJ were significantly less insulin resistant than mice fed HFD (Fig. 2B). Consistent with the observed improvement of insulin sensitivity in mice fed HFD + LJ, blood glucose (BG) concentrations were significantly lower in HFD + LJ mice (175.8 ± 3.35 mg/dL) compared with HFD mice (227.6 ± 1.46 mg/dL) (Table 2). Additionally, mice fed HFD supplemented with SF or HF also showed lowered BG concentrations compared to mice fed HFD, although their AUC was not reduced compared to the HFD group. In fact, BG concentrations of mice fed HFD + LJ or HFD + HF did not differ from those of mice fed LFD, and BG concentrations of mice fed HFD + UP were as high as those of mice fed HFD only.
Effects of seaweed consumption on CLS formation and size of gonadal adipose tissue in C57BL/6N mice fed HFD
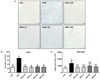
To investigate whether or not seaweed consumption can prevent inflammation in adipose tissue of mice fed HFD for 16 weeks, gonadal and subcutaneous adipose tissues were subjected to H&E staining to evaluate the size and number of dead adipocytes as well as the number of CLS (Fig. 3A and B). The frequency of dead adipocytes was about 3-fold greater in mice fed HFD than in mice fed LFD, whereas the number of CLS was dramatically reduced in gonadal adipose tissues of all mice supplemented with HFD and four types of seaweeds compared to mice fed HFD only. It has been reported that the insulin responsiveness of adipose tissue depends on the size of adipocytes. That is, larger adipose cells are less sensitive to insulin [25]. We measured the size of adipocytes from gonadal adipose tissues of obese mice fed HFDs containing seaweeds (Fig. 3C). Adipocyte size in obese mice fed seaweeds with HFDs was smaller than that of obese mice fed HFD only, although the difference was not statistically significant.
Effects of seaweeds on inflammatory response in BMDM stimulated with LPS
BMD immune cells play a significant role in inflammation associated with obesity [2627]. It was previously reported that increased concentrations of fatty acids and endotoxins such as lipopolysaccharide (LPS) induce inflammatory signaling via toll-like receptor (TLR) 4 activation [28]. To evaluate the inflammatory responses of BMD immune cells from mice fed LFD, HFD, or HFDs supplemented with four types of brown seaweeds, we isolated BM cells and differentiated them into BMDM treated with or without the TLR agonist LPS for 24 h. In response to LPS treatment, BMDM from mice fed HFD showed significantly greater IL-1β expression compared with BMDM isolated from mice fed LFD, HFD + UP, HFD + LJ, HFD + SF, or HFD + HF (Table 3). IL-6 production by LPS challenge in BMDM from mice fed HFD + LJ was significantly lower than that in BMDM from mice fed HFD for 16 weeks, whereas supplementation of HFD with UP, SF, or HF did not inhibit IL-6 production by LPS challenge in BMDM.
DISCUSSION
Obesity has become a global health problem [29]. Excessive adipogenesis in the body along with white adipose tissue causes obesity, which results in an increased risk of many serious diseases such as type II diabetes, hypertension, and heart disease [30]. Numerous chemokines and cytokines secreted from adipose tissue are major contributors to chronic and systemic inflammation in obesity. Moreover, chronic inflammation in adipose tissue plays a crucial role in the development of obesity-related IR, which is highly associated with accumulation of circulating BMDM [45]. That is, not only adipocytes but also macrophages are able to produce and secrete adipokines [31]. Anti-inflammatory and anti-diabetic effects of brown seaweeds and their extracts have been reported in previous studies [171819]. However, this study particularly evaluated the long-term beneficial effects of brown seaweed supplementation on obesity, insulin sensitivity, and chronic inflammation using a HFD-induced obese mouse model. Our results demonstrate that supplementation of HFD with 5% brown seaweed, especially Lamina japonica (LJ), can inhibit the inflammatory events in adipose tissue that promote obesity-associated IR in the absence of any reduction of adipose depot mass.
Consistent with our results of improved insulin sensitivity by LJ supplementation in the present study, Li et al. [32] demonstrated that Laminaria japonica polysaccharides (LJPS) had an anti-diabetic effect by significantly lowering fasting blood glucose levels and increasing serum insulin levels after 2 weeks of LJPS treatment (0.75g/kg body weight) in a type 2 diabetes mellitus mouse model. On the other hand, a prior study examining the anti-obesity and anti-diabetic effects of Laminaria japonica Areshoung ethanol extract (LE) on diet-induced obesity for 6 weeks reported that LE reduced adipocyte mass, surface area of epididymal adipose tissue, and food intake in rats [33]. It should be noted that the HFD used in this previous study consisted of 45% calories from fat, whereas the HFD used in the current study consisted of 60% calories from fat. Moreover, feeding duration seems to have an impact on the anti-obesity effect of seaweed supplementation with HFD [33]. In fact, we observed that BWs of mice fed HFD+ LF and HFD + HF were slightly lighter than the BW of mice fed HFD (P = 0.1) between weeks 8 and 11, although this trend eventually disappeared as feeding continued until week 16. Thus, dietary seaweed consumption reduced body weight gain during short-term HFD feeding but could not overcome prolonged HFD feeding in C57BL/6N mice.
