

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Bone is actually a dynamic organ.1 Normal bone mass and strength are maintained by constant "bone remodeling" processes.2 The old bone is degraded by osteoclasts and replaced by osteoblasts. Therefore, the balance between osteoblastic bone formation and osteoclastic bone resorption is important for normal bone homeostasis. Excessive bone resorption by osteoclasts often causes osteopenic diseases including osteoporosis and rheumatoid arthritis.1
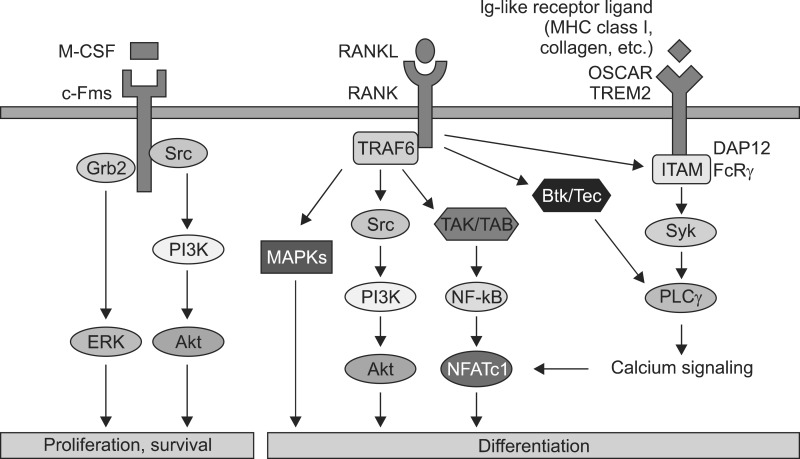
Osteoclasts differentiate from bone marrow monocyte/ macrophage lineage cells under the control of two essential cytokines.3 The binding of the macrophage colony-stimulating factor (M-CSF) to c-Fms provides signals required for proliferation and survival of osteoclast precursor cells, whereas the binding of receptor activator of nuclear factor-κB ligand (RANKL) to RANK stimulates signals required for osteoclast differentiation and the resorptive function as well as the survival of mature osteoclasts.45678 Recently, several genetic studies have shown that additional signals that can support RANK signaling are necessary for the full differentiation of osteoclasts (Fig. 1).910
Here, we discuss the important signaling pathways in osteoclast differentiation.
M-CSF-C-FMS SIGNALING
The pivotal roles of M-CSF (CSF-1) in osteoclast differentiation were revealed by studies using mice (op/op) and rats (tl/tl). The op/op mice and tl/tl rats, which have a point mutation in the csf1 gene and express non-functional M-CSF, develop severe osteopetrotic bone phenotypes due to a complete absence of osteoclasts.1112 M-CSF plays an important role in supporting the proliferation and survival of osteoclast precursor cells. The binding of M-CSF to its cognate receptor c-Fms results in auto-and trans-phosphorylation of specific tyrosine residues in the cytoplasmic tail of c-Fms. It has been well demonstrated that four (underlined) of the eight tyrosine residues (Y544, Y559, Y697, Y706, Y721, Y807, Y921, and Y974) within the cytoplasmic tail of c-Fms functionally regulate the proliferation and survival of osteoclast precursor cells.131415161718 Particularly, phosphorylation of Y559 is required for the full activation of c-Fms. Phosphorylated Y559 interacts with c-Src.19 The resulting phosphor-Y559/c-Src complex recruits the phosphatidylinositol 3-kinase (PI3K) and c-Cbl complex, which in turn activates the Akt pathway and causes c-Fms ubiquitination, respectively.2021 The c-Cbldependent c-Fms ubiquitination augments its tyrosine phosphorylation and activation via a conformational change in the kinase domain. Phosphorylated Y721 also activates the Akt pathway through direct interaction with PI3K.2022 On the other hand, phosphorylated Y697 and Y974 interact with Grb2 to mediate activation of ERK.23 Therefore, M-CSF-induced activation of c-Fms results in enhanced osteoclast precursor proliferation and survival through the ERK and PI3K/Akt pathways. Although binding partners and the precise signaling mechanism have not been fully identified, phosphorylation of Y544 and Y807 are also required for c-Fms activation and osteoclast differentiation.1824 The pivotal roles of M-CSF in osteoclast differentiation are also supported by analysis of the csf1r (gene coding c-Fms)-lacking mice, which exhibit an osteopetrotic bone phenotype.25
RANKL-RANK SIGNALING
RANKL (OPGL, ODF, and TRANCE) and its cognate receptor RANK are also key osteoclastogenic factors.26 Osteopetrotic bone phenotypes without osteoclasts of both RANKL-and RANK-deficient mice have well revealed that both factors are implicated in regulating osteoclast formation and function.2728
Binding of RANKL to RANK leads to recruitment of TNF receptor-associated factor (TRAF) adaptor proteins including TRAFs 1, 2, 3, 5, and 6 to the conserved TRAF domain within the cytoplasmic domain of RANK.2930 Among the TRAF members, TRAF6 is the most critical for osteoclast formation and function since TRAF6-lacking mice develop severe osteopetrosis owing to impaired osteoclast differentiation or bone resorption.3132 TRAF6 transmits the RANKL/RANK signal to downstream targets such as nuclear factor kappa B (NF-κB), c-Jun N-terminal kinase (JNK), extracellular signal-regulated kinase (ERK), p38, Akt, and NFATc1.83132333435 However, IL-1 administration failed to induce osteoclast differentiation in RANK knockout mice, although TRAF6 is a downstream molecule for both RANK and IL-1R (IL-1 receptor), which suggests that RANK may also activate a TRAF6-independent signaling pathway to induce osteoclast differentiation.36
