

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Pseudorabies virus (PRV) is a porcine alpha-herpes virus and is the etiological agent of Aujeszky's disease, which produces neurological disorders, respiratory ailments, and reproductive failure in pigs, its natural host [1234]. However, despite successful PRV vaccination campaigns that have been conducted worldwide in an attempt to ensure disease eradication, pseudorabies outbreaks have still occurred in swine populations in recent years [56]. PRV has the ability to establish a lifetime latent infection in the trigeminal ganglia of the peripheral nervous system of infected pigs, and latently infected pigs can act as potential reservoirs for virus reactivation, shedding, and dissemination in susceptible swine populations [7]. During the latency period, it has been determined that a large latency transcript (LLT) is involved in maintaining the latent stage of the PRV [8].
MicroRNA (miRNAs) are small non-coding RNAs (21 to 24 nt), which regulate mRNA expression at the post-transcriptional level, either by inducing mRNA degradation or by suppressing mRNA translation [910]. Some reports have proposed that miRNAs have critical roles in host-virus interactions. More recent reports have indicated that miRNAs have the ability to regulate virus replication and host immune responses by modulating the expressions of viral and host genes associated with virus replication [1112]. In addition, it has been determined that several herpesviruses, such as Epstein Barr Virus, Bovine Herpesvirus 1 (BoHV-1), and Herpes Simplex Virus, as well as other herpes viruses, contain miRNAs that regulate their own cycles [131415]. In addition, several viruses are known to be able to harness host miRNAs in order to facilitate their replication [161718]. Therefore, it is not surprising that many viral-encoded miRNA molecules have been shown to potentially block host immune recognition, activation, and viral antigen presentation processes in order to escape from the host's immune responses and to facilitate viral replication [1920].
In recent years, it has been reported that miRNAs encoded by several alpha-herpes viruses often have been observed to be clustered in the viral genome, mapped within the latency-associated transcript locus, or clustered in adjacent regions [13142122]. With regard to the PRV, clusters of 11 miRNA genes have been identified using deep-sequencing techniques in immature porcine dendritic cells, as well as in a porcine kidney cell line (PK15), during lytic infections [22]. In our previous study, we reported that there was a cluster of 8 miRNA genes of PRV, along with 10 differentially expressed cellular miRNAs, in infected mouse neuro-2a cells [21]. However, the detailed function of the LLT miRNA cluster required further research as it has been previously reported that the target genes included both viral and host genes.
In the present study, it was observed that the PRV-encoded prv-miR-LLT11a was first downregulated and then upregulated in PRV-infected PK15 cells, which may have significantly repressed PRV replication. This study hypothesized that prv-miR-LLT11a may be associated with latent infections of the PRV. Our investigations demonstrated that the swine leukocyte antigen (SLA)-1 gene, which was the target gene of prv-miR-LLT11a, could be downregulated in a dose-independent manner by the overexpression of prv-miR-LLT11a. The obtained data not only provided a new insight into PRV-host interaction but also suggested a potential antiviral strategy against future PRV infections.
MATERIALS AND METHODS
Cells
In this study, PK15 cells were cultured in Dulbecco's modified Eagle's medium (Gibco, USA) that had been supplemented with 10% fetal bovine serum (HyClone, USA) and penicillin-streptomycin (Life Technologies, USA). PRV QXX, which is a variant PRV strain isolated in 2012, was utilized in this study [2123]. The PK15 cells were cultured to approximately 80% confluence and then infected with PRV QXX at a multiplicity of infection (MOI) of 1. The infected cell cultures were harvested at different post-infection times.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) method
For the detection of the prv-miR-LLT11a, the previously described quantitative stem-loop qRT-PCR method was utilized [24]. Then, the total RNA extracted from the PRV-infected PK15 cells at different times was reverse transcribed. The qRT-PCR was performed with a prv-miR-LLT11a-specific forward primer 5′-ACACTCCAGCTGGGAGGCTGGGAGTGGGG-3′ and a reverse primer 5′-TGGTGTCGTGGAGTCG-3′. In the current study, the U6 gene served as an internal reference as follows: Forward 5′-TTATGGGTCCTAGCCTGAC-3′ and reverse 5′-CACTATTGCGGGTCTGC-3′.
For the detection of the mRNA level of the SLA-1 gene, total RNAs were extracted from the PK15 cells post-transfection with prv-miR-LLT11a, or post-infection with PRV, at different times and then reverse transcribed. At that point, the qRT-PCR was performed with a specific forward primer 5′-AAGTCAAGGAAACCGCACAG-3′ and reverse primer 5′CAAGTA GCAGCCAAACATGC 3′ for SLA-1. In this study, β-actin mRNA was used as an internal reference.
