

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Inflammation is an important part of immune pathogenesis and is a response to tissue injury, infection, and stress. Macrophages play key roles in providing an immediate defense against foreign agents [1]. Activation of macrophages has been detected in inflammatory tissue and induced after exposure to tumor necrosis factor-α (TNF-α), interferon-γ, and a microbial lipopolysaccharide (LPS) [2]. During the progression of an inflammatory process, macrophages excessively produced various inflammatory mediators including nitric oxide (NO), prostaglandins, as well as cytokines such as interleukin (IL)-1, IL-6, and TNF-α [3]. However, the prolonged production of inflammatory mediators by macrophage can cause damage to the host and contributes to the pathology of many diseases including arthritis, asthma, cancer, diabetes, and atherosclerosis [4]. Therefore, anti-inflammatory agents are a therapeutic target for healing of inflammation-related diseases.
Activation of the mitogen-activated protein kinases (MAPKs) pathway plays an essential role in the initiation and development of inflammatory processes that are transmitted by sequential phosphorylation events [5]. The MAPKs signaling pathway, including extracellular signal-regulated kinase (ERK), c-Jun NH2-terminal kinase (JNK), and p38, have been activated by LPS via the Toll like receptor of macrophages [6]. This signaling pathway in turn activates a variety of transcription factors including nuclear factor kappa B (NF-κB) and activator protein-1 (AP-1), which coordinate the induction of a range of inflammatory proteins, such as inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), and cytokines [78]. Hence, any substances that inhibit the activation of NF-κB and AP-1 are considered potential anti-inflammatory agents.
Several biological properties of pigmented rice, such as red and black rice, including anti-oxidant and anti-cancer activities, have been reported [910]. Red and black rice are a potent source of antioxidant in functional foods. Several studies have indicated that the antioxidant capacity of red rice showed higher potency compared with white rice extract [1112]. Antioxidant compounds in red rice were classified as phenolic, flavonoid, vitamin E derivatives, γ-oryzanol, and proanthocyanidin [13]. The antioxidant properties of red rice in terms of scavenging and quenching activity for reactive oxygen species (ROS). Inflammation may be identified as the biological response to oxidative stress. Oxidative stress can lead to chronic inflammation, which in turn could mediate most chronic diseases, including cancer, atherosclerosis, and diabetes [1415]. There is significant evidence indicating that LPS induced mediators of oxidative stress lead to generation of the production of ROS, which themselves induced cytokine production [1617]. Oxidative stress can induce activation of transcription factors including NF-κB and AP-1 which up regulate the expression of inflammatory cytokines [14].
A recent study showed that some natural products could ameliorate antioxidant responses and the inflammatory response in LPS-stimulated macrophages. Some reports have demonstrated that lipophilic phytochemicals contained in rice such as γ-oryzanol, and vitamin E derivatives exert anti-inflammatory activities [1819]. On the other hand, red rice contains high amounts of hydrophilic compounds such as phenolics, flavonoids, and proanthocyanidin. Among those hydrophilic compounds catechin, ferrulic acid, and proanthocyanidin have been proven to exhibit anti-inflammatory activities [202122].
Despite these encouraging studies, reports on the cellular and molecular mechanisms responsible for the anti-inflammatory effect of red rice extract have not been published. Here, we investigated the anti-inflammatory effect of red rice extracts by measuring their ability to inhibit inflammatory mediators and cytokine production. Next, we determined the molecular mechanism underlying the anti-inflammatory impact of red rice extract in LPS-induced Raw 264.7 macrophages.
MATERIALS AND METHODS
Chemicals
ELISA kits for detection of IL-6 and TNF-α were purchased from Biolegend (San Diego, CA, USA). Fetal bovine serum (FBS) was purchased from Hyclone (Logan, UT, USA). 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and LPS were obtained from Sigma Aldrich (St Louis, MO, USA). Antibodies for detection of ERK1/2, p38, JNK, c-Jun, p65, and iNOS were purchased from Cell Signaling Technology (Danver, MA, USA). Antibody for detection of COX-2 was purchased from Calbiochem (San Diego, CA, USA). The extraction solvents and the mobile phase of HPLC, including n-butanol, ethanol, and methanol, were purchased from RCI Labscan (Samutsakorn, Thailand).