Recruitment of TRAF6 to RANK forms a signaling complex containing TGF-β-activated kinase (TAK) 1 and TAK-1-binding protein (TAB) 2 to activate all three mitogen-activated protein kinase (MAPK) pathways including ERK, JNK, and p38.37 The importance of TRAF6-dependent MAPK activation was confirmed by several studies. RANKL could not activate JNK and p38 in TRAF6-deficient spleen cells.33 A specific inhibitor of p38α and β (SB203580) suppressed RANKL-mediated osteoclast differentiation in RAW 264.7 cells, and osteoclast precursor cells derived from jnk1-lacking mice but not from jnk2-lacking mice exhibited reduced ability to differentiate to osteoclasts.3839 Based on a recent study, mice with genetic deletion of erk1 exhibited reduced osteoclast formation in vivo, suggesting that ERK1 plays an important role in osteoclast differentiation.40 RANKL activates ERK, JNK, and p38 through activation of MEK1/2, MKK7, and MKK6 to induce activation of their downstream targets such as c-Fos, AP-1 transcription factors, and MITF in osteoclast precursors, respectively.414243444546
RANKL also activates the PI3K/Akt pathway through TRAF6.8 In osteoclasts, activation of PI3K/Akt may be dependent on Src kinase activity, since a Src family kinase inhibitor or genetic deletion of c-Src inhibits RANKL-mediated Akt activation.8 Activated PI3K generates phosphatidylinositol-(3,4,5)-phosphate (PIP3) at the plasma membrane to recruit Akt.47 Two negative regulators of PIP3 production, phosphatase and tensin homolog (PTEN) and SH2-containing inositol phosphatase 1 (SHIP1), negatively regulate osteoclast differentiation. The PI3K inhibitor LY294002 also has an inhibitory effect on osteoclast formation.484950 Taken together, these studies show that the PI3K/Akt pathway is involved in osteoclast differentiation.
NF-κB is one of the important transcription factors for osteoclast differentiation that are activated by RANKL. NF-κB is activated through two pathways: classical and alternative. It has been shown that TRAF6 activates the classical NF-κB pathway, whereas TRAF2 and TRAF5 activate both classical and alternative NF-κB pathways.3751 Five protein groups—Rel, RelA, RelB, NF-κB1 (p50), and NF-κB2 (p52)—constitute NF-κB proteins, which are dimeric transcription factors that bind to κB sites. Among these protein groups, p50 and p52 lack a transcription activation domain and therefore require dimerization with other NF-κB proteins possessing a transcription activation domain, such as Rel, RelA, and RelB.5253 The importance of NF-κB proteins is recognized by p50/p52 double-knockout mice, in which osteoclastogenesis was prevented, although single knockout of p50 or p52 did not show a distinct phenotype.5455 In its inactive state, NF-κB exists as a complex with inhibitory κB protein (IκB) in the cytoplasm. Upon RANKL stimulation, IκB kinase (IKK) is activated; it phosphorylates IκB and mediates ubiquitin-dependent proteasomal degradation of IκB, and ultimately results in release of NF-κB from the NF-κB/IκB complex. NF-κB released from the NF-κB/IκB complex is now free to translocate from the cytoplasm to the nucleus and binds to its target genes.5253 IKK consists of a catalytic component (IKKα, IKKβ) and a regulatory component (IKKγ). Although both IKKα and IKKβ are involved in catalytic activity, IKKβ seems to function more critically as a catalytic component. Interruption of IKKα resulted in the disruption of osteoclast differentiation in vitro only, whereas interruption of IKKβ disrupted osteoclast differentiation both in vitro and in vivo, suggesting that IKKβ is critical for RANKL-mediated IκB degradation.56 An alternative pathway of NF-κB activation exists, in which RelB:p52 dimers are involved. In the alternative NF-κB activation pathway, NF-κB-inducing kinase (NIK) and IKKα are required for the formation of the RelB:p52 complex from p100.525356 Therefore, NIK plays an important role in the alternative pathway. However, osteopetrosis was not observed in NIK-deficient mice, suggesting that NIK-induced alternative NF-κB activation is not critical for NF-κB activation.57
