

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Diabetes mellitus (DM) is a chronic disease, characterized by hyperglycemia. DM leads to various complications, such as retinopathy, nephropathy, and neuropathy. It has become a major public health problem throughout the world [1]. Diabetic nephropathy (DN) is considered one of the most serious microvascular complications of diabetes. It is the leading cause of end-stage renal disease and accounts for high morbidity and mortality in diabetic patients [2].
Hyperglycemia induced reactive oxygen species (ROS) production in diabetes may inflict direct and indirect damage through oxidative stress and inflammatory response that trigger cellular dysfunction and progression of DN [3]. To protect against the toxic effect of ROS, the anti-oxidative defense system operates chiefly via anti-oxidant enzymes, including heme oxygenase-1 (HO-1), catalase, glutathione peroxidase (GPx), Glutathione S-transferase (GST), and superoxide dismutase isoforms (SODs) [4]. These enzymes play key roles in the detoxification of superoxide radicals with respect to hydrogen peroxide and water in different cellular compartments [5]. Therefore, regulation of anti-oxidant enzymes would be a desirable target for DN prevention. It is of interest to identify the relationship between an impaired antioxidant system and DN.
Both oxidative stress and chronic inflammation contribute to the development and progression of diabetes and its complications. ROS may initiate and exaggerate inflammatory response owing to their ability to stimulate a number of genes regulating the inflammatory-signaling cascades. Abnormalities in renal metabolism, such as increases in diacylglycerols, non-enzymatic glycation, advanced glycation products and protein kinase activities as well as oxidative stress, evidently contribute to the development of DN. Among them, PKC plays a primary role in many cellular functions and has effects on many signal transduction pathways [6]. Especially, PKCβ is well known to accelerate renal dysfunction by increases in transforming growth factor-beta I (TGF-β), vascular endothelial growth factor (VEGF), connective tissue growth factor (CTGF), type IV collagen, and fibronectin in the renal glomeruli [7]. An increase in TGF-β1 promotes renal fibrosis by regulation of mesangial cell proliferation and extracellular matrix (ECM) synthesis. TGF-β1 stimulates the synthesis of matrix molecules, such as type I and IV collagens, fibronectin, laminin, and proteoglycans. Enhanced glomerular expression of TGF-β1 has been reported in DN patients [8].
Moreover, the activation of PKC is known to contribute to the intracellular pathways responsible for activation of nuclear factor kappa B (NFκB), a redox sensitive transcription factor [9]. NFκB activated by oxidative stress regulates the host immunity and inflammatory responses [10] and controls cellular proliferation and apoptosis [11]. NFκB activation triggers expression of a variety of inflammatory molecules, including interleukin-1 beta (IL-1β), interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), monocyte chemotactic protein-1 (MCP-1), intercellular adhesion molecule (ICAM), and TGF-β1 [12]. Recent reports suggest that increased ICAM-1 and TGF-β1 by NFκB activation cause renal diseases leading to excessive fibronectin production and extracellular matrix (ECM) accumulation in DN [13]. Therefore, modulation of NFκB signaling via PKC by improving the antioxidant defense system may ameliorate DN.
Dietary anti-oxidants have been shown to protect various organs, including the liver [14], retina [15], and kidney [16], against excessive ROS production. Among anti-oxidants, vitamin E is known as a primary lipid-soluble chain-breaking antioxidant. It has been well described as having several beneficial effects on the prevention of metabolic disorders, such as cardiovascular disease (CVD) [17], obesity [18], cancer [19], and diabetes [20]. Alpha-tocopherol (AT), the major tocopherol in mammalian tissue, possesses the most biological activity with antioxidant and anti-inflammatory properties [2122] whereas γ-Tocopherol (GT), the most common form of vitamin E in the US diets [23], may be a more potent pro-apoptotic agent than AT in trapping ROS and reactive nitrogen species (RNS) in colon cancer [24]. However, there is limited research focusing on the effects of tocopherols, especially GT on diabetes induced kidney damage. Moreover, it is often ignored that diabetes and diabetic complications would be treated differently according to fasting blood glucose levels in terms of kinds or doses of antioxidants treated. In the present study, we investigated whether short-term supplementations of AT and GT protect hyperglycemia induced acute kidney damage as an initial stage of DN, depending on fasting blood glucose levels through modulation of oxidative stress, inflammation, and pre-fibrosis in diabetes.
Go to : 
MATERIALS AND METHODS
Animals
4 weeks old male ICR mice were purchased from the Nara Bio (Gyeonggi-do, South Korea). During the experiments, the mice were individually housed in cages and access to food and water ad libitum. The environment was controlled in term of light (12h dark/light cycle), humidity (50 ± 1%), and room temperature (22 ± 1℃). All animal procedures were performed in accordance with the guidelines issued by Kyung Hee University for the care and use of laboratory animals (KHUASP (SE)-14-007). One week after their arrival, diabetes was induced by intraperitoneal (i.p.) injection of 150 mg/kg alloxan monohydrate (Sigma-Aldrich, St Louis, MO, USA) dissolved in 0.9% NaCl. Non-diabetic control mice were injected with only 0.9% NaCl. After one week, fasting blood glucose was measured from the mouse tail vein using the One-Touch blood glucose meter (LifeScan Inc., Milpitas, USA). Once fasting blood glucose levels reached above 250 mg/dl, mice were considered as diabetes and treated for this study.
Experimental design
Diabetic mice were divided into two groups according to fasting blood glucose (FBG) levels; mild FBG (m-DMC; 250 mg/dl ≤ FBG ≤ 450 mg/dl), and severe FBG (s-DMC; 450 mg/dl < FBG). M-DMC and s-DMC were subdivided into four groups depending on AT or GT supplementation. Dosage of each tocopherol was referred to Jiang et al.'s study [25]. Mice were consequently divided into seven groups as follows: CON, non-diabetic control mice were administrated with AT stripped corn oil (TSCO, MP biomedicals Inc, Solon, OH, USA); m-DMC, diabetic-control mice with mild FBG levels (250 mg/dl ≤ FBG ≤ 450 mg/dl) which were administrated with a TSCO; m-AT, diabetic mice with mild FBG levels which were administrated with AT (35 mg/kg, Acros organics, Geel, Belgium) dissolved in a TSCO; m-GT, diabetic mice with mild FBG levels which were administrated with GT (35 mg/kg, Acros organics) dissolved in a TSCO; s-DMC, diabetic mice with severe FBG levels (450 mg/dl < FBG) which were administrated with a TSCO; s-AT, diabetic mice with severe FBG levels which were administrated with AT (35 mg/kg, Acros organics) dissolved in a TSCO; s-GT, diabetic mice with severe FBG levels which were administrated with GT (35 mg/kg, Acros organics) dissolved in a TSCO. All mice were fed a modified AIN-76A rodent diet (Research Diets, New Brunswick, NJ, USA). As previously reported [1626], treatments were administered by gavage daily for 2 weeks (at least 6 mice in each group). During the experiment period, body weights, food consumption and fasting blood glucose levels were measured once a week. Mice were fasted 8 hours before sacrifice, and anesthetized by isoflurane (Ilsung pharmaceutical, Seoul, South Korea). Blood and kidney were collected. Blood was collected by cardiac puncture in heparin (Sigma-Aldrich, St. Louis, MO., USA) and isolated by centrifugation at 500 X g for 15min at 4℃. The kidney was cleaned with saline and frozen immediately in liquid nitrogen. All samples were stored at -80℃ until they were analyzed.
Measurement of plasma insulin
We determined the levels of plasma insulin by insulin immunoassay kit (Morinaga Institute of Biological Science, Yokohama, Japan), according to the manufacturer's protocol.
Renal function tests
Renal function was measured by blood urea nitrogen (BUN) and plasma creatinine. Specimens were examined by a commercially available kit (Asan pharmaceutical, Seoul, South Korea) and then, concentrations were determined at 580nm using an ELISA reader (BIO-TEK instruments, Winooski, VT, USA). Plasma creatinine levels were examined by creatinine assay kit (Bio-Clinical System, Gyeonggi-do, South Korea) according to the manufacturer's protocol. Concentrations were determined at 515nm using an ELSA reader.
Measurement of lipid peroxidation
Lipid peroxidation levels were measured by levels of malondialdehyde (MDA) using thiobarbituric acid method [27]. Kidney tissues were homogenized with 0.15M KCl (Duksan, Gyeonggi-do, South Korea) solution. 0.2ml of homogenous kidney tissue was added to 0.2ml of 8.1% SDS (Sigma-Aldrich, St. Louis, MO., USA) and incubated at room temperature for 10min. 3ml of 20% acetic acid (Duksan, Gyeonggi-do, South Korea)-0.8% TBA mixture (Lancaster Synthesis, Morecambe, England) (1:1, v/v) and 0.6ml of distilled water were added. The reaction mixture was heated in a water bath at 95℃ for 1h, and then cooled by tap water immediately. To each tube, 1ml distilled water and 5ml of n-butanol (Duksan, Gyeonggi-do, South Korea) and pyridine (Duksan, Gyeonggi-do, south Korea) (15:1, v/v) were added and shaken using a vortex. After centrifuging at 4,000 rpm for 10 min, the pink supernatant was measured at 532 nm using ELISA reader (BIO-TEK instruments, Winooski, VT, USA) with 1,1,3,3-tetramethoxypropane (Sigma-Aldrich) as a standard.
