

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
It has been known that the skin graft across minor histocompatibility of male antigen (HY) mismatch is normally rejected (1,2). However, in our previous report, islet transplantation across HY barrier without any immunological manipulation induced HY antigen specific immunological tolerance which prevented the rejection of subsequent skin graft in C57BL/6 mouse model (3). In that tolerance, increased Foxp3+ regulatory T cells (Treg) mediated the tolerance induction. To explore the factor that made Treg increase and tolerance induction caused by islet transplantation, we started subtraction study using knock-out (KO) mouse. Our first choice was interferon-γ (IFN-γ) KO mouse.
IFN-γ was regarded traditionally as a pro-inflammatory cytokine. However its protective role in Th-1 cell-associated autoimmune diseases has recently been reported (4). For instance, ablation of IFN-γ resulted in aggravation of murine autoimmune disease, collagen-induced arthritis (CIA), indicating that endogenous IFN-γ inhibits important inflammatory pathway (5,6). The protective effects of endogenous IFN-γ also have been reported in other autoimmune models such as experimental autoimmune encephalomyelitis (EAE), experimental autoimmune uveitis (EAU), autoimmune nephritis, and myocarditis (7-13).
In Treg development, activation of signal transducer and activator of transcription-1 (STAT-1) seems to have a role (14). This suggests that cytokine, such as IFN-γ, that activate STAT-1 have a role in Treg development (4). Wang et al. (15) have reported that in vitro treatment of CD4+CD25- T cells with IFN-γ leads to their conversion to Foxp3+ Treg cells. And, copolymer-I induced in vitro Treg conversion was dependent on IFN-γ (16). In addition, Feng et al.(17) reported that ex vivo IFN-γ conditioning of CD4+ T cells with immature dendritic cells resulted in the Treg conversion, and these induced Treg cells could prevent allograft rejection. Another study in mice showed that IFN-γ expression in Treg cells is transiently up-regulated after alloantigen exposure in vivo and is important for their regulatory function (18). In another study, in vitro activated Treg cells have shown to produce IFN-γ and to also stimulate dendritic cells to produce indoleamine 2,3-dioxygenase (IDO) (19). This enzyme converts the essential amino acid tryptophan into N-formylkynurenine (20). By depleting tryptophan, IDO exerts the capacity to inhibit T-cell response. Recent evidence suggests an important role of IDO in the induction of tolerance (21).
Considering roles of IFN-γ in self-tolerance and Treg development, we speculated that deficiency of IFN-γ might result in the failure of tolerance induction in our male islet transplantation model. We examined immunogenicity of male skin in IFN-γ KO mice, and transplanted male islets to IFN-γ KO mice. By these experiments, we confirmed the role of IFN-γ in tolerance induction by male islet transplantation.
MATERIALS AND METHODS
Mice
C57BL/6-Ifngtm1Ts(IFN-γ knock-out) mice were purchased from The Jackson Laboratory (Bar Harbor, ME, USA). All IFN-γ KO mice used in this study were genotyped for IFN-γ. PCR was done with primers and cycling conditions recommended by The Jackson Laboratory. The normal endogenous gene was detected with forward primer IMR126 (5'-AGA AG T AAG TGG AAG GGC CCA GAA G-3') and reverse primer IMR127 (5'-AGG GAA ACT GGG AGA GGA GAA ATA T-3'). The mutant (knock-out) IFN-γ was detected with forward primer IMR128 (5'-TCA GCG CAG GGG CGC CCG GTT CTT T-3') and reverse primer IMR129 (5'-ATC GAC AAG ACC GGC TTC CAT CCG A-3'). Endogenous IFN-γ resulted in a 220-bp PCR product and mutant IFN-γ resulted in a 375-bp PCR product. Mice were bred and housed in a specific pathogen-free facility. Animal studies were conducted under protocols approved by Seoul National University Institutional Animal Care and Use Committee.
Induction of diabetes mellitus and blood glucose monitoring
To induce diabetes mellitus, mice were injected intra-peritoneally with 125 mg/kg streptozotocin (Sigma, St. Louis, MO, USA) in two consecutive days. After the injections, non-fasting blood glucose levels were monitored using a glucometer, OneTouch Ultra (LifeScan Inc., Milpitas, CA, USA) from blood obtained by tail snipping. Mice with two consecutive non-fasting blood glucose levels higher than 250 mg/dl were considered as hyperglycemic mice. Hyperglycemic mice were selected for the islet transplantation.
Isolation and transplantation of pancreatic islets
Sacrificed donor mice's pancreases were injected with Hanks balanced salt solution containing 0.5 mg/ml of Collagenage P (Roche, Mannheim, Germany) via pancreatic duct. Inflated pancreases were excised and incubated in a 37℃ water bath for 20 minutes. Digested pancreases were filtered through a sieve and washed. Then, islets were purified from digests using Euro-Ficoll gradients. Obtained islets were re-suspended with RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 50µg/ml gentamycin, 100µM non-essential amino acids (all purchased from Invitrogen). After overnight culture, islets were used for the transplantation. Recipient mice were anesthetized with Isoflurane. Then, the left kidney was exposed and 250~300 islet equivalent number (IEQ) islets were delivered beneath the renal capsule.
Skin grafting
Full-thickness tail skins obtained from donors were transplanted to the anesthetized recipient mice's graft beds on the left flank and covered with Vaseline gauze and Band-Aid (Johnson&Johnson, New Brunswick, NJ, USA). Bandages were removed after 7 days, and grafts were observed every 2 to 3 days for 3 week, and weekly thereafter. The graft was scored as rejected when less than 10% of viable tissue remained and visible inflammation ended.
Nephrectomy & histological analysis
Islet recipient's left kidney was removed under the anesthesia to confirm the graft function and to analyze graft histology. Removed kidney was frozen embedded in optimal cutting temperature (OCT) compound (Sakura Finetek, Torrance, CA, USA). 5µm frozen section was done using Leica CM1850 cryocut microtome (Leica, Wetzlar, Germany). Acetone-fixated sections were stained with the hematoxylin and eosin. For insulin staining, sections were treated with guinea pig anti-insulin antibody (Dako Cytomation, Glostrup, Denmark) for 1 hr at room temperature. After wash with PBS, sections were treated with Alexa Fluor 555-conjuated anti-guinea pig IgG antibody (Invitrogen) for 1 hr at room temperature. Fluorescence microscopy was performed using Carl Zeiss Axio Imager A1 (Carl Zeiss, Jena, Germany). Images were analyzed using AxioVision software (Carl Zeiss).
RESULTS
Rejection of male skin in interferon-γ knock-out model
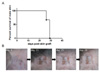
In our previous study, wild type C57BL/6 female mice rejected male skin because of the minor antigen (HY) disparity (3). To test whether the absence of interferon-γ (IFN-γ) has an influence on the immunogenicity of male skin, we obtained skin from male IFN-γ knock-out (KO) mouse and transplanted it to female IFN-γ KO mice. Female recipients rejected male skin within 29 days (Fig. 1A). On the other hand, the recipients successfully accepted co-transplanted female skin. We could observe new hair growth of grafted female skin (Fig. 1B). Therefore, non-acceptance of male skin was not technical failure but rejection.
Acceptance of male islet and exclusion of endogenous islet regeneration
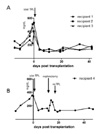
Wild type C57BL/6 female mice did not reject male islet and accepted subsequently grafted male skin (3). To test whether the absence of IFN-γ has an influence on the tolerance induction by male islet transplantation, we performed same male islet transplantation in the IFN-γ KO model. Islets were obtained from male IFN-γ KO mice, and transplanted to the left kidney subcapsular area of streptozotocin-induced diabetic female IFN-γ KO mice. Male islet transplantation normalized high blood glucose level of recipients, and the recipients did not reject male islet for >42 days (Fig. 2A).