Extraction of polar and non-polar fractions from red rice
The grains of red rice (Oryza sativa L.) were harvested from Chiang Mai, Thailand. A voucher specimen number was certified by the herbarium at the Flora of Thailand, Faculty of Pharmacy, Chiang Mai University (voucher specimen no. 023148), which was kept for future reference. To obtain the whole grain of red rice, the grain was dehusked without removing bran and germ. The whole grain was ground using a mortar. The powdered specimens of red rice grains (1.0 kg) were extracted with 50% ethanol by shaking at room temperature overnight. The ethanoic solution was further extracted to determine polar fraction (RR-P) and the remaining rice grains were used to determine the nonpolar fraction (RR-NP). The ethanol was evaporated and the remaining solution was further extracted by shaking with water-saturated butanol in a separating funnel. The butanol was separated and evaporated from the water, and the water fraction was lyophilized and dried to yield the RR-P fraction (0.209% of raw material). For extraction of non-polar compounds including γ-oryzanol and vitamin E derivatives in red rice, the remaining rice grains were extracted with n-butanol by shaking at room temperature overnight. The n-butanol was collected and evaporated. This fraction was lyophilized to yield the RR-NP fraction (0.827% of raw material).
Determination of total phenolic and proanthocyanidin
Total phenolic content in red rice extract fractions was measured using Folin-ciocalteu's reagent [23]. Briefly, 300 µL of Folin-ciocalteu was added to the extract. Then, 3 ml of 5% (w/v) Na2CO3 was added to the mixture, followed by incubation for 1 h. The absorbance was measured at 600 nm and gallic acid was used as a standard.
Total proanthocyanidin (condensed tannin) in red rice extract fractions was analyzed using vanillin assay with slight modification, and using catechin as a standard [24]. Red rice extract fractions were reconstituted in sulfuric acid/methanol solution and mixed with 0.1 ml of 1% (w/v) of vanillin in methanol solution. Then 0.1 ml of sulfuric acid (H2SO4) was added, followed by incubation for 15 min in a 30℃ water bath. The absorbance of the sample was measured at 490 nm using a UV-visible spectrophotometer against a reagent blank and compared with a standard curve of catechin at various concentrations. The amount of total proanthocyanidin content in red rice extract was presented as milligram catechin equivalents per gram of extract (mg CE/g extract).
HPLC analysis for phenolic, γ-oryzanol, and vitamin E derivatives in red rice extract fractions
The phenolic compounds, γ-oryzanol and vitamin E derivatives were determined by HPLC using an Inertsil ODS-3-C18 column (phenolic compounds, γ-oryzanol) and HPLC C30 column (vitamin E derivatives) as described in our previous report [10].
Cell viability assay
Cytotoxicity of red rice extract fractions on Raw 264.7 cells was determined by MTT assay as described previously [25].
Measurement of IL-6 and TNF-α production
Raw 246.7 cells (1 × 106 cells/well) were pre-treated with or without red rice extract fractions (0-200 µg/ml) for 4 h, followed by incubation with 1 µg/ml of LPS for 24 h. Production of TNF-α and IL-6 in the condition media was determined using an ELISA kit (Biolegend Inc, San Diego, CA) according to the manufacturer's instructions [26].
Nitric oxide determination
The level of NO in the culture medium was measured using Griess reagent [27]. Raw 264.7 cells (5 × 105 cells/well) were pre-treated with red rice extract fractions (0-200 µg/ml) for 4 h, followed by incubation with or without 1 µg/ml of LPS for 24 h. The supernatant was collected and mixed with an equivalent volume of Griess reagent for 10 min. The absorbance at 550 nm was measured. A standard nitrite curve was generated in the same fashion using sodium nitrite.