Western blot analysis
0.1 g of kidney tissues were homogenized in a lysis buffer (containing Trizma base, 150 mM NaCl2, pH7.5, 10% NP40, 10% Na-deoxycholate, 100 mM EDTA 10% SDS) with a mixture of protease inhibitors (1:200 v/v, Sigma-Aldrich) and centrifuged at 14,000 rpm for 30min at 4℃. The protein supernatants were used for western blot analysis. For the preparation of nuclear fraction, kidney tissues were homogenized in buffer A (0.6% Nonidet P40, 150 mMNaCl, 10 mM HEPES (pH 7.9), 1 mM EDTA, 0.5 mM PMSF, leupeptin, pepstatin and aprotinin). Next the homogenization procedure, samples were centrifuged 1000 × g at 4℃ for 30sec. The supernatants incubated for 5 min on ice and centrifuged 5,000 rpm for 5 min. The nuclear pellet was suspended in buffer B (25% Glycerol, 20 mM HEPES (pH 7.9), 420 mM NaCl, 1.2 mM MgCl2, 0.2 mM EDTA, 0.5 mM Dithiothreitol (DTT), 0.5 mM PMSF, Benzamidine, Leupeptin, Pepstatin and Aprotinin) followed by incubation on ice for 20 min. The supernatant was used to determine nuclear protein concentration. Protein concentration was measured using the Bradford assay, equal amounts of protein were separated with SDS-PAGE and then transferred to the PVDF membrane (Millipore, Billerica, MA, USA). After blocking for blocking in 5% skim milk or 3% BSA, the membranes incubated overnight at 4℃ with primary antibodies: Nrf2 (Abcam, Cambridge, MA, 1:1000), HO-1 (Stressgen, Victoria, Canada, 1:1000), GPx (Abcam, Cambridge, MA, 1:16000), catalase (cell signaling technology, Inc., Danvers, MA, 1:1000), pIκBα (cell signaling technology, Inc., Danvers, MA, 1:1000), IL-1β (Santa Cruz Biotechnology, CA, USA, 1:200), CRP (Abcam, Cambridge, MA, 1:1000), MCP-1 (Santa Cruz Biotechnology, CA, USA, 1:1000), TNF-α (Santa Cruz Biotechnology, CA, USA, 1:200), PKC-βII (Santa Cruz Biotechnology, CA, USA, 1:200), TGF-β (Santa Cruz Biotechnology, CA, USA, 1:200), β-actin (Santa Cruz Biotechnology, CA, USA, 1:1000)
Statistical analysis
All data are presented as means ± SEM. The significance of differences are analyzed by one-way ANOVA followed by Duncan's multiple range test. Values of P<0.05 were considered statistically significant. All statistical analyses were performed using SPSS software (version 20.0K for Windows) and Graph Pad Prism (version 5.0).
Go to :
RESULTS
Effects of AT and GT supplementations on body and kidney weights and food intakes in alloxan-induced diabetic mice
After 1 week of alloxan injection to induce diabetes, diabetic mice significantly had lower body weight than CON. However, AT and GT supplementations did not affect body weight change in diabetic mice throughout the experimental periods (Fig. 1A). There were no significant differences of body weights among diabetic mice regardless of FBG levels. In addition, the kidney weights of diabetic mice, as shown in Fig. 1B, were significantly higher in the diabetic mice regardless of supplementation, as compared to the CON group. The diabetic mice showed significantly higher food intake compared to the CON. However, AT and GT supplementations did not cause change of food intake in diabetic mice (data not shown).
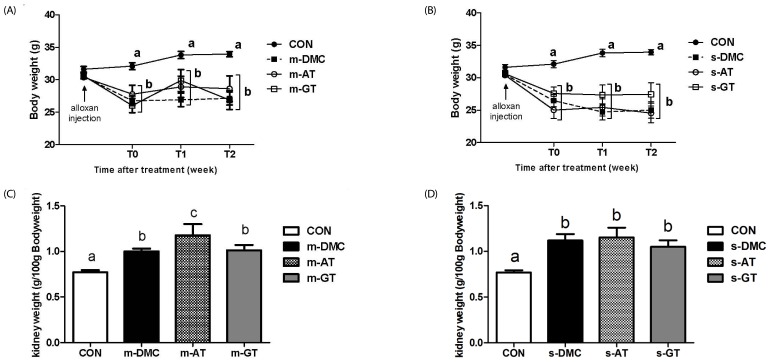 | Fig. 1Effects of AT and GT supplementations on body weight change and kidney weight in diabetic mice.Body weight change in mild DMC group (A), body weight change in severe DMC group (B), kidney weight per 100g body weight in mild DMC group (C), and kidney weight per 100g weight in severe DMC group (D). Values are means ± SEM (N = 6). Mean values with unlike letters were significantly different (P < 0.05). CON: non-diabetic mice, m-DMC: diabetic control mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-AT: alpha-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-GT: gamma-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), s-DMC: diabetic control mice (450 mg/dl < FBG), s-AT: alpha-tocopherol supplemented diabetic mice (450 mg/dl < FBG), s-GT: gamma-tocopherol supplemented diabetic mice (450 mg/dl < FBG)
|
Effects of AT and GT supplementations on fasting glucose levels in alloxan-induced diabetic mice
After 1 week of alloxan injection, fasting blood glucose levels significantly higher in the all diabetic groups than in the CON group. After 2 weeks of dietary treatment, the m-AT group significantly lower FBG levels, whereas there were no significant differences in FBG levels between the m-GT group and the m-DMC group (Fig. 2A). On the other hand, the FBG level of s-GT group was significantly lowered compared to that in the s-DMC group but there were no significant differences in FBG levels between the s-DMC and the s-AT group (Fig. 2B).
