

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Globally, lung cancer is the second most prevalent cancer worldwide and a major cause of cancer-related deaths, with a prognosis that is among the worst [1]. In China, it tops the list for both incidence and mortality rates [2]. Lung cancer is clinically classified into non-small cell lung cancer (NSCLC)—encompassing adenocarcinoma, squamous cell carcinoma, and large cell carcinoma—and small cell lung cancer [3]. Over 85% of lung cancer cases fall under the category of NSCLC [4], among which lung adenocarcinoma (LUAD) is a frequently encountered subtype [5]. The early-stage symptoms of LUAD are typically not prominent, meaning that by the time of hospital admission, the cancer is often in an advanced stage with local or systemic metastasis, which is a key reason for the poor 5-year survival rate [6]. To this day, chemotherapy remains the primary option for advanced LUAD [4], with platinum-based combination therapies favored as the first-line treatment plan offered to patients [7,8]. Unfortunately, the presence of chemotherapy resistance cripples the efficacy of chemotherapy, greatly hindering the treatment of LUAD patients. Specifically, cancer cells evade the effects of chemotherapy by minimizing the uptake of platinum, rendering it inactive through associations with glutathione and metallothioneins, and quickening DNA repair process. These resistance strategies markedly undermine the potency of chemotherapeutic drugs [9]. The reprogramming of tumor metabolism is a special catalyst in the development of chemoresistance [10]. Hence, clarifying the mechanisms of tumor metabolism in chemoresistance is of great clinical relevance for advancing the treatment of LUAD patients.
A widely accepted perspective is that, even in oxygen-sufficient environments, cancer cells will engage in glycolysis for ATP production, offering them a survival advantage in hypoxic settings [11]. Also, an upregulation of glycolysis has become a definitive sign of cancer [12]. An array of research has shown that interventions in glycolytic metabolism can affect the initiation and progression of various cancers, such as hepatocellular carcinoma (HCC) [13], pancreatic cancer [14], and breast cancer [15]. In HCC, the transcription factor MKL-1 facilitates the progression of liver cancer by targeting PINK1-AS expression and modulating the ALDOA-mediated glycolysis pathway [13]. Ye et al. [16] reported that MicroRNA-16-1-3p curbs breast cancer growth and spread by inhibiting the PGK1-driven Warburg effect. There is also a correlation between the level of glycolysis and chemoresistance in tumor cells. Lin et al. [17] discovered that the POU2F1-ALDOA pathway enhances glycolysis, contributing to the resistance of colon cancer cells to oxaliplatin. Likewise, a marked increase in glycolytic activity is detected in bladder cancer cells with gemcitabine resistance [18]. Hence, pinpointing biomarkers that impact glycolysis in tumor cells could be a viable method to enhance the chemosensitivity of cancer patients.
Hyaluronan-mediated motility receptor (HMMR, also known as RHAMM or CD168), is an intracellular microtubule-associated factor that influences spindle assembly by directing the movement of dyneins and kinesins. The signaling it mediates is known to contribute to cancer progression [19,20]. For example, HMMR is highly expressed in renal cell carcinoma, and its depletion markedly suppresses cancer cell proliferation [21]. In breast cancer, overexpression of HMMR, a vital oncogenic factor in cancer progression, is linked to unfavorable patient outcomes [22]. A multitude of studies have evidenced that HMMR contributes to the progression of LUAD, with its high expression markedly lowering patient survival rates, highlighting its candidacy as a prognostic indicator [23-25]. Furthermore, HMMR has been correlated with tumor cell stemness [26] and chemoresistance [27]. In light of this, we surmised that HMMR might be associated with the stemness and chemoresistance of LUAD cells, a mechanism which the present study is dedicated to deciphering.
Our investigation has validated the high expression levels of HMMR in LUAD and their association with adverse patient prognoses, along with the regulatory effects of HMMR on aerobic glycolysis, cell stemness, and chemoresistance in LUAD. On a mechanistic level, we have shown that HMMR stimulates aerobic glycolysis, