

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Diabetes mellitus (DM) is a modern epidemic and the incidence of this metabolic disease is growing. It is currently recognized as a global public health concern. Approximately, 90-95% of patients with DM present with hyperglycemia resulting from defects in insulin secretion or peripheral insulin resistance [1]. There are two types of diabetes, type I diabetes (T1D) which accounts for 3-5% of DM cases, and type 2 diabetes (T2D) which accounts for more than 90% of DM cases [2]. Dysregulated carbohydrate metabolism leads to high-blood glucose levels, low levels of insulin secretion, and/or insulin insensitivity of tissues (such as liver or skeletal muscle) in T2D [3].
Administration of both streptozotocin (STZ) and nicotinamide (NA) has been shown to induce an experimental T2D model in rats, while administration of STZ alone has been used to induce a model of T1D [4]. STZ induces DM by destroying pancreatic β-cells which produce insulin for the body. Nitric oxide induced by STZ may play a role in the cytotoxic action of STZ in pancreatic β-cells [56]. Nicotinamide (pyridine-3-carboxyamide, NA), the amide form of niacin, has been shown to mediate a protective effect against cellular damage induced by different toxic agents, including STZ [78]. Blood glucose levels are substantially higher, and blood insulin levels are much lower, in the T1D model induced by STZ alone compared to the STZ-NA-induced T2D model in rats [9]. It appears that the cytotoxic action of STZ on pancreatic β-cells is partially protected by administration of NA in the STZ-NA-induced T2D model.
The liver is one of the main organs for glucose storage and it plays a crucial role in blood glucose regulation. Hepatic glucose production has been shown to be important in the development of fasting hyperglycemia in DM [10]. Therefore, abnormal activities of the enzymes that regulate hepatic glycolytic and gluconeogenic pathways may represent potential targets for controlling hepatic glucose and blood glucose homeostasis under diabetic conditions [11].
High sucrose intake is a public concern due to its potential for increasing the risk of DM and related chronic diseases [12]. Over the past few decades, intake of added sugar has increased and this may contribute to the intake of excess energy and the development of metabolic diseases, including DM [13]. It is recommended that < 10% of an individual's daily energy intake should be from added sugar [14]. It is hypothesized that a delay and/or inhibition of carbohydrate digestion can prevent postprandial hyperglycemia in DM patients. Recently, various low-calorie sugars and sugar alcohols have been developed to manage the problems associated with obesity and diabetic complications. There are natural products that also have the capability to lower blood glucose levels, and these are considered safer than therapeutic drugs. D-Xylose is an aldopentose, a precursor of hemicelluloses, and it has been reported to be an influential sucrase inhibitor [15] and to suppress postprandial glucose loading in healthy individuals [16]. The administration of 20 mg/kg body weight (b.w.) of D-xylose via gavage with 1 g/kg sucrose has also been found to effectively decrease levels of blood glucose [17]. In normal rats, intake of a solution containing sucrose (2 g/kg) and D-xylose (0.1g/kg) resulted in significantly lower blood glucose levels 30 min and 60 min later compared to control rats that received a sucrose solution alone [18]. Despite the previous evidence of an acute effect of D-xylose on blood glucose levels, it remains to be determined whether long-term intake of this aldopentose can induce a hypoglycemic effect in diabetic animal models.
Thus, the objectives of the present study were to evaluate the chronic ameliorative potential of D-xylose on hyperglycemia in STZ-NA-induced diabetic rats and to investigate D-xylosemediated mechanism(s) in vitro.
MATERIALS AND METHODS
Drugs and chemicals
D-Xylose was obtained from the TS Corporation R&D Center Food Research Unit (Incheon, Korea) and was stored in an auto-desiccator (Sanpla Dry Keepter, Sanplatec Corp, Japan). The compound, 2-[N-(7-notrobenz 2-oxa-1,3-daizol-4-yl)amino]-2-deoxy-d-glucose (2-NBDG) was purchased from Invitrogen (Carlsbad, CA, USA). Insulin, STZ, and NA were purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA).