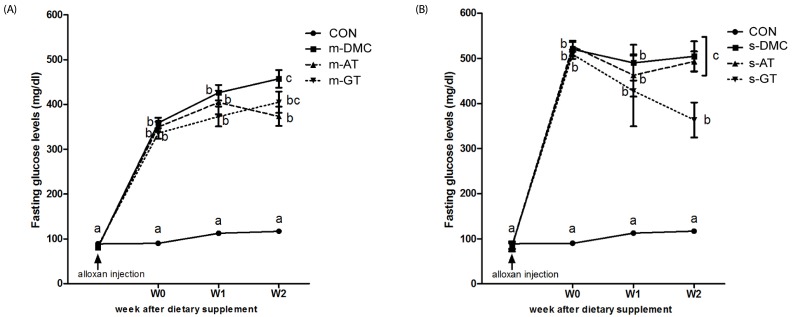 | Fig. 2Effects of AT and GT supplementations on fasting blood glucose levels in diabetic mice.Fasting blood glucose levels in mild DMC group (A), and fasting blood glucose levels in severe DMC group (B) Values are means ± SEM (N = 6). Mean values with unlike letters were significantly different (P < 0.05). CON: non-diabetic mice, m-DMC: diabetic control mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-AT: alpha-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-GT: gamma-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), s-DMC: diabetic control mice (450 mg/dl < FBG), s-AT: alpha-tocopherol supplemented diabetic mice (450 mg/dl < FBG), s-GT: gamma-tocopherol supplemented diabetic mice (450 mg/dl < FBG)
|
Effects of AT and GT supplementations on kidney function in alloxan-induced diabetic mice
Blood urea nitrogen
BUN levels were significantly higher in the m-DMC and s-DMC groups compared to the CON group (Fig. 3A). The BUN levels in m-AT and m-GT groups were significantly lower than those in the m-DMC group. In severe FBG groups, BUN levels in the s-AT and s-GT groups were significantly lower than those in the s-DMC group.
 | Fig. 3Effects of AT and GT supplementations on BUN (A), and plasma creatinine (B) levels in diabetic mice. Values are means ± SEM (N = 6).Mean values with unlike letters were significantly different (P < 0.05). CON: non-diabetic mice, m-DMC: diabetic control mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-AT: alpha-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-GT: gamma-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), s-DMC: diabetic control mice (450 mg/dl < FBG), s-AT: alpha-tocopherol supplemented diabetic mice (450 mg/dl < FBG), s-GT: gamma-tocopherol supplemented diabetic mice (450 mg/dl < FBG)
|
Plasma creatinine
The DMC groups showed higher levels of plasma creatinine compared to the CON group (Fig. 3B). The levels of plasma creatinine in AT and GT supplemented groups showed no significant differences with those in the DMC groups regardless of FBG levels.
Effects of AT and GT supplementations on MDA levels and protein levels of oxidative stress markers in diabetic kidney damage
Malondialdehyde (MDA) concentration in kidney was measured as an indicator of lipid peroxidation. MDA levels in the DMC groups were significantly higher than those in CON as shown in Fig. 4A. GT supplemented group showed lower renal MDA levels compared to the DMC groups regardless of FBG levels. The renal MDA levels in the m-AT group were lower than those in the m-DMC group, but there were no significant difference in the renal MDA levels between the s-AT and s-DMC groups.
