

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Oligonol, a polyphenolic compound derived from grape seed or lychee fruit, is composed of catechin-type monomers and proanthocyanidin oligomers [1]. In vivo studies in animal models and human studies have indicated that oligonol has antioxidant and anti-inflammatory activities, indicating that it could serve as a potential candidate for the treatment and prevention of cancer, type-2 diabetes, inflammatory, and metabolic diseases [2345]. Recent studies have suggested that the antioxidant role of oligonol may be important for prevention of many diseases and we sought to determine whether the effects of oligonol were directly or indirectly associated with the activation of sirtuin and its targets.
Sirtuins are NAD+-dependent protein deacetylases that regulate metabolism and aging and these enzymes have been shown to mediate the lifespan-extending effects of caloric restriction [678]. Resveratrol, a polyphenolic compound found in wine and grapes, is a sirtuin-stimulating compound. Resveratrol has been found to increase the activation of the yeast Sir2 gene and mammalian SIRT1 gene [9] and it was recently shown that SIRT1 activation alleviated metabolic diseases in mice [10]. Because of SIRT1's role in aging, metabolism, and age-related diseases, small molecules or drugs targeting sirtuins have been implicated in the control of the biochemical, epigenetic, and cellular processes of aging [6].
Recent studies indicate that one of the major roles of the SIRT1-activated pathways is in regulating autophagy [11] and AMP-activated kinase (AMPK), the central energy sensor in the cell [12]. The responsiveness of AMPK signaling attenuates as aging increases, and this loss of sensitivity to cellular stress impairs metabolic regulation, increases oxidative stress, and reduces autophagic clearance [12]. These age-related changes activate innate immunity, triggering low-grade inflammation and metabolic disorders. Autophagy, a highly orchestrated intracellular degradation process, has emerged as a potential anti-aging mechanism and genetic inhibition of autophagy induces degenerative changes in mammalian tissues that resemble those associated with aging [13].
We wanted to characterize the effect of oligonol on sirtuin expression and downstream signaling pathways using A549 human lung epithelial cells in vitro and C. elegans in vivo. Our results demonstrated that this antioxidant induced AMPK/autophagy signaling pathways. We also showed that oligonol treatment increased T cell proliferation in old mice and increased the resistance of C. elegans to Vibrio cholerae infection. Taken together, these data suggested a potential delaying effect of oligonol on cellular senescence and aging.
MATERIALS AND METHODS
Reagents and cell culture
Human lung epithelial cells (A549) were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). A549 cells were cultured in RPMI-1640 medium (Thermo Scientific, Waltham, MA, USA) supplemented with 10% (v/v) fetal bovine serum (FBS), 100 U/mL penicillin, and 100 µg/mL streptomycin, and maintained at 37℃ with 5% CO2 in a humidified atmosphere. Oligonol was prepared as described previously [14] and dissolved in dimethyl sulfoxide (DMSO) to a final concentration of 10 mg/mL.
Reactive oxygen species (ROS) assay
Cells were treated with 10 µg/mL oligonol. After 24 h, the cells were washed twice with phosphate-buffered saline (PBS) and stained with 10-µM dicholorofluorescein diacetate (DCF-DA) (Sigma, St. Louis, MO, USA) in PBS for 30 min in the dark. Cells were then washed twice with PBS and extracted with PBS for 10 min at 37℃. Fluorescence was recorded using a spectrofluorometer (VICTOR3, Perkin-Elmer, Waltham, MA, USA) with an excitation wavelength of 490nm and an emission wavelength of 525nm.
Mitochondrial superoxide measurement
Mitochondrial superoxide levels were determined by staining with MitoSOX Red (Invitrogen, Carlsbad, CA, USA). Oligonol-treated cells were incubated with 2.5-µM MitoSOX Red at 37℃ for 15 min. The coverslips were mounted onto glass slides using mounting media containing 4,6-diamidino-2-phenylindole (Sigma) prior to examination using a confocal microscope (LSM700, Zeiss, Jena, Germany).
Reverse transcription PCR (RT-PCR) and quantitative real-time PCR (qRT-PCR)
RNA isolation and cDNA synthesis were performed as described previously [15]. Briefly, total RNA (1 µg) was isolated with Trizol reagent (Invitrogen) and reverse transcribed using the RT system (Promega, Madison, WI, USA) for 1 h at 42℃. For RT-PCR, the thermal cycling conditions consisted of 40 cycles of denaturation at 95℃ for 30 s, annealing at 60℃ for 30 s, and extension at 72℃ for 30 s. For qRT-PCR, Power SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA) was used to measure human and mouse mRNA expression. Primer sequences are shown in Table 1. The PCR included one incubation at 95℃ for 15 min, followed by 40 cycles of 30 s at 95℃ and 1 min at 60℃. β-actin or Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA was used to normalize the data. Expression was calculated using the ΔCt method, where the amount of target, normalized to an endogenous reference mRNA and relative to a calibrator, is given by 2ΔΔCt, where Ct is the cycle number of the detection threshold.
Western blotting
Cellular lysates were prepared with lysis buffer (Cell Signaling, Beverly, MA, USA) and then separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on 10-12% polyacrylamide gels and transferred to polyvinyldifluoride membranes. The membranes were incubated in blocking buffer (TBS/T; 5% non-fat milk in 0.2 M Tris, 1.36 M NaCl, 0.1% Tween 20) for 1 h at room temperature, and then washed three times for 5 min each with 15 mL of TBS/T. Membranes were incubated overnight with the primary antibodies at 4℃. Antibodies against SIRT1, p21, Microtubule-associated protein 1A/1B-light chain 3 (LC3) LC3A/B, p62, phospho-AMPK, total AMPK, and Phospho-Acetyl-CoA Carboxylase (Ser79) were purchased from Cell Signaling. An anti-GAPDH antibody was purchased from Sigma. The membranes were washed three times with TBS/T and then incubated with horseradish peroxidase-conjugated anti-rabbit IgG secondary antibody for 1 h at 25℃. After washing three times with TBS/T, the membranes were incubated with Pierce™ ECL Western Blotting Substrate (Invitrogen) and exposed to film.
Enzyme-linked immunosorbent assay (ELISA)
Supernatants were collected after 24-h exposure to oligonol and stored at -80℃ until assay. Interleukin-8 (IL-8) protein levels were determined by ELISA according to the manufacturer's instructions (Biolegend, San Diego, CA, USA). Absorbance was detected with a microplate reader at 450nm.
Lymphocyte proliferation studies using mouse splenocytes
This animal study was approved by the Korea University School of Medicine Institutional Animal Care and Use Committee (KUIACUC-20141105-3). Spleens were collected from young (3-4 months) and old (18-24 months) C57BL/6 mice under aseptic conditions in HBSS. Splenocytes were collected by passing spleen tissue through a fine steel mesh to obtain a homogeneous cell suspension; the erythrocytes were lysed with ammonium chloride (0.8%, w/v). After centrifugation (380 × g at 4℃ for 10 min), the pelleted cells were washed three times with PBS and resuspended in complete medium [RPMI 1640 supplemented with 12 mM HEPES (pH 7.1), 0.05 mM 2-mercaptoethanol, 100 U/mL penicillin, 100 µg/mL streptomycin, and 10% FBS]. The spleen cell suspension (2 × 105 cells/mL) was pipetted into 96-well plates (200 µL/well) and cultured at 37℃ for 72 h in a humid atmosphere containing 5% CO2 in the presence of recombinant protein IL-2. After 72 h, (4, 5-dimethylthiazolyl-2)-2,5- diphenyltetrazolium bromide (MTT) solution was added to each well and the plates were incubated for 4 h. Then, DMSO was added for 15 min prior to reading absorbance. The absorbance was evaluated in a microplate reader at 540nm
Mouse lung tissues
Mouse lung tissues were obtained from the Aging Tissue Bank (Pusan, Korea). Senescence marker protein (SMP)30Y/+ and SMP30Y/- mice were described previously [16]. Total RNA was purified using the RNeasy Mini Kit (Qiagen), according to the manufacturer's recommended protocol.
Influenza virus infection model
Human influenza virus A/Puerto-Rico/8/34 (H1N1) PR8 virus was kindly provided by Dr. Man Sung Park (Department of Microbiology, College of Medicine, Korea University, Korea). Virus infectivity titers of supernatants were determined y by plaque assay [17]. Human lung epithelial cells (A549) were seeded on 60-mm dishes and treated with oligonol at a concentration of 10 µg/mL for 2 h followed by PR8 infection at a multiplicity of infection of 0.1 for 1 h. Virus infection was performed in infection media (DMEM containing 2% FBS and 2 µg/mL tolylsulfonyl phenylalanyl chloromethyl ketone (TPCK)) for 1 h at 37℃. Cells were then washed with PBS and cultured in DMEM with 2% FBS for another 24 h. qRT-PCR was performed to determine mRNA expression of influenza hemagglutinin (HA).