Animals and diet
Seven-week-old male Wistar rats (210-230 g) were purchased from Central Lab Animal Inc. (Seoul, South Korea). Rats were maintained under standard conditions of 22 ± 2℃ and 50 ± 5% humidity with a 12 h/12h light/dark cycle. The rats were kept in clean cages with ad libitum access to water and diet. The rats were supplemented the adaptation diet (Purina Inc, Gyeonggi-Do, Korea) during the acclimation period for 7 d. Weight and diet intake were monitored twice a week throughout the study period. The animal study protocol for this study was approved by the Institutional Animal Care and Use Committee of Ewha Womans University (IACUC 2014-01-005).
Induction of T2D
After acclimation period, T2D was induced as described by Masiello et al. [9]. Briefly, overnight-fasted rats received a single intraperitonial injection (i.p.) of freshly prepared NA (150 mg/kg b.w.). Fifteen minutes later, 65 mg/kg b.w. of freshly prepared STZ dissolved in 0.1 M citric buffer (pH 4.5) was administered via an i.p. injection. Control rats were injected with vehicle alone. Blood glucose levels were detected using blood drops obtained from an incision made in each tail vein. Hyperglycemia was confirmed with the detection of elevated fasting blood glucose levels on day 14 post-injection using a glucometer (ACCU-CHECK Active kit, Roche Diagnostics, Mannheim, Germany). The animals with fasting blood levels higher than 200 mg/dl were used.
Experimental design
After establishing an experimental model of DM, the rats were randomized into four groups (n = 8-9/group). (i) normal control (control); (ii) diabetic control (DB); (iii) diabetic rats supplemented with a diet where 5% of the total sucrose content in the diet was replaced with D-xylose (Xylo 5); and (iv) diabetic rats supplemented with a diet where 10% of the total sucrose content in the diet was replaced with D-xylose (Xylo 10). The composition of the diets provided was listed in Table 1. The rats received these diets for two weeks. At the end of two weeks, rats were anesthetized with isoflurane (Piramal Critical Care, Bethlehem, PA, USA). Blood was collected from the inferior vena cava for analysis of plasma insulin. Both the liver and pancreas were immediately resected and washed in saline to remove excess blood.
Oral glucose tolerance test (OGTT)
OGTTs were performed after 11 days of treatment. Briefly, after 12 h of fasting, the rats were given a glucose solution (1 g/kg b.w.). Blood samples were subsequently collected from the tail vein of the rats 0, 30, 60, 90, and 120 min later. Blood glucose levels were measured using a glucometer (ACCU-CHEK Active kit, Roche Diagnostics GmbH, Germany).
Plasma insulin concentration
Blood samples were taken from the abdominal artery and were stored in tubes containing EDTA. Blood samples were centrifuged at 13,000 × g for 15 min to separate the plasma. Plasma insulin concentrations were determined using an ELISA kit (Crystal Chem, Downers Grove, IL, USA).
Western blot analysis
Liver tissues were homogenized in cold PRO-PREP protein extract solution (Intron Biotechnology, Seoul, Korea). After centrifugation, the supernatants were collected, separated using 10% SDS-polyacrylamide gel electrophoresis, and transferred to polyvinylidene diflouride (PVDF) membranes. These membranes were probed with primary antibodies raised against phosphoenolpyruvate caroxylase (PEPCK) (Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4℃ and then were washed and incubated with an appropriate secondary IgG-conjugated horseradish peroxidase antibody (Santa Cruz Biotechnology) for 1 h at room temperature. Bound antibodies were detected using enhanced chemiluminescence (ECL) detection reagents (Animal Genetics Inc. Suwon, Korea). Detection of α-tubulin (Santa Cruz Biotechnology) was used as a loading control.