DISCUSSION
RANKL signaling strongly induces NFATc1, which is a master transcription factor for the terminal differentiation of osteoclasts. RANKL activates NF-κB and c-Fos to stimulate induction of NFATc1 in the early phase of osteoclastogenesis.58 RANKL-mediated NFATc1 induction is impaired in both p50/p52-deficient and c-fos-deficient cells.5960 The activated c-Fos may cooperate with NFATc1 itself for vigorous induction of NFATc1 during terminal osteoclastogenesis.58 Thus, RANKL enhances NFATc1 transcription through the activation of two essential transcription factors for osteoclast differentiation, NF-κB and c-Fos, which in turn induce a self-sustaining positive autoregulatory system to maintain sufficient NFATc1 expression. NFATc1 regulates several osteoclast-specific genes including tartrate-resistant acid phosphatase (TRAP), osteoclast-associated receptor (OSCAR), and cathepsin K in cooperation with other transcription factors.3558616263 An essential role of NFATc1 in osteoclasts has been well established both in vitro and in vivo. In vitro, NFATc1-deficient embryonic stem cells failed to differentiate into osteoclasts, and ectopic expression of NFATc1 in osteoclast precursor cells induced osteoclast differentiation even in the absence of RANKL.35 The in vivo observation that deletion of NFATc1 in young mice results in osteopetrosis owing to impaired osteoclastogenesis also supported the important role of NFATc1 in osteoclasts.64
CALCIUM SIGNALING AND COSTIMULATORY SIGNALING FOR RANK
The activation of most NFAT transcription factor family members (NFATc1/c2/c3/c4) is originally regulated by calcium/ calmodulin signaling. In fact, since RANK does not seem to directly initiate calcium signaling and RANKL can only induce a partial activation of NFATc1 in osteoclast precursor cells, it has been suggested that costimulatory signaling for RANK may cooperate with RANKL to induce full activation of NFATc1 through calcium signaling pathways.35 It has been shown that tyrosine-based activation motif (ITAM)-bearing molecules such as DNAX-activating protein 12 (DAP12) and Fc receptor common γ chain (FcRγ) mediate calcium signaling and activate NFAT in immune cells.65 In osteoclasts, DAP12 and FcRγ also play an important role in the activation of NFATc1 through calcium signaling pathways. The severe osteopetrotic bone phenotype of mice doubly deficient in DAP12 and FcRγ suggests that immunoglobulin-like receptors associated with DAP12 and FcRγ are critical for osteoclast differentiation.910 DAP12 is associated with triggering receptor expressed in myeloid cells (TREM) 2 and signal-regulator protein β1 (SIRPβ1), whereas FcRγ interacts with OSCAR and paired immunoglobulin-like receptor (PIR-A) in osteoclasts.9 RANKL-mediated phosphorylation of ITAM through an unknown mechanism results in the activation of Syk and PLCγ. Activated PLCγ mobilizes intracellular calcium, which in turn activates the calmodulin-dependent phosphatase calcineurin. Calcineurin directly dephosphorylates serine residues in NFATc1, allowing for its rapid translocation into the nucleus and subsequent activation. Recently, specific tyrosine kinases that may provide a link between ITAM and RANK signaling were suggested. RANKL-mediated activation of Tec family tyrosine kinases such as BTK and Tec leads to phosphorylation of PLCγ to release calcium from the endoplasmic reticulum.66 Tec and BTK double-deficient mice develop an osteopetrotic phenotype, suggesting that these two kinases are associated with the regulation of osteoclastogenesis.66
CONCLUSION
Increased osteoclast formation is involved in bone diseases including osteoporosis and rheumatoid arthritis. Therefore, study of signaling pathways regulating osteoclast differentiation is crucial for a thorough understanding of the skeletal system in pathological conditions. Although significant signaling pathways for osteoclast differentiation have been elucidated, future studies of delicate regulatory networks involved in bone homeostasis are required for development of useful therapeutic strategies. Notably, future studies should focus on investigating the exact mechanisms underlying RANKL-mediated activation of costimulatory signals for RANK.
 XML Download
XML Download