Preparation of whole cell lysate and nuclear extract
Whole cell extraction was performed to determine the expression of COX-2, iNOS, and MAPKs signaling proteins in Raw 246.7 cells. The cells were pretreated with RR-P (0-150 µg/ml) for 4 h, followed by incubation with or without LPS (1 µg/ml) for 24 h or 20 min. The cells were collected and extracted with cell lysis buffer as described previously [25]. For preparation of the nuclear extract fraction, after treating the Raw 246.7 cells with RR-P (0-150 µg/ml) for 4 h, LPS (1 µg/ml) was added to the cells followed by incubation for 30 min at 37 ℃. The treated cells were then collected and washed twice with ice-cold PBS. The nuclear extraction was performed using the NucBusTer protein extraction kit (Novagen, San Diego, CA).
Western Blot analysis
Equal amounts of whole cell lysate or nuclear extract proteins were separated by 10% SDS-polyacrylamide gel electrophoresis and immunoblotted with specific antibodies (anti-COX-2, iNOS, MAPKs, c-Jun, and p65).The levels of β-actin and PARP were used as loading control for whole cell lysate and nucleus protein, respectively [28].
Statistical analysis
All experiments were performed in triplicate. Quantifications are defined as mean ± SD of three independent experiments and expressed as percentage of the control, which was considered 100%. Throughout this study statistically significant differences were calculated using one-way ANOVA (Prism version 6.0 software). A difference between the experimental groups was considered statistically significant and determined at *P < 0.05, ** P < 0.01, *** P < 0.001 or **** P < 0.0001.
RESULTS
Effect of RR-P and RR-NP on the level of inflammatory mediators
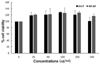
We first examined the cytotoxicity of red rice extract fractions in Raw 264.7 macrophages using an MTT assay. As shown in Fig. 1, RR-P and RR-NP at a high concentration (200 µg/ml) had no effect on cell viability of the macrophages. To examine the effect of red rice extract on the expression level of the inflammatory mediators, Raw 264.7 cells were pretreated with 200 µg/ml of RR-P or RR-NP for 4 h, followed by incubation with LPS for 24 h. As shown in Fig. 2A, only RR-P significantly reduced LPS-stimulated production of IL-6, TNF-α, and NO. In contrast, treatment with RR-NP had no effect on production of LPS-induced inflammatory mediator. In addition, LPS induced production of TNF-α, IL-6, and NO in Raw 246.7 cells was reduced by treatment with RR-P in a dose dependent manner (Fig. 2B).
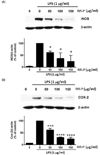
RR-P suppressed LPS-induced iNOS and COX-2 production in Raw 264.7 macrophages
To determine the effect of RR-P on the expression of iNOS and COX-2, the cells were treated with RR-P (0-150 µg/ml) for 4 h, followed by incubation with LPS for 24 h. As shown in Fig. 3A and B, the expression of iNOS and COX-2 enzymes was almost undetectable in unstimulated Raw 264.7 cells. However, after LPS stimulation, the levels of iNOS and COX-2 were significantly increased and pretreatment of macrophages with RR-P resulted in significantly reduced LPS-induced expression of iNOS and COX-2 in a dose dependent manner.
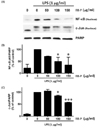
Effect of RR-P on LPS induced AP-1 and NF-κB activation
To further determine transcriptional control of RR-P in Raw 264.7 cells, we examined the effects of RR-P on the level of AP-1 (c-jun) and NF-κB (p-65) in the cell nucleus (Fig. 4). Treatment of Raw 264.7 cells with LPS alone induced the level of c-Jun and p65 in the nuclear extract and pretreatment of the cells with RR-P resulted in dramatic attenuation of the level of c-Jun and p65.