Caenorhabditis elegans survival assays
The C. elegans fer-15(b26)II; fem-1(hc17) line was selected for liquid assay experiments because it is unable to produce progeny at 25℃ [18]. These worms were infected as described previously [19]. Worms were cultured and maintained on nematode growth medium agar containing lawns of Escherichia coli OP50. To assess the susceptibility of C. elegans to infection by V. cholerae, the bacteria were cultured in brain-heart infusion media (BHI) at 37℃. Synchronized young adult/L4 worms were infected on lawns of V. cholerae strains for 24 h at 25℃. After washing three times with M9 buffer, worms were transferred into 6-well plates (30 worms per well). Each well contained 2 mL assay medium (20% BHI:80% M9; v/v) supplemented with 1, 10, or 100 µg/mL oligonol. DMSO only was employed as control. The plates were incubated at 25℃ and examined for viability at 24-h intervals for 12 days using a Nikon SMZ645 dissecting microscope. Biological replicates of each experiment were conducted.
Statistical analysis
The results were expressed as the mean ± SD. The data were analyzed using Student's t-test to determine the significance of the difference between two groups. Statistics were performed using Prism software. A P-value < 0.05 was considered statistically significant. Differences in C. elegans survival were tested for significance by Kaplan-Meier and log-rank tests (STATA6; STATA, College Station, TX, USA). A two-tailed P-value of < 0.05 was considered statistically significant.
RESULTS
Activation of SIRT1, autophagy, and the AMPK pathway in A549 cells exposed to oligonol
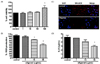
To study aging using an in vitro model, we used lung epithelial cells (A549) at a high passage number, because aging increases susceptibility to respiratory infections. To determine whether oligonol caused cytotoxicity in A549 cells, various concentrations of oligonol were added to the cells for 24 h prior to MTT assay. Even a high concentration of oligonol (100 µg/mL) did not inhibit the growth of high-passage A549 cells (Fig. 1A). ROS formation was significantly attenuated by oligonol, as shown by the DCF-DA fluorescence (Fig. 1B). We also investigated whether oligonol modulated mitochondrial superoxide formation in A549 cells. Oligonol treatment decreased the numbers of cells with mitochondrial superoxide formation, indicating that it had antioxidant activity (Fig. 1C). Furthermore, oligonol-treated cells produced less pro-inflammatory IL-8 than untreated cells (Fig. 1D).
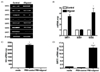
Sirtuins regulate metabolism and aging, and augment lifespan in some organisms [6]. To estimate the anti-aging effects of oligonol on sirtuin gene expression, A549 cells were treated with oligonol for 2 h. qRT-PCR was performed to investigate sirtuin (SIRT1-7) mRNA expression (Fig. 2A). SIRT 2, 3, 4, 5, and 6 mRNA expression levels were not affected by oligonol, whereas SIRT7 expression was attenuated in cells exposed to oligonol. However, qRT-PCR analysis indicated that SIRT1 mRNA expression was increased by 3-fold in cells exposed to oligonol. We also examined the expression of two mitochondrial antioxidant enzymes, superoxide dismutase (SOD) 1 and 2, and found that SOD2 mRNA levels were also up-regulated by oligonol treatment (Fig. 2B).
Given that oligonol has been shown to have anti-influenza activity [20], and SIRT1 also showed an anti-viral effect [21], we sought to determine whether oligonol modulated SIRT1 expression during influenza virus infection in A549 cells. As previously reported by another group [20], we observed that oligonol treatment of A549 cells prior to influenza virus infection led to a significant reduction in viral replication, resulting in a complete block of the expression of influenza HA in A549 cells. In the present study, influenza infection alone did not induce SIRT1 mRNA expression; however, treatment of influenza-infected cells with oligonol significantly increased SIRT1 mRNA expression by ~15-fold (Fig. 2D). These data suggested the possibility that oligonol-mediated anti-influenza activity may be associated with its up-regulation of SIRT1 expression.