Histopathology of liver and pancreas section
The left lateral lobes of liver and pancreas tissues were removed and immediately fixed with a 10% formalin-saline solution. The tissues were dehydrated with a series of alcohols, and xylene and then were embedded in paraffin. Liver and pancreas sections (4-5 µm thick) were prepared and stained with hematoxylin and eosin (H&E), then were examined microscopically with a ECLIPSE Ti-S inverted microscope (Nikon Instruments Co. Ltd, Tokyo, Japan).
Cell culture
C2C12 skeletal muscle cells were obtained from the Korean Cell Line Bank and were maintained in high-glucose DMEM medium containing 10% fetal bovine serum (FBS, Hyclone, Logan, UT, USA), INS-1 cells and pancreatic β-cells were grown in RPMI 1640 medium containing 10% FBS with 100 µg/ml streptomycin and 100 U/ml penicillin (Invitrogen) at 37℃ in a humidified 5% CO2 atmosphere.
Muscle differentiation and glucose uptake assay
Upon reaching 100% confluence, the cell culture medium for C2C12 cells was replaced with high-glucose DMEM containing 4% horse serum instead of FBS for 6 d. Fully differentiated cells were incubated in low-glucose and serum-free medium for 24 h, then the cells were treated with 20 uM 2-NBDG in the presence or absence of insulin (100 nM) or samples for 2 h. D-Xylose treatments represented a 5% or 10% substitution of the total glucose content, while 20% glucose was used as a control. After two washes with phosphate buffered saline (PBS), the cells were lysed with 0.5% TritonX-100 and fluorescence intensity was recorded at excitation and emission wavelengths of 485 nm and 535 nm, respectively, using a fluorescence micro plate reader (Molecular Devices, Sunnyvale, CA, USA).
Effect of D-xylose on insulin secretion by INS-1 pancreatic β-cells
INS-1 cells were grown in 12-well plates for 72 h. Upon reaching confluence, the cells were starved in Krebs-Ringer buffer (KRB, 125 mM NaCl, 5.9 mM KCl, 1.2 mM MgCl2, 1.28 mM CaCl2, 25 mM HEPES (pH 7.4) containing 5 mM glucose. After 2 h, the KRB was replaced with fresh KRB in the absence or presence of various concentrations of glucose (5, 10, or 20 mM) or 5% or 10% of 20 mM glucose replacement with D-xylose. After an additional 2 h, the cells were centrifuged at 1500 g for 10 min at 4℃. The supernatants were collected and insulin secretion was detected using an Ultra Sensitive Rat Insulin ELISA kit (Crystal Chem).
Statistical analysis
Results are expressed as the mean ± standard error of mean (SEM). Differences among the groups were analyzed by one-way analysis of variance (ANOVA), and differences between groups were analyzed using Newman-Keuls test using GraphPad Prism software (GraphPad Software, Inc, San Diego, CA, USA). Differences with a P-value less than 0.05 were considered statistically significant.
RESULTS
Effects of D-xylose on body weight and OGTT
The final weights for the DB group were significantly lower compared to the normal control group (Fig. 1A; P <0.001). The Xylo 10 group tended to show a higher body weight compared to the DB group, although the difference was not statistically significant. Upon administration of glucose (1 g/kg b.w.), blood glucose level was the highest at 30 min and then gradually decreased in all four groups (Fig. 1B). The area under the glucose curve (AUC) for the OGTT significantly increased for the DB control group by approximately 3-fold compared to the normal control group (P < 0.001). The Xylo 10 group exhibited a decrease in the AUC for the OGTT, although the difference was not statistically significant (Fig. 1C).