Plasmids
The full-length 3′UTR or 3′UTR-mut of SLA-1 were cloned into a 3′UTR luciferase reporter plasmid pmirGLO (Promega, USA). Then, the SLA-1-3′UTR was amplified using specific primers: SLA-1-3′UTR forward 5′-CGAGCTCGGTGGCTGGAGTTGTGATCTGGAGG-3′ and reverse 5′-CTCTAGAGTTCTCAATCCTTCCATTTATTTCC-3′. The resultant 533 bp products were ligated into the SacI and XbaI of the pmirGLO vector. These were referred to as pmirGLO-SLA-3′UTR. The SLA-1 mutant 3′UTR recombinant plasmid was generated by overlapping the PCR with the primers 5′-CTGGCCCAGAACTGCCCCCCAC-3′ and 5′-CAGTTCTGGGCCAGGGTCCCCAC-3′ and was termed pmirGLO-SLA-3′UTR-mut. These constructions were then validated by using restriction cleavage and sequencing processes (Huada Gene, China).
Dual luciferase assay method
In the present study, in order to determine whether the prv-miR-LLT11a directly targeted the SLA-1 gene, synthesized prv-miR-LLT11a or miR-NC mimics, along with the recombinant plasmids pmirGLO-SLA-3′UTR or pmirGLO-SLA-3′UTR-mut, were co-transfected. Then, at 48-h post-transfection (hpt), the cell lysates were harvested for the luciferase assay that used a dual luciferase assay kit (Promega) and a TD-20/20 Luminometer (Turner Designs, USA). The values of each of the obtained samples were normalized using the Renilla luciferase value and were then expressed as relative luciferase activities (firefly luciferase/Renilla luciferase) in order to evaluate the interactions between prv-miR-LLT11a and SLA-1. Each of the sampling processes was performed in triplicate in 3 independent experiments.
Western blot analysis
Proteins were extracted from the PK15 cells using a lysis buffer, separated using a 10% polyacrylamide gel, and blotted onto a PVDF membrane. The membrane was probed with a goat anti-SLA polyclonal antibody (Santa Cruz, USA) at a 1:500 dilution, or by using β-actin at a dilution of 1:1,000 for 2 h at room temperature. This was followed by incubation with horseradish peroxidase-conjugated donkey anti-goat IgG at a 1:2,000 dilution (Santa Cruz). The signal was then visualized with an enhanced chemiluminescence reagent.
Statistical analysis
In the current study, all of the measured values between the different groups were analyzed using Student's t-test. The data are expressed as mean ± standard deviation values. The error bars represented the standard deviations from the mean of the 3 replicates, and values of p < 0.05 were considered to be of statistical significance.
RESULTS
Expression profiles of prv-miR-LLT11a during PRV infection
Our previous research regarding small RNA libraries from PRV-infected cells using Illumina deep sequencing had shown that the PRV-encoded prv-miR-LLT11a had been induced by the PRV infection [21]. However, in the current study, in order to validate those results, the expression profiles of prv-miR-LLT11a in the PRV-infected PK15 cells were determined by using a stem-loop qRT-PCR process. First, it was observed that the expression level of prv-miR-LLT11a had been upregulated significantly after 1 h, and then downregulated between 2 and 6 h later, as shown in Fig. 1A. Also, the expression level was reduced to a minimum at 6 hpt, but then had gradually increased along with replication of the PRV. It was also observed that the abundance of prv-miR-LLT11a in the PK15 cells that had been infected with PRV had increased by 34-, 40-, and 94-fold at 8, 12, and 24 hpt, respectively, when compared with the non-infected controls. The results demonstrated that the PRV infection had affected the prv-miR-LLT11a expression, suggesting that prv-miR-LLT11a had a potential regulatory role during the PRV infection.
Fig. 1
PRV infection affects prv-miR-LLT11a expression. PK15 cells were infected with PRV (MOI = 1) and harvested at 0, 1, 2, 4, 6, 8, 12, and 24 h post-transfection. (A) The expression levels of prv-miR-LLT11a were detected by using a stem-loop qRT-PCR method. (B) One-step growth curve for PRV replication in PK15 cells. The fold changes in the mRNA levels were calibrated to the level of U6 using a 2−ddCt method. The data are representative of 3 independent experiments (mean ± standard deviation). Statistical significance of differences is indicated by ns.
PRV, pseudorabies virus; MOI, multiplicity of infection; qRT-PCR, quantitative reverse transcription polymerase chain reaction; ns, not significant.