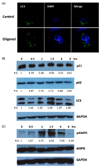
We used immunofluorescence staining with LC3, a marker protein for autophagy, to assess the effect of oligonol on autophagy in A549 cells. We observed that oligonol induced higher numbers of visible LC3 puncta in A549 cells, as compared with control treated cells (Fig. 3A). Next, we measured the protein levels of LC3-II and p62/SQSTM1, two protein markers of autophagy, using western blotting. Detection of LC3 by immunoblotting or immunofluorescence provides a reliable method to monitor autophagy, whereas the levels of p62/ SQSTM1, a ubiquitin- and LC3-binding protein, increase when autophagy is impaired [13]. Oligonol treatment significantly promoted LC3-II levels and reduced p62/SQSTM1 levels in A549 cells (Fig. 3B). In addition, expression of a cellular senescence marker (p21) was reduced by oligonol treatment. We next explored the potential mechanisms underlying oligonol-induced autophagy and examined AMPK, a ubiquitous sensor of cellular energy status, which has been shown to be connected to increased autophagic activity [22]. We found that oligonol increased the phosphorylation of AMPK at early time-points (Fig. 3C).
Oligonol affected splenocyte proliferation in old mice
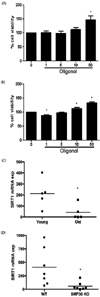
To investigate the effect of oligonol on the cellular immune response using an ex vivo system, we evaluated IL-2-induced splenocyte proliferation in mice using the MTT assay. Splenocytes were isolated from both young (3-4 months) and old (18-24 months) mice and cell viability was measured in the presence or absence of oligonol. The results indicated that whereas oligonol treatment of splenocytes isolated from young mice only induced a significant increase in splenocyte proliferation at a concentration of 50 µg/mL, oligonol levels as low as 1 µg/mL increased the proliferation of splenocytes from old mice in a concentration-dependent manner, suggesting that oligonol induced cellular proliferation in aged cells (Fig 4A).
It has been suggested that older animals produce less SIRT1 [23]. Hence, we examined the expression of SIRT1 in the lungs of young (3-4 months) and old (18-24 months) mice. Similarly, we also compared the expression of SIRT1 in the lungs of 6-month-old wild-type and SMP-30 knockout mice, which has been suggested to provide a useful model of senescence [24]. SIRT1 mRNA levels were significantly lower in the lungs of the old mice than in those of the young mice (Fig 4B). This suggested that the increased susceptibility of aged animals could be attributed to their lower SIRT1 expression and the subsequent decrease in the expression of other receptors and signaling molecules in the lungs. On the other hand, the mean expression levels of inflammatory molecules, such as IP-10, RANTES, MCP-1, and TNF-α transcripts, were significantly higher in old mice than in young mice (data not shown).
Oligonol treatment promoted survival of C. elegans infected with V. cholerae
C. elegans is an established invertebrate host model for investigating the molecular mechanisms of aging and longevity [25]. Moreover, it can be used for identifying and assessing the virulence of several human pathogens, including V. cholerae [2627]. We tested the ability of oligonol to promote longevity and induce anti-microbial activity in vivo using C. elegans liquidkilling assays. Consistent with previous results, exposure of control C. elegans to V. cholerae infection led to their death within ~8 days. Surprisingly, oligonol treatment (100 µg/mL) increased the lifespan of V. cholerae-infected worms significantly (P = 0.0001, as compared with DMSO control worms) (Fig. 5). These data suggested that oligonol increased C. elegans survival by modulating the in vivo host defense against enteric pathogens.
DISCUSSION
Oligonol has been suggested to have a beneficial effect on inflammatory diseases or cancer [2320282930]. This oligonol-mediated protection could be due to its antioxidant activity. In this study, we examined the role of oligonol as an anti-aging molecule. Our studies demonstrated that oligonol inhibited the production of ROS and mitochondrial superoxide in human lung epithelial cells. Oligonol also significantly up-regulated SIRT1 gene expression and the autophagic AMPK pathway in a time-dependent manner. Furthermore, oligonol treatment increased the proliferation of splenocytes from old mice and prolonged C. elegans lifespan by enhancing the host defense response against a microbial infection. These and other results are consistent with the suggestion that oligonol had anti-aging effects.