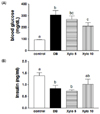
Effects of D-xylose on the fasting blood glucose and insulin secretion levels
In Fig. 2, fasting blood glucose and plasma insulin levels are shown for the four experimental groups. The DB control rats exhibited a significant increase in blood glucose levels that was approximately 3-fold that of the normal control group (P < 0.001). In contrast, rats in the Xylo 10 group exhibited a significant decrease in blood glucose levels by 31.3% compared with the DB control group (P < 0.05) (Fig. 2A). They were no significant changes in the insulin levels among all the diabetes-induced groups (Fig. 2B).
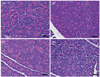
Effects of D-xylose on histopathological changes in the pancreas and liver, and changes in PEPCK expression in the liver
Histopathologic results for the pancreas tissues from all of the experimental groups are shown in Fig. 3. Pancreatic tissues from the non-diabetic controls showed a normal structure (Fig. 3A), whereas pancreatic tissues from the DB group exhibited focal necrosis, dilated acini, a decreased number and size of the pancreatic islets, and degranulation of cells (Fig. 3B). The mice treated with D-xylose showed less focal necrosis, increased number and size of pancreatic islets, and well-defined degranulation of cell compared to DB control group (Fig. 3C & 3D), although tissues from the Xylo 10 rats exhibited enhanced regeneration of the islets of Langerhans, as well as restoration of the islets (Fig. 3D).
Histopathologic results for the liver tissues from all of the experimental groups are shown in Fig. 4. Tissues from the DB group showed a disordered arrangement of hepatocytes, as well as hepatic cords and hepatic necrosis (Fig. 4A, b). In contrast, liver tissues from the mice that received D-xylose supplementation exhibited an improved arrangement of hepatocytes and regenerate hepatic cells (Fig. 4A, c, d).
Previously, expression of PEPCK was shown to play a role in gluconeogenesis [19]. Therefore, levels of PEPCK expression were analyzed in the liver tissues from each group (Fig. 4B). PEPCK levels in the DB group samples were approximately 3-fold higher compared to the control group (P < 0.001), whereas tissues from the Xylo 10 group exhibited a 28.2% reduction in PEPCK expression levels compared to the DB group (P < 0.05).
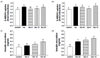
Effects of D-xylose on 2-NBDG uptake by C2C12 cells and insulin secretion by INS-1 pancreatic β-cells
To understand the mechanism(s) mediating the hypoglycemic effect of D-xylose, glucose uptake was examined in C2C12 muscle cells in the absence and presence of D-xylose. As a positive control, insulin treatment significantly increased the uptake of 2-NBDG by 15.9% (P < 0.001). Similarly, glucose treatment significantly increased 2-NBDG uptake in a dose-dependent manner (Fig. 5A; P < 0.001). Various concentrations of D-xylose were also associated with significant increase in 2-NBDG uptake in C2C12 cells. In particular, Xylo 10 increased glucose uptake higher than insulin treatment (Fig. 5B; P < 0.001). In INS-1 pancreatic β-cells, a dose-dependent increase in secretion of insulin was observed following treatment with 10 mM and 20 mM glucose (19.2% and 28.1%, respectively; P < 0.001 in each case) (Fig. 5C). Treatment with D-xylose also significantly increased insulin secretion by the INS-1 pancreatic β-cells in a dose-dependent manner. In particular, Xylo 10 increased insulin secretion by 24.3% compared to 20 mM glucose (Fig. 5D; P < 0.001).
DISCUSSION
D-Xylose has been reported to be an influential sucrase inhibitor and also suppresses postprandial glucose loading [16]. Due to its role as a sucrase inhibitor, D-xylose may be useful as a complement of sugar in sucrose-rich food products to inhibit a rise in blood glucose levels in diabetic patients or to prevent DM. However, no explicit evidence has been provided for its effectiveness in diabetic animal models or in DM patients. STZ-NA-induced diabetic rats are characterized by an acute loss in body weight despite an increase in food intake, both as a result of compromised glycemic control [20]. This decrease in body weight suggests that changes in the carbohydrate metabolism of these rats are related to the gluconeogenesis process in the liver and muscle, and structural proteins have previously been shown to play a role in body weight [2122]. Correspondingly, in the present study, D-xylose supplementation tented to be associated with increase in body weight and compared with the DB control group.