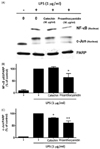
Effect of RR-P on LPS induced phosphorylation of MAPKs in Raw 264.7 macrophages
We determined the effect of RR-P on LPS-induced phosphorylation of ERK1/2, p-38, and JNK in Raw 264.7 cells. As shown in Fig. 5, phosphorylation of MAPKs was increased after LPS stimulation. However, pretreatment of the cells with RR-P resulted in significantly reduced LPS-induced phosphorylation of ERK1/2, p-38, and JNK in a dose dependent manner, while RR-P had no effect on the level of the non-phosphorylated forms of ERK1/2, p-38, and JNK. These results demonstrated that RR-P regulated LPS-induced phosphorylation of MAPKs signaling in macrophage cells.
Phytochemical content in red rice fractions
As shown in Table 1, the RR-P extract fraction had a high content of total phenolic and proanthocyanidin. In contrast, low levels of γ-oryzanol, tocotrienols, and tocopherols were detected in RR-P. On the other hand, high concentrations of γ-oryzanol, γ-tocopherol, and γ-tocotrienol were detected in the RR-NP extract fraction (Table 2). RR-NP extract fraction presented trace amounts of total phenolic. The phenolic compounds in the red rice fraction were further identified by HPLC. As shown in Table 3, the RR-P fraction contained a high content of catechin, followed by hydroxybenzoic acid and vanilic acid and low amounts of ferulic acid, protocatechuic acid, and coumaric acid.
The main phytochemicals in RR-P, catechins, and proanthocyanidin were investigated to determine the phytochemicals present in RR-P responsible for the anti-inflammatory property. Raw 264.7 cells were pretreated with 50 µg/ml of proanthocyanidin and catechin for 4 h, followed by treatment with LPS for 30 min. The nuclear extracts were used to determine the level of two critical transcription factors in the inflammatory response, NF-κB and AP-1. As shown in Fig. 6, proanthocyanidin reduced LPS-induced activation of NF-κB and AP-1. In contrast, catechins had no effect on LPS-induced activation of AP-1 and NF-κB.
DISCUSSION
In the current study, we reported for the first time that the RR-P fraction exerts anti-inflammatory activities by inhibiting the production of TNF-α, IL-6, and NO in LPS-activated macrophages. In contrast, the RR-NP fraction had no effect on LPS-induced inflammation in macrophages. In addition, RR-P suppressed the LPS-stimulated expression of iNOS and COX-2 protein in Raw 264.7 cells. These findings suggest that the RR-P fraction possesses useful anti-inflammatory activities by regulation of pro-inflammatory mediators and cytokines.
Many studies have shown that the expression of inflammatory proteins is regulated not only by NF-κB [2930] but also by other transcription factors, including AP-1 [31]. The most common component member of AP-1 is the c-Jun/c-Fos heterodimers that bind to the TPA-response elements (AP-1 site) in the promoter of various genes. In this study, we found that RR-P inhibited AP-1 and NF-κB activity by decreasing the nuclear level of c-Jun and p65, respectively. These findings are consistent with those of studies showing that the AP-1 and NF-κB response elements are present in the promoters, which can regulate COX-2, iNOS, TNF-α, and IL-6 genes.
Phosphorylation of MAPKs signaling proteins, p38, ERK1/2, and JNK, has been shown to initiate inflammatory gene expression in LPS-induced macrophages. Recent studies have indicated that the inhibition of LPS-induced MAPK signaling pathways results in the inactivation of NF-κB and AP-1 [32]. To determine whether the inhibition of NF-κB and AP-1 activation by RR-P is mediated through the MAPK pathway, the LPS-induced phosphorylation of p38, JNK, and ERK1/2 was assessed in Raw 264.7 cells. The results showed that the phosphorylation of p38, JNK, and ERK1/2 by LPS was inhibited by RR-P treatment. Thus, it is likely that the inhibition of MAPKs phosphorylation by RR-P may contribute to the RR-P mediated inhibition of NF-κB and AP-1 in LPS-induced macrophages.