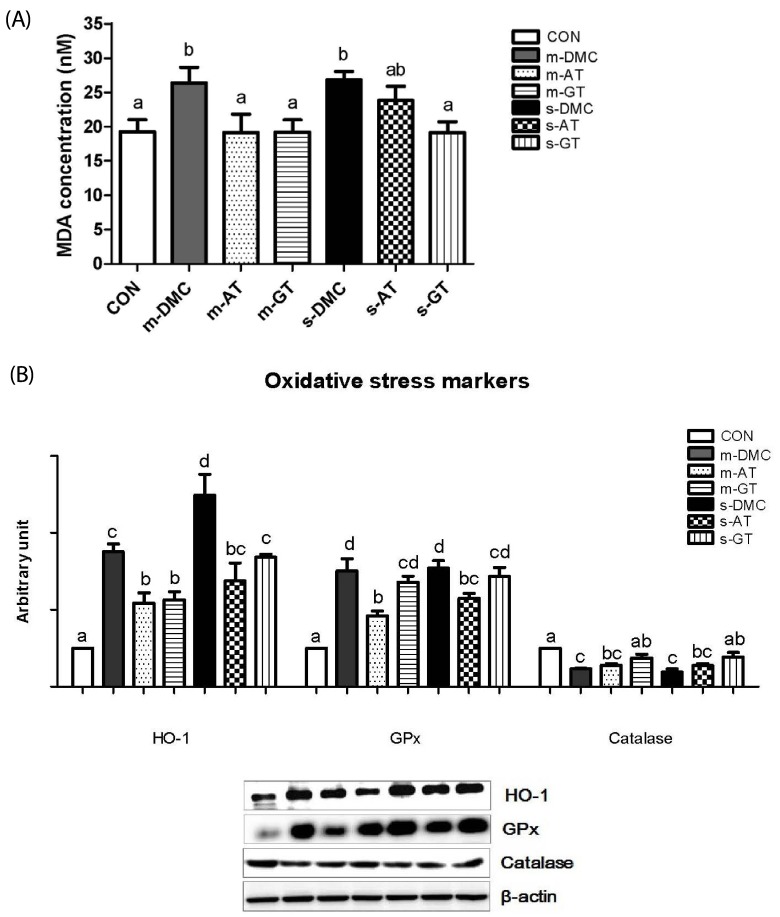 | Fig. 4Effects of AT and GT supplementations on kidney MDA concentrations (A) protein levels of HO-1, GPx, and catalase (B) in diabetic mice.Values are means ± SEM (N = 6). Mean values with unlike letters were significantly different (P < 0.05). CON: non-diabetic mice, m-DMC: diabetic control mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-AT: alpha-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-GT: gamma-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), s-DMC: diabetic control mice (450 mg/dl < FBG), s-AT: alpha-tocopherol supplemented diabetic mice (450 mg/dl < FBG), s-GT: gamma-tocopherol supplemented diabetic mice (450 mg/dl < FBG)
|
We performed western blot analysis to determine whether AT and GT supplementations decreased levels of oxidative stress proteins in DN. Protein levels of HO-1, a representative target gene in Nrf2 pathway, were significantly higher in both the m-DMC and s-DMC as compared to the CON. In addition, the protein levels of HO-1 were much higher in the m-DMC than in the s-DMC. AT and GT supplemented groups, regardless of FBG levels, showed significantly lower protein levels of HO-1 in diabetic mice (Fig. 4B,A). The GPx levels were significantly higher in the DMC groups than in the CON (Fig. 4B,B). The protein levels of GPx in both m-AT and m-GT groups were significantly lower than those in the m-DMC group. In severe FBG groups, the protein levels of GPx not in s-GT but in s-AT were significantly lower than those in s-DMC. As shown in Fig. 4B and 4C, the protein levels of catalase were higher in the CON mice compared to those in the m-DMC and s-DMC groups. The protein levels of catalase in GT supplemented mice but not in AT supplemented mice were significantly lower than those in diabetic control mice regardless of FBG levels.
Effects of AT and GT supplementations on protein levels of inflammatory markers in diabetic kidney damage
We examined whether AT and GT supplementations reduced the protein levels of inflammatory markers in hyperglycemia induced kidney damage. Protein levels of cytosolic pIκBα in kidney were measured as an indirect marker for measuring the activation of NFκB (Fig. 5A). The protein levels of pIκBα in the m-DMC and s-DMC groups were significantly increased compared to those in the CON group. The protein levels of pIκBα in AT and GT supplemented mice, regardless of FBG levels, were significantly decreased compared to those in m-DMC and s-DMC (P < 0.05). Protein levels of IL-1β, a target gene of NFκB activation, in the DMC groups were significantly higher than those in the CON group (Fig. 5B). AT and GT supplemented groups did not showed differences in the protein levels of IL-1β compared to m-DMC, while AT and GT supplemented groups showed significantly lower protein levels of IL-1β than s-DMC. As shown in Fig. 5C, the protein levels of CRP in the DMC groups were higher compared to those in the CON group. In particular, the protein levels of CRP in the s-DMC group were much higher than those in the m-DMC group. The s-AT and s-GT groups showed significantly lower protein levels of CRP compared to s-DMC. Additionally, the protein levels of MCP-1 in the m-DMC group did not significantly differ from those of the CON, m-AT and m-GT groups. On the other hand, the protein levels of MCP-1 in the s-DMC group were significantly higher than those in the CON group and the protein levels of MCP-1 in both s-AT and s-GT groups were lower than those in the s-DMC group (Fig. 5D). Next, protein levels of TNF-α were remarkably higher in all DMC groups than in the CON mice (Fig. 5E). However, there were no significant differences in the protein levels of TNF-α between diabetic control groups and AT or GT supplemented groups regardless of FBG levels.