*p < 0.05; **p < 0.01.

To analyze the replication dynamics of variant PRV strains, PK15 cells were infected with PRV. To establish a one-step growth curve for PRV, cells, as well as supernatants, were harvested in order to detect virus titers by plaque assay at the indicated times. As shown in Fig. 1B, PRV exhibited decreased virus titers at 1 hpt but increased virus titers starting at 2 hpt. Subsequently, the titers were reduced at 8 hpt and reached near plateau levels at 24 hpt. One possible reason for the change in expression level of prv-miR-LLT11a between 2 and 6 hpt is this was the period when the PRV entered into PK15 cells, which suggests that the differential expression of prv-miR-LLT11a in a PRV infection might be associated with PRV replication against the host immune response.
Overexpression of prv-miR-LLT11a causes inhibition of PRV replication
In the present study, for the purpose of determining the effects of the prv-miR-LLT11a on PRV replication, PK15 cells were transfected with synthesized prv-miR-LLT11a mimics or with miRNA negative control mimics (miR-NC) in order to cause overexpression and were then infected with PRV (MOI = 1) at 24 hpt. Cell supernatants were harvested at the indicated times, and virus titers were detected using a plaque assay process. As shown in Fig. 2A, transfection with prv-miR-LLT11a resulted in significantly reduced PRV titers at 21 and 27 hpt, when compared with the miR-NC transfection group. In addition, the effect of prv-miR-LLT11a transfection on cell viability in a cell counting kit-8 assay was analyzed, and the result showed that miR-LLT11a mimics transfection results were not significantly different from those from transfection with control mimics (Fig. 2B). These results indicate that prv-miR-LLT11a has the potential to suppress the replication of PRV in the PK15 cells.
Fig. 2
Effects of prv-miR-LLT11a on PRV replication. PK15 cells were transfected with prv-miR-LLT11a or NC mimics for 24 h and then infected with PRV at an MOI of 1. The supernatants were then harvested at different times and (A) viral titers were detected by using a plaque assay method. (B) Cell viability was determined by using a cell counting kit-8. The data are representative of 3 independent experiments (mean ± standard deviation).
NC, negative control; hpi, hours post infection; PRV, pseudorabies virus; MOI, multiplicity of infection.
Statistically significant differences are indicated by *p < 0.05; **p < 0.01.

SLA-1 mRNA as the target gene of prv-miR-LLT11a
In order to uncover the mechanism involved in prv-miR-LLT11a inhibition of PRV replication, this study focused on searching for the potential target of prv-miR-LLT11a. The SLA-1 gene was considered to be a potential target (Fig. 3A), as suggested by using the online software RNAhybrid (https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid/). However, in order to confirm that prediction, the SLA-1-3′UTR luciferase reporter plasmid (pmirGLO-SLA-1-3′UTR) was utilized, and a binding site mutation plasmid (pmirGLO-SLA-1-3′UTR-mut) was used as the negative control. These were then co-transfected into PK15 cells with prv-miR-LLT11a or miR-NC mimics. At 48 h post-transfection, the cell lysates were subjected to dual luciferase activities. As shown in Fig. 3B, the prv-miR-LLT11a mimics significantly suppressed the luciferase activities of the pmirGLO-SLA-1-3′UTR when compared with that of the SLA-1-3′UTR-mut. In contrast, the luciferase activities of the pmirGLO-SLA-1-3′UTR-mut were observed to have not been inhibited in the cells transfected with SLA-1-3′UTR or SLA-1-3′UTR-mut. The results demonstrated that the prv-miR-LLT11a had the ability to directly target the 3′UTR of the SLA-1 mRNA.
Fig. 3
Prv-miR-LLT11a targets the 3′UTR of SLA-1. (A) Predicted target sites of prv-miR-LLT11a in the 3′UTR of SLA-1; the SLA-1 miRNA sequences are labeled in red; (B) PK15 cells were transfected with SLA-1-3′UTR or SLA-1-3′UTR-mut luciferase reporters and 40 pmol of prv-miR-LLT11a or miR-NC mimics for 36 h. The cell lysates were harvested to determine reporter activity levels by using a luciferase assay method. The data are expressed as relative firefly luciferase activity normalized to Renilla luciferase activity. The data are presented as mean ± standard deviation values from 3 independent experiments.
SLA, swine leukocyte antigen; NC, negative control.
Statistically significant values are indicated by *p < 0.05.