To rule out the possibility of recipient's endogenous islet regeneration, we removed transplanted male islet in the left kidney by nephrectomy. Three days after the removal of transplanted islet, recipient returned to the hyperglycemic state (Fig. 2B). Therefore, the maintenance of normal blood glucose level was not because of the endogenous islet regeneration, but was dependent on the presence of transplanted male islet. To re-normalize the blood glucose level, and to check the recipient's response to 2nd antigen challenge, we again transplanted male IFN-γ KO islets to the recipient. 2nd islet transplantation promptly re-normalized the hyperglycemia, and the recipient maintained normal blood glucose level for >32 days (Fig. 2B). Because recipient did not reject 2nd male islet, we could conclude that non-rejection of 1st male islet was not just adaptation, and 1st male islet transplantation did not provoked immunological memory to male antigen.
Presence of insulin in grafted islet
To confirm whether the grafted islets were functioning, we analyzed the insulin secretion by grafted islet using the insulin staining of the graft site. Removed kidney containing grafted islet of recipient in Fig. 2B was frozen-sectioned and stained with hematoxylin & eosin to identify the position of graft site (Fig. 3A). Insulin staining was also conducted on the serial section. We could detect plenty of insulin-specific spots in islet region (Fig. 3B). Indicating that grafted islets were actively secreting insulin. As a control staining, only secondary Alexa Fluor 555-conjugated anti-guinea pig IgG antibody staining without primary anti-insulin antibody staining, did not produced any positive signal (Fig. 3C). Therefore, we could exclude the possibility of non-specific secondary antibody binding in Fig. 3B.
DISCUSSION
Role of IFN-γ in self tolerance was occasionally reported in autoimmune disease models. However its role in transplantation tolerance is not yet reported. As both the autoimmune disease studied and the graft rejection are mediated by Th1 dependent immune responses. We speculated that IFN-γ could have a role in transplantation tolerance, too. Our male islet transplantation model is unique in that transplantation of a graft without any immunological intervention induced antigen specific T cell tolerance. Therefore, we tried to check the role of IFN-γ in transplantation tolerance using our male islet transplantation model.
IFN-γ has been known to play a very important role in graft rejection (22), we first tested whether the absence of IFN-γ in the IFN-γ KO mouse could affect the consequences of skin and islet transplantation across HY barrier. Despite the absence of IFN-γ, female IFN-γ KO mice rejected male IFN-γ KO skin and accepted male IFN-γ KO islet as was observed in the previous results in wild type C57BL/6 model.
Although results of male skin and islet transplantation were the same as those in WT mice, the fate of male skin graft in the recipient harboring prior male islet graft should be checked to confirm the induction of tolerance. However, 2nd islet transplantation result indirectly suggests the possibility of tolerance induction. If the 1st islet transplantation provoked normal immune response to male antigen, there should be accelerated immune response by the induction of immunological memory. However, the recipient accepted 2nd challenge of male islet graft indicating the recipient got tolerized to male antigen of HY.
Our investigation on the role of IFN-γ in this study could not be generalized to transplantation tolerance. To induce transplantation tolerance, many investigators used co-stimulatory blockade or co-receptor blockade regimens (23-27). However our male islet transplantation induced tolerance without any immunological manipulation. Although our method also induced transplantation tolerance, this unique trait distinguished our method from other conventional methods. So, it is hard to say that our model is a representative of general tolerance induction model. Therefore, further investigations are required to confirm the role of IFN-γ in transplantation tolerance.
As an effort to delineate the underlying mechanisms of immune tolerance induced by the male islet transplantation, we checked the possible involvement of IFN-γ. The achievement of immunological tolerance across HY barrier in IFN-γ KO mouse suggests that IFN-γ would not be the key factor for the islet transplantation mediated tolerance induction. Further study should be needed to see if other factors known to be required for tolerance induction, such as TGF-β (28), PD-L1 (29) would be involved in the process of tolerance induction.

 XML Download
XML Download