Pigmented rice contains a variety of bioactive components with anti-inflammation activities [33]. However, no studies have reported on the anti-inflammatory effect of red rice extract, and the active compound associated with anti-inflammation in red rice has not been investigated. To minimize the toxicity of organic extraction solvents, alcohol base solvents including ethanol and butanol were used in this study. In the report from Jun HI et al., rice brans extracted with an aqueous mixture of ethanol showed a higher content of phenolic and flavonoid compounds than absolute ethanol [34]. In addition, 50% ethanol has been used in extraction of phenolic, flavonoids, and proanthocyanidin from different types of seeds [3536]. Therefore, we used 50% ethanol as the first solvent for extraction of the hydrophilic phytochemical compounds from red rice. On the other hand, ethyl acetate is the best solvent for extraction of phenolic and flavonoids. However, the polarity of butanol is close with ethyl acetate but with less toxicity. Thus, we used water-saturated butanol to further purified phenolic, flavonoids, and proanthocyanidin from the 50% ethanoic fraction and this extract was called RR-P. Srisaipet A et al. reported that n-butanol has been used in extraction of γ-oryzanol and vitamin E derivatives from rice bran oils comparable with hexane [37]. Here in, n-butanol was used in extraction of hydrophobic phytochemical from the remaining rice grains after 50% ethanoic extract and this n-butanol extract was called RR-NP.
Our results demonstrated that RR-P was rich in phenolic compounds and proanthocyanidins, while lower amounts or none at all were detected in RR-NP. The main phenolic component in RR-P was a catechin followed by hydroxybenzoic acid and vanilic acid. It is rational to speculate that catechins and proanthocyanidins might contribute to the anti-inflammation function of RR-P. In this study, we found that proanthocyanidins at 50 µg/ml reduced the activities of AP-1 and NF-κB, while catechins at 50 µg/ml had no effect. This result was similar to certain previous findings, which showed that proanthocyanidin derived from blackberries inhibited the inflammatory mediator in LPS-induced Raw 264.7 cells [38]. In addition, treatment of proanthocyanidins from Ephedra root reduced TNF-α and IL-1-β via inhibited NF-κB activity [39]. Our data collectively support the acknowledgement of the inhibitory effect of RR-P on anti-inflammation activity, which might be the effect of proanthocyanidin. On the other hand, the content of γ-oryzanol, tocotrienols, and tocopherols was detected in RR-NP fractions. Specifically, γ-oryzanol is the major component of the RR-NP fraction. Results of this study indicated that RR-NP did not exhibit an obvious anti-inflammatory activity at the concentration of up to 200 µg/ml. This finding was in contrast with the study by Saenjum et al., which reported that γ-oryzanol-rich extract from Thai rice, prepared and semi-purified by column chromatography, reduced NO production in LPS-induced Raw 264.7 cells [40]. However, a high concentration (IC50; 29.3 µg/ml) of the extract was used to achieve the anti-inflammation activity. Although suppression of LPS-induced inflammation by pure tocotrienols and tocopherols (8-16 µM) has been reported, the concentrations of tocotrienols and tocopherols present in RR-NP (< 1 µmol/l) were not sufficient doses to promote anti-inflammatory activity [41]. In addition, a recent study reported that γ-tocopherol or γ-tocotrienol inhibited LPS-stimulated production of IL-6 without having an effect on TNF-α, IL-10, and COX-2 in Raw 264.7 cells [42]. These observations are consistent with our findings showing that RR-NP had no significant effect on LPS-stimulated increases of TNF-α and on NO production. This could imply that the concentrations of the bioactive component in RR-NP may not reach their potential to inhibit LPS-induced inflammation.
In conclusion, this study has shown that RR-P inhibited the production of NO, IL-6, TNF-α, COX-2, and iNOS in LPS-treated Raw 264.7 cells. The inhibitory effects are mediated by the inhibition of AP-1, NF-κB activation and the MAPKs signaling pathway. The anti-inflammation activity of the RR-P fraction may be due in part to the effect of the proanthocyanidin. These results also suggest that the RR-P fraction of red rice may be a potent natural anti-inflammatory agent.





 XML Download
XML Download