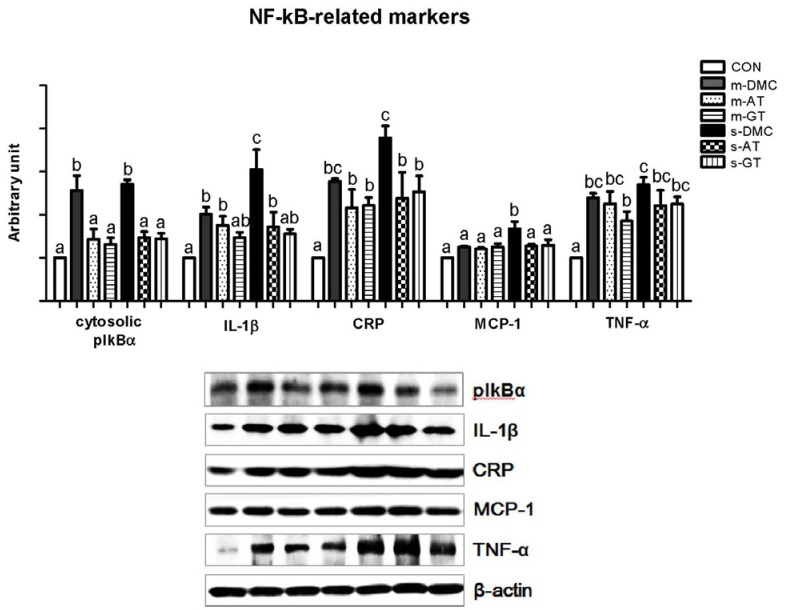 | Fig. 5Effects of AT and GT supplementations on kidney protein levels of pIkBα, IL-1β, CRP, MCP-1, and TNF-α in diabetic mice. Values are means ± SEM (N = 6).Means values with unlike letters were significantly different (P < 0.05). CON: non-diabetic mice, m-DMC: diabetic control mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-AT: alpha-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-GT: gamma-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), s-DMC: diabetic control mice (450 mg/dl < FBG), s-AT: alpha-tocopherol supplemented diabetic mice (450 mg/dl < FBG), s-GT: gamma-tocopherol supplemented diabetic mice (450 mg/dl < FBG)
|
Effects of AT and GT supplementations on protein levels of pre-fibrosis associated markers in diabetic kidneys
Protein levels of PKC-βII and TGF-βI in the DMC groups were significantly lower than those in the CON group. The protein levels of PKC-βII in the m-DMC group were not significantly different from those in m-AT and m-GT groups (Fig. 6A). However, the s-AT and s-GT groups showed significantly lowered protein levels of PKC-βII compared to the s-DMC group. Protein levels of TGF-βI, a potent fibrogenic marker, in the AT or GT supplemented groups were lower than those in the DMC groups regardless of FBG levels (Fig. 6B).
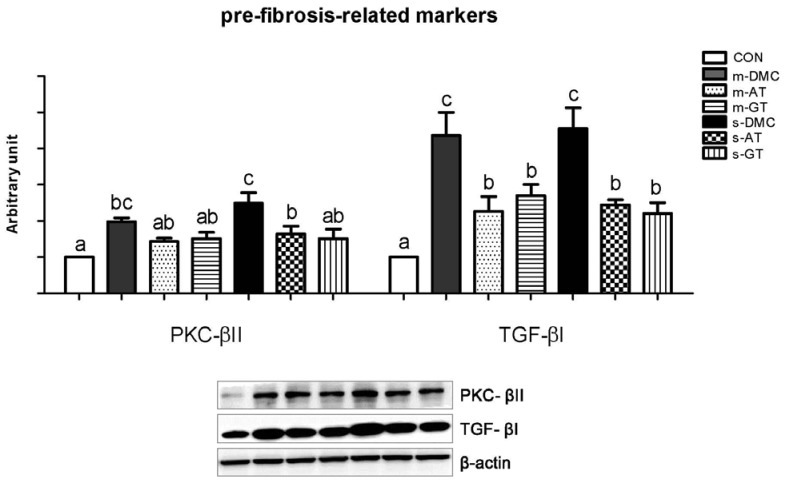 | Fig. 6Effects of AT and GT supplementations on kidney protein levels of PKC-β II, and TGF-βI in diabetic mice. Values are means ± SEM (N = 6).Mean values with unlike letters were significantly different (P < 0.05). CON: non-diabetic mice, m-DMC: diabetic control mice (250 mg/dl ≤ FBG ≤ 45 0mg/dl), m-AT: alpha-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), m-GT: gamma-tocopherol supplemented diabetic mice (250 mg/dl ≤ FBG ≤ 450 mg/dl), s-DMC: diabetic control mice (450 mg/dl < FBG), s-AT: alpha-tocopherol supplemented diabetic mice (450 mg/dl < FBG), s-GT: gamma-tocopherol supplemented diabetic mice (450 mg/dl < FBG)
|
Go to :
DISCUSSION
In the current experiment, we investigated whether AT and GT supplementations ameliorate hyperglycemia induced acute kidney inflammation through improvement of oxidative stress, inflammation, and pre-fibrosis in alloxan-induced diabetic mice with different effects according to FBG levels. Although AT and GT have beneficial influences on both oxidative stress and inflammation [20], there is no obvious fundamental mechanism in which AT and GT can improve protective roles in DN progression in accordance with FBG levels.