Additionally solvents applied to seaweed extracts have crucial effects on compounds in those particular extracts. For example, Kang et al. demonstrated an anti-obesity effect for water extract from brown seaweed, Petalonia binghamiae (PB), both in vitro and in vivo [20], which was in contrast to their previous result of ethanol extract from PB increasing adipogenesis in 3T3-L1 cells [34]. The differing effects of these two PB extracts can be attributed to their major components; water extract is comprised of 25.39 mg/g of polyphenols and 8.3 mg/g of polysaccharides, whereas ethanol extract contains 3.96 mg/g of polyphenols and 424.3 mg/g of polysaccharides. In fact, seaweeds are a rich source of proteins, vitamins, essential minerals, fiber, polyunsaturated fatty acids, and various bioactive components in addition to polyphenols and polysaccharides [16]. Therefore, the observations in the present study may be due to combined effects of various compounds in freeze-dried seaweeds, thus warranting further studies.
Unexpectedly, mice fed HFD + UP had significantly greater subcutaneous fat mass and showed no reduction of plasma leptin or fasting blood glucose level compared to mice fed HFD only. In contrast to our findings, Park et al. [35] showed that UP ethanol extract (0.69%), fucoxanthin-rich extract, lowers the amount of visceral fat, adipocyte size, fasting blood glucose concentration, plasma insulin concentration, and IR index in C57BL/6J mice after 9 weeks of feeding compared to the HF control group. These contrasting results might be due to the types of UP supplementation (i.e., UP ethanol extracts vs. UP freeze-dried powder), duration of feeding (i.e., 9 weeks vs. 16 weeks), and types of HFD used. Although supplementation of UP did not protect HFD-fed mice from obesity in the current study, the number of CLS in gonadal adipose tissue (Fig. 3) was significantly reduced compared to the HFD group. In addition, BMDM from mice fed HFD+UP for 16 weeks produced significantly less IL-1β upon LPS stimulation compared with BMDM from mice fed HFD only (Table 3), suggesting that UP supplementation could provide anti-inflammatory and antidiabetic effects against long-term HFD-associated complications.
The observed improvement of systemic insulin sensitivity upon LJ supplementation with HFD in the current study appears to be partially due to significantly reduced adipocyte death in gonadal adipose tissue as well as less priming of BMDM. Previous studies reported that the pro-inflammatory actions of recruited BMD immune cells play a crucial role in adipose tissue inflammation [4826]. That is, accumulation of pro-inflammatory macrophages in adipose tissue is strongly associated with the metabolic complications of obesity [7]. Consequently, these macrophages selectively localize to CLS surrounding dead adipocytes [4], which we observed as significantly reduced in all seaweed-supplemented HFD groups compared to the HFD group (Fig. 3). Moreover, the size of adipocytes affects insulin responsiveness of adipose tissue, as larger adipose cells are less sensitive to insulin [25]. Adipocyte size of obese mice fed seaweeds with HFDs was smaller than that of obese mice fed HFD only, although the difference was not statistically significant.
In order to evaluate the pro-inflammatory actions of BMDM, we isolated BM cells from a weight-matched representative mouse fed the assigned diets for 16 weeks and differentiated them into BMDM. As a result, IL-1β production induced by LPS stimulation of BMDM from mice fed HFD + UP, HFD + LJ, HFD + SF, and HFD + HF was significantly lower than IL-1β production by BMDM from mice fed HFD only (Table 3). It was previously shown that IL-1 receptor antagonist binds to IL-1 receptor in order to block activation of IL-1β, which consequently improves glycemia and β-cell function and decreases systemic inflammatory markers [36]. Thus, less primed BMDM may partially contribute to the significantly reduced number of CLS in gonadal adipose tissue of mice fed HFD supplemented with 5% brown seaweeds in the current study.
In conclusion, dietary consumption of brown seaweeds led to attenuated IR by reducing adipose tissue inflammation in a long-term HFD-induced obese animal model. Especially, mice fed LJ with HFD showed improved IR and reduced circulating pro-inflammatory cytokines despite being obese. Additionally, supplementation of HFD with UP, SF, and HF reduced inflammation in the absence of decreased adipose depot mass and improvement of IR. Collectively, dietary brown seaweeds can provide metabolic benefits to overcome HFD-induced obesity-associated complications.





 XML Download
XML Download