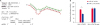
SLA-1 expressions downregulated by the prv-miR-LLT11a
In the present study, in order to further determine the regulatory effects of prv-miR-LLT11a on SLA-1, PK15 cells were transfected with prv-miR-LLT11a or miR-NC mimics 24 h prior to being infected with PRV. Subsequently, the mRNA and protein levels of SLA-1 were examined using qRT-PCR and western blot processes, respectively. As shown in Fig. 4A and B, transfection of the prv-miR-LLT11a mimics resulted in a significant decrease in the mRNA and protein levels of SLA-1 when compared with those of miR-NC-treated cells. Moreover, the effects of prv-miR-LLT11a on the expressions of the SLA-1 genes in the PRV-infected PK15 cells were determined, and it was determined that the inhibition efficiency on SLA-1 expressions was higher in infected PK15 cells than in uninfected PK15 cells. Therefore, it was speculated in this study that the inhibition effects of prv-miR-LLT11a on SLA-1 during a PRV infection may be induced via other pathways.
Fig. 4
Overexpression of prv-miR-LLT11a reduces the mRNA and protein levels of SLA-1. (A, B) PK15 cells were transfected with 120 pmol of prv-miR-LLT11a or miR-NC mimics or infected with PRV at an MOI of 1. The mRNA (A) and protein (B) levels of SLA-1 were measured at different times after transfection or infection by using qRT-PCR and western blot methods, respectively. (C, D) PK15 cells were transfected with different doses of prv-miR-LLT11a mimics for 48 h. The mRNA (C) and protein (D) levels of SLA-1 were measured by using qRT-PCR and western blot methods, respectively and β-actin was used as the internal reference control. The data are presented as mean ± standard deviation values from three independent experiments.
SLA, swine leukocyte antigen; MOI, multiplicity of infection; qRT-PCR, quantitative reverse transcription polymerase chain reaction; PRV, pseudorabies virus; ns, not significant.
Statistically significant values are indicated by *p < 0.05; **p < 0.01.

Subsequently, this study examined whether different doses of prv-miR-LLT11a could influence SLA-1 expressions. As shown in Fig. 4C and D, there were no significant differences observed in the mRNA and protein levels of SLA-1 during transfections with different doses of prv-miR-LLT11a mimics. Therefore, the results indicate that prv-miR-LLT11a can successfully downregulate both the mRNA and protein levels of SLA-1 in a dose-independent manner.
TAP1 transcriptions downregulated by prv-miR-LLT11a
The peptide loading complex (PLC) includes several proteins including tapasin (a transporter associated with antigen processing [TAP]); endoplasmic reticulum protein 44 (ERp44); calnexin (CNX); and calreticulin (CRT). The components of PLC have vital roles in cellular immune functions by synergizing with SLA-1. In the current study, for the purpose of investigating the effects of prv-miR-LLT11a on PLC expressions, as well as the synergistic roles of PLC and SLA-1 during antigen presentations, PK15 cells were transfected with prv-miR-LLT11a or miR-NC mimics at 12 h prior to being infected with PRV. Total RNA was extracted at the indicated post-infection times, and the transcript levels of the TAP1, CRT, CNX, ERp44, and tapasin genes were assessed by performing qRT-PCR (primers as shown in Table 1). The results showed that prv-miR-LLT11a overexpression had significantly inhibited the mRNA levels of the TAP1 gene. However, no significant changes were observed in the mRNA levels of the CRT, CNX, tapasin, and ERp44 genes (Fig. 5A).
Table 1
The sequences of primers used in this study

Fig. 5
Overexpression of prv-miR-LLT11a downregulated TAP1 expression. (A) PK15 cells were transfected with prv-miR-LLT11a or miR-NC mimics. The mRNA (A) levels of tapasin, ERp44, CRT, TAP1, and CNX were measured at different times after transcription by using a qRT-PCR method. (B, C) PK15 cells were infected with PRV or transfected with different doses of prv-miR-LLT11a mimics for 48 h. The mRNA (B) level of TAP1 was measured by using a qRT-PCR method; (C) the effects of PRV infection on the TAP1 expression levels were evaluated by using qRT-PCR with β-actin used as the internal reference control. The data are presented as mean ± standard deviation values from three independent experiments.
NC, negative control; TAP, transporter associated with antigen processing; CRT, calreticulin; CNX, calnexin; ns, not significant; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
Statistically significant values are indicated by *p < 0.05; **p < 0.01.

Moreover, the inhibitory effects of the different dosages of prv-miR-LLT11a on the TAP1 gene had indicated no significant differences, which suggested that the inhibition effects on the TAP1 gene were dose-independent (Fig. 5B). The results of this study's further investigations showed the TAP1 gene was significantly downregulated during PRV infection of PK15 cells (Fig. 5C); the mechanism(s) involved will be further investigated in further studies.