AT is a dominant tocopherol in the body due to alpha-tocopherol transfer protein whereas GT is more common form in Western [23] and Korean diets. Although GT is easily metabolized in the liver and remains a little amount in plasma and tissues, GT showed potent biological functions including antiinflammatory effects in rodents [25]. Moreover, previous studies have shown similar patterns in plasma AT and GT and their metabolites in rodents and humans [2829].
In this study, the FBG between 250 and 450 mg/dl was considered as mild hyperglycemia [30], and the FBG higher than 450 mg/dl was regarded as severe hyperglycemia [31]. Interestingly, this study showed that AT supplementation decreased the FBG levels in the DM with m-FBG, whereas GT supplementation showed an ability to reduce the FBG levels in the DM mice with s-FBG. In other words, short-term AT and GT supplementations could have beneficial effects on the regulation of FBG depending on basal FBG levels, respectively. In a previous study conducted by Kim et al [32], DL-α-tocopheryl acetate supplement at a 50IU/kg body weight dose for 7 weeks decreased FBG in streptozotocin (STZ)-induced rats. Although the dosage and treatment period in the previous study were higher and longer, respectively than this study, the results showed the similar effect of AT on FBG levels as the current study.
BUN and plasma creatinine, nitrogenous end products of metabolism, are often used to diagnose kidney function and damage [33]. Haidara et al. [34] reported that vitamin E supplementation at a dose of 300 or 600 mg/kg for 4 weeks resulted in reductions of both BUN and serum creatinine in STZ-induced diabetic rats. The results demonstrated a beneficial effect of vitamin E on kidney function. In this study, BUN levels in diabetic mice were increased, but AT and GT supplementations significantly decreased BUN levels in the DM mice regardless of FBG levels. BUN is usually performed together with plasma creatinine, which is a more sensitive renal damage marker. In these data, both AT and GT supplementation didn't have beneficial effects on plasma creatinine levels in the m-DM and s-DM mice. The different result between the previous study and the current study may be caused by dosage and treatment period of vitamin E supplementation. However, there was no histological changes of kidney in tocopherol treated and DMC mice (data not shown). The present study suggests that short-term supplementations of AT and GT may not be enough to normalize kidney function in alloxan induced DM mice.
Lipid peroxidation demonstrated by MDA was increased in diabetic subjects [35]. Increased oxidative stress and lipid peroxidation in a diabetic condition could induce prostaglandin production, which is the basis for the pathogenesis of DN as well [36]. A previous study showed that vitamin E reduced MDA levels in plasma, liver, brain and kidney in STZ-induced diabetic mice [37]. This study also showed that GT supplementation reduced kidney MDA levels in DMC mice regardless of FBG levels, whereas AT supplementation could decrease MDA levels only in the m-DM group, not in the s-DM group. Thus, the data suggest that GT supplementation reduces oxidative stress more effectively compared to AT in the s-DM group.
Hyperglycemia-induced oxidative stress is critical in the progress of DN. It has been known that the anti-oxidative defense system was depressed in diabetic kidney damage [5]. HO-1, a redox sensing enzyme, can be induced to protect injured tissue and tends to be increased in pathological conditions such as diabetes [38]. The present study demonstrated that AT and GT supplementations, regardless of FBG levels, reduced the expression of HO-1 levels in diabetic mice. GPx is another antioxidant enzyme which reduces highly cytotoxic products, such as lipidic or non-lipidic peroxides and other organic hydroperoxides [39]. Although there were no clear results regarding GPx expression in a diabetic condition [4041], hyperglycemia induced oxidative stress affected the balance between oxidants and antioxidants through regulation of key antioxidant enzymes such as glutathione peroxidase and catalase [42]. Our data demonstrated that protein levels of GPx were significantly increased in both m-DMC and s-DMC contrary to those in CON. AT supplementation decreased the GPx level regardless of FBG levels, whereas GT supplementation did not show any beneficial effect compared to the DMC groups. Catalase contributes to causing cells and tissues to be more resistant to H2O2 toxicity and oxidant-mediated injuries. The role of catalase is to prevent ROS generation and to attenuate kidney injury in a diabetic condition [43]. A previous study showed that loss of catalase buffering capacity causes an increase of oxidant-mediated renal fibrosis in catalase-deficient mice with progressive kidney disease [44]. Our finding demonstrated that only GT supplementation increased the levels of catalase in kidneys in the m-GT and s-GT groups compared to the DMC groups, while AT supplemented diabetic mice did not show any differences. Olivier et al. [45] provided evidence for GPx-catalase cooperativity. Briefly, upregulation of GPx may reduce toxicity from H2O2 by maintaining catalase in an active form. That is to say, enzymatic reaction time can be different and GPx may forego catalase. Collectively, AT and GT have a beneficial effect on GPx and catalase, respectively, in hyperglycemia induced renal inflammation established in this study although both GPx and catalase are target genes of nuclear factor (erythroid-derived 2)-like 2 (Nrf2). Further research is required to investigate mechanism in which AT and GT show different effect on antioxidant enzymes, GPx and catalase.