Mounting evidence points to the beneficial effects of plant-based polyphenolic compounds in aging-associated inflammatory disorders. We found that oligonol induced phosphorylation of proteins in the AMPK signaling pathway, as well as inducing autophagy. AMPK, a member of the AMPK protein kinase family, is a central energy sensor and metabolic switch found in all eukaryotes [12]. Recent reports have indicated that AMPK mediates the acute autophagic response by interacting with ULK1 [22], implying an essential role for AMPK in the autophagy process.
Excessive accumulation of ROS and oxidative damage has been linked to multiple diseases, including neurodegenerative diseases, diabetes, cancer, and premature aging [3132]. There are multiple sources of ROS in the cell, and mitochondria are a major site of ROS production. It was recently shown that increased mitochondrial ROS production was directly associated with increased production of pro-inflammatory cytokines and susceptibility to pathological conditions as diverse as malignancies, autoimmune diseases, and cardiovascular diseases [33]. Thus, we examined the antioxidant effect of oligonol by measuring total ROS in high-passage A549 cells, as well as mitochondrial superoxide production. Increased ROS production is closely linked to mitochondrial dysfunction and we therefore measured superoxide production from mitochondria specifically by MitoSOX staining [34]. DCF-DA revealed lower ROS levels in oligonol-treated cells and there was a specific inhibition of mitochondrial superoxide formation (Fig. 1C). Thus, targeting mitochondrial ROS can be driven by the antioxidant effects of oligonol and this could directly contribute to delaying cellular senescence.
The mammalian sirtuins (SIRT1-7) are a conserved family of NAD+-dependent deacetylases and ADP-ribosyltransferases involved in numerous fundamental cellular processes, including gene silencing, DNA repair, and metabolic regulation [8]. Recent human genetic studies also supported a role for SIRT1 in maintaining human health status with age [35]. In 2003, a screen for small molecule activators of SIRT1 identified 21 different SIRT1-activating molecules, the most potent of which was resveratrol [7]. The effects of resveratrol on aging and disease have been extensively studied to date and this compound has been found to increase lifespan in Saccharomyces cerevisiae, C. elegans, and Drosophila melanogaster in a sirtuin-dependent manner, although the lifespan extension in yeast and flies, and the Sir2-dependence in worms, have subsequently been challenged [36]. Sirtuins are attractive drug targets and the compounds and mechanisms regulating their activity have been studied intensely [6]. Given that oligonol treatment promotes survival in C. elegans during V. cholerae infection and up-regulated proliferation of splenocytes isolated from old mice, it may be worthwhile to carry out further studies of the benefits of oligonol treatment in other animal models of aging-mediated disease.
In addition to exploring the efficacy of oligonol as a potential treatment for inflammation, viral diseases, and cancer, recent studies have addressed the role of oligonol in metabolic control. For example, oligonol inhibited adipogenesis via Akt-mTOR inhibition in a cellular model [37], whereas oligonol treatment of diabetes-prone mice attenuated diabetes-induced renal damage [38]. Correlating well with previous data, our results also suggested a metabolic control effect of oligonol, which activated AMPK. Interestingly, SIRT1 and AMPK have been shown to play many similar roles, including their ability to respond to stress and nutrient status, induce mitochondrial biogenesis, regulate glucose homeostasis, and control the activity of important transcriptional regulators such as Peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC-1α), FoxO, and p300 [39] AMPK has been shown to activate SIRT1, likely indirectly via an increase in the cellular NAD+ level [40]. Given that AMPK has been shown to mediate the antioxidant effects of resveratrol through regulation of the transcription factor, FoxO1, it is highly possible that the antioxidant effect of oligonol depends on AMPK pathway activation.
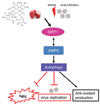
The present study generated data indicating that oligonol treatment may exert an antioxidant effect in high-passage cells, potentially via the activation of the SIRT1/AMPK/autophagy pathway (Fig. 6). Our data showing that oligonol extended the lifespan of C. elegans infected with V. cholerae suggest an important role for oligonol as an anti-aging compound in vivo. One limitation of this study was the lack of an aged in vivo model to explore the long-term effects of oligonol (as a dietary supplement) on the aging process. Further studies including oral administration of oligonol in aged animal models will provide an insight into the possible development of oligonol in preventing age-related disorders. Collectively, our data suggested that SIRT1-inducing oligonol could be introduced as a potential supplement to promote healthy aging.


 XML Download
XML Download