DISCUSSION
In recent years, an increasing amount of evidence has demonstrated that virus-encoded miRNAs have pivotal roles in viral infections by regulating gene expression at the post-transcriptional level [2526]. There have been numerous reports presented which have indicated that miRNAs have vital roles in the virus-cell interactions of PRV and its hosts [272829]. Our previous study identified 8 PRV-encoded miRNAs that were mainly located in the LLT region of the PRV [21]. However, reports elucidating their functions during PRV infections are limited. Therefore, given the importance of the functions of virus-encoded miRNAs in virus infections, this study focused on determining the functions of prv-miR-LLT11a during PRV replication in PK15 cells.
MiRNA expression profiles, which reflect the vital characteristics of a virus, are related to viral latency and host immune responses [30]. In our previous study, it was determined that high levels of miRNAs were expressed during the early stages of a PRV infection, suggesting that the miRNAs may be associated with viral latency. In the current study, the expression levels of prv-miR-LLT11a in PRV-infected cells were monitored. It was observed that there were significant differences in the early infection stages following inoculation of the PRV (Fig. 1A), implying that prv-miR-LLT11a had a role during the early stages of a PRV infection. Meanwhile, the one-step growth curve of PRV replication exhibited increased titers at 2 and 6 hpt. The detailed mechanisms associated with the PRV replication and the prv-miR-LLT11a differential expression require verification from further research. Moreover, the overexpression of prv-miR-LLT11a was observed to inhibit PRV replication (Fig. 2), which may also be related to the latency characteristics of PRV.
Target gene predictions, as well as verifications with dual luciferase assays, are generally used to confirm the functions of miRNAs [31]. The current study demonstrated that SLA-1 was a potential target gene of prv-miR-LLT11a by using RNAhybrid software based on the miRNAs and target gene's secondary structures; in addition, a dual luciferase assay method was utilized for verification of results. In previous study, SLA class 1 molecular (SLA-1) genes have been shown to have pivotal roles in specific cellular immune responses against viral infections [32]. Furthermore, it was reported that SLA-1-mediated antigen presentation pathways can be antagonized by PRV to evade the immune responses of the hosts [33]. Moreover, recent studies have revealed that a prevalent Chinese PRV variant could downregulate SLA-1 on the surface of PK15 cells and had targeted the cells for degradation through lysosome pathways [34].
In the present study, it was observed that prv-miR-LLT11a had the ability to inhibit the expression of SLA-1, implying that prv-miR-LLT11a may have downregulated SLA-1 expression through either mRNA degradation or translation repression [30]. Previous studies have reported that miRNAs play important roles in viral replications and virus-host interactions [3035]. Also, it was determined that viral miRNAs could either regulate viral replications or repress antiviral responses in infected cells [363738]. The results of the current study demonstrate that the PRV-encoded prv-miR-LLT11a targeted the 3′UTR of SLA-1, which is known to be associated with immunity and host defense responses to PRV infection. Therefore, it was assumed that prv-miR-LLT11a had inhibited the antiviral functions of the SLA-1 gene and could potentially promote replication of the PRV in some situations.
As previously mentioned, TAP is a pivotal element within the adaptive immune system, and it has been suggested that viruses have evolved complicated strategies to evade host immune surveillance strategies by interfering with TAP functions. Most recently, it was reported that the UL49.5 protein of BoHV-1 and equine herpesvirus 1 (EHV1), as well as PRV, can suppress MHC I cell surface expressions by blocking TAP functions [33]. TAP functions can be inactivated by the inhibition of peptide translocation of transporters due to interaction with the UL49.5 protein. This inhibition is independent of the non-specific mRNA cellular shut-off produced by UL41 protein [39]. In this study, the obtained data revealed that the overexpression of prv-miR-LLT11a had significantly inhibited TAP1 expression. These results suggest that PRV may regulate antigen transportation from the cytoplasm to the endoplasmic reticulum by the encoding of miRNAs; as a result, PRV is able to influence the immune responses of the host.
In summary, the present study demonstrated that prv-miR-LLT11a-related differential expressions were induced in response to a PRV infection in PK15 cells. It was determined that overexpression of prv-miR-LLT11a inhibited PRV replication. It was also observed that prv-miR-LLT11a directly targeted the 3′UTR of the SLA-1 gene and had downregulated the mRNA and protein expressions of SLA-1. These findings will not only potentially benefit further research regarding the functions of PRV-encoded miRNAs in viral replication, but also provide new insights into the interactions between viral infections and hosts.
 XML Download
XML Download