Another major target of ROS is the inflammatory-mediated transcription factor called NFκB. NFκB plays an important role in many aspects, such as DM development, progression, and treatment. Many previous studies have demonstrated experimental evidence that NFκB was activated in DN [10]. Proinflammatory cytokines regulated by NFκB have been considered as key markers in the development and progression of DN. Under normal conditions, IκBα and NFκB exist as a unified complex. However, inflammatory conditions might induce phosphorylation of IκBα, and consequently, phosphorylated IκB α is separated from the complex of IκBα and NFκB. After this dissociation, NFκB translocates into the nucleus and then activates the transcription of specific inflammatory genes [46]. Thus, the pIκBα level indirectly reflects NFκB activation in cytosol. Our data showed that the cytosolic levels of pIκBα were increased in DMC and decreased in AT and GT supplementations. The results imply that AT and GT supplementations obstructed NFκB activation demonstrated by reduction of pIκB α. An increase in NFκB activation is followed by an increase in the induction of inflammatory cytokine, such as IL-1β, CRP, MCP-1 and TNF-α in diabetic mice, which are in good agreement with other studies [12]. IL-1β, an important pro-inflammation cytokine, affects nearly every cell type. It plays an important role in the regulation of immune and inflammatory responses and the synthesis of intercellular adhesion molecule-1 (ICAM-1) and vascular cellular adhesion molecule-1 (VCAM-1), which increases the proliferation of mesangial cells and matrix synthesis [474849]. Induction of CRP is caused by a variety of pro-inflammatory cytokines such as IL-1β, IL-6 and IL-17 and has been associated with pathological conditions such as diabetes and its complications [50], and concentrations of CRP over 10 mg/L are regarded as clinically significant inflammation states [51]. Daily supplementation of tocopherol has an ability to reduce the concentration of CRP levels in human plasma [52]. In our findings, AT and GT supplementations in diabetic mice attenuated the elevated levels of IL-1β, especially severe FBG groups, thereby modulating the consequence of inflammation-mediated kidney damage. Moreover, overproduction of CRP in diabetic mice with severe FBG levels was significantly reduced by AT and GT supplementations, but not in mild FBG groups. In other words, AT and GT supplementations can be more effective under severe inflammatory conditions associated with the increase in glucose blood levels. The present study indicated that AT and GT supplementations have the similar beneficial effects on reduction of NFκB associated inflammatory responses in hyperglycemia induced renal damage. Hyperglycemic conditions stimulate cellular hypertrophy and matrix production [53]. Hyperglycemia can also activate PKC by multiple factors, such as increased polyol pathway flux and increased intracellular formation of AGEs [54]. Activated PKC contributes to pathogenic consequences by activating NFκB and various membranes associated with NADPH oxidase and by affecting expression of TGF-β, eNOS, VEGF and PAL-1, resulting in excessive ROS production [55]. PKC-β, in particular, increased levels of matrix protein synthesis mediated either directly or overexpression of TGF-βI in the glomeruli of diabetic rats [56]. These phenomena have been supported by several in vivo experiments, which showed that mice lacking PKC-β (PKCβ-/- mice) with STZ-induced diabetes were protected from glomerular hypertrophy and matrix protein accumulation [57]. As another key factor in diabetic kidney disease, TGF-βI mediated all of the pathologic changes and induced the accumulation of matrices to stimulate extracellular matrix molecules, including type I collagen, type IV collagen, fibronectin, and laminin [58]. Many studies have shown that TGF-β plays an important role in kidney hypertrophy and enhances expression of genes related to the extracellular matrix in diabetic mice [59] and diabetic patients [60]. Therefore, PKC β and TGF-βI inhibitors may be effective in prevention of DN. The current study identified the beneficial effects of AT and GT supplementations on PKC-β II in DM mice with severe FBG levels. In addition, AT and GT supplementations protected against the fibrosis process, represented by TGF-βI, regardless of FBG levels. The results suggest that both AT and GT have the similar beneficial effects on hyperglycemia induced renal fibrosis.
Taken together, both AT and GT supplementations showed similar beneficial effects on NFκB associated inflammatory response and pre-fibrosis markers as well as HO-1 in diabetic mice. On the other hands, AT and GT showed different beneficial effects on kidney weight, FBG, and oxidative stress associated makers except HO-1. In particular, GT significantly preserved kidney weight in m-DM and improved FBG levels in s-DM and lipid peroxidation and catalase in m- and s-DM, while AT significantly attenuated FBG levels in m-DM and improved glutathione peroxidase in m- and s-DM. The current findings suggest that AT and GT with similarities and differences would be considered as beneficial nutrients to modulate hyperglycemia induced acute renal inflammation in diabetic mice. Further research with careful approach is needed to confirm beneficial effects of tocopherols in diabetes with different FBG levels for clinical application.
Go to :
 XML Download
XML Download