In the present study, STZ-NA-induced T2D rats were used since this animal model manifests symptoms that are comparable to those exhibited by patients with T2D [23]. STZ damages pancreatic β-cells by inducing DNA strand breakage, while NA protects pancreatic β-cells from damage and enhances their regeneration [2023]. NA has also been reported to attenuate the inhibitory effect of STZ on glucose-stimulated insulin secretion from the pancreas in rats and to improve islet cell viability [24]. Previously, diabetic rats induced by STZ-NA have been characterized by increased levels of hyperglycemia and 50% decrease in levels of pancreatic insulin [2526]. In the present study, low levels of insulin and high levels of blood glucose were detected in the STZ-NA-treated rats, thereby suggesting that the β-cells were partially injured. The moderate hyperglycemic state (≥ 200 mg/dl) that was recorded for the STZ-NA-induced rats throughout the present study confirms the diabetic status of the animals used. Furthermore, blood glucose levels increased and blood insulin levels decreased in these STZ-NA-induced diabetic rats. However, supplementation with D-xylose resulted in a significant reduction in blood glucose levels for Xylo 10 groups, while insulin secretion tended to increase in the Xylo 10 group. These data suggest that D-xylose supplementation mediates antidiabetic effects via inhibition of sucrose absorption in this T2D model. Severe postprandial hyperglycemia is commonly experienced by diabetic patients and could be prevented by reducing the rate of glucose uptake from the intestine into the circulation by inhibiting the digestion and absorption of carbohydrates.
STZ directly damages β cells of the pancreatic islets of Langerhans and, results in degranulation and loss of insulin secretion [27]. For diabetes, clarification of the regeneration potential in experimentally-induced diabetic animals would be important for identifying alternative therapies. In the present study, the histopathological analysis of pancreas tissues from the control group detected a normal appearance for the Langerhans islet. However, in pancreas tissues from the DB group, destruction of the pancreatic β cells was accompanied by loss of the nuclear envelope and mitochondrial cristae, and dilation of the rough endoplasmic reticulum. For the rats that received D-xylose supplementation, the islet tissues were protected from damage and regeneration of the pancreatic islet was observed with hyperplasia. The hypoglycemic effect associated with the latter may be due to regeneration of the islet of Langerhans, and this would be consistent with the efficacy of D-xylose in the management of diabetes. However, insulin secretion in the Xylo 10 group was slightly increased compared to the DB control group, not significantly. This result suggests that the rats treated D-xylose did not fully recover from glucose tolerance and glycemia, and the β-cells were still affected by the functional damage induced by STZ. Because blood glucose level and the histopathological analysis of pancreas tissues was improved, but insulin secretion was not significantly increased in the rats treated D-xylose.
Endogenous glucose production primarily occurs in the liver. In DM patients, lower levels of enzyme activity have been associated with the glycolytic and pentose phosphate pathways, while higher levels of enzyme activity have been associated with the gluconeogenic and glycogenolic pathways [11]. T2D is accompanied by a higher risk of liver dysfunction, and hyperglycemia can occur as a result of impaired insulin action on hepatocytes, thereby leading to a dramatic increase in hepatic glucose production via glycogenolysis and gluconeogenesis [28]. In the present study, a controlled degree of histopathological changes were observed, including hydropic degeneration and cell necrosis in the liver tissues from the DB group. In contrast, mild degeneration of these histopathological changes was observed in the Xylo 5 and Xylo 10 groups. PEPCK is a rate-limiting enzyme for gluconeogenesis and acts by catalyzing the conversion of oxaloacetate to phosphoenolpyrubate, the process whereby glucose is synthesized and is inhibited by insulin. Insulin resistance often results in an increased hepatic glucose out-put due to an increase in gluconeogenic gene expression. [1929]. In most animal models of diabetes, expression levels of PEPCK are elevated [3031]. Activation of AMP-activated protein kinase (AMPK) regulates glucose metabolism by triggering intracellular metabolic changes such as an increase in glucose uptake by muscle and other tissues and a decrease in glucose production in the liver via suppression of PEPCK expression (thereby inhibiting gluconeogenesis) [3233]. The protein, p38 MAPK, mediates the pH-responsive induction of PEPCK mRNA in glucogenic cells, while inhibition of p38 suppresses gluconeogenesis by blocking PEPCK gene [34]. In the present study, protein levels of PEPCK did not markedly increase in the Xylo 10 group, suggesting that D-xylose may reduce blood glucose levels by activating AMPK and suppressing p38 MAPK-mediated PEPCK regulation of gluconeogenesis. Thus, further studies to investigate the roles of AMPK and p38 MAPK are warranted.
In many studies, the fluorescent deoxyglucose analog, 2-NBDG, has been transported into skeletal muscle, thereby providing a marker for glucose uptake [353637]. In the present study, the uptake of 2-NBDG by C2C12 myotube cells was increased with D-xylose treatment and the level was higher than that of following treatment with 100 nM insulin. Insulin secretion from INS-1 pancreatic β-cells was also found to be dose-dependent and it was increased following treatment with Xylo 5 and Xylo 10 supplementation compared with exposure to 20 mM glucose. Taken together, these results indicate that the anti-diabetic effects that are mediated by D-xylose may involve an increased uptake of glucose by muscle tissue and increased insulin secretion from pancreatic β-cells. In the present study, C2C12 cells were not exposed to D-xylose alone in glucose-free medium because this condition would mimic the effects of glucose starvation. Insulin secretion was slightly increased in the Xylo 10 group but not significantly in vivo, although insulin secretion in treatment with Xylo 10 supplementation were significantly higher than compared with exposure to 20 mM glucose alone in vitro. It is possible this discrepancy is due to inter-animal variations and the effects of cell damage. For example, INS-1 is an intact normal pancreatic β-cell cell line. In contrasts, the pancreactic β-cells that were assayed in vivo had been damaged by STZ. This damage may have caused the β-cell to secret lower levels of insulin in vivo, and it may have been difficult for these cells to fully recover during the D-xylose supplementation period.
Previously, several studies have reported the induction of a hypoglycemic effect by D-xylose in both animals and humans [1618]. In rats that received a mixed solution of 5% xylose and 2 g/kg sucrose, lower blood glucose levels were detected 30 min and 60 min later compared with rats that received a sucrose solution [18]. In a study of healthy individuals, ingestion of a solution with 10% or 15% of its sucrose content replaced by D-xylose resulted in a significant decrease in blood glucose levels 15, 30, and 45 min later [16]. In the present study, 5% or 10% of the total sucrose content of the diet administered was supplemented with D-xylose, and these doses are comparable to those used in human trials. Dietary protein and fat may also affect blood glucose levels [3839]. Accordingly, the results from the present study suggest that D-xylose supplementation as part of a diet may not exert the same effects as D-xylose administered as a sucrose solution. It is also possible that the metabolic disturbances, impaired insulin secretion, and insulin resistance of the STZ-NA-induced rat model of T2D differ from healthy individuals [3].
To the best of our knowledge, the results of the present study demonstrate, for the first time, that D-xylose exerts anti-diabetic effects in STZ-NA-induced T2D rats by regulating blood glucose levels. This regulation may involve the regeneration of pancreas tissue and control of gluconeogenesis through suppression of PEPCK and increased glucose uptake by muscle tissue. Thus, the present results demonstrate the potential therapeutic benefit of D-xylose in regulating blood glucose levels in diabetic patients, and additional long-term studies are needed to confirm this beneficial effect.
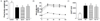


 XML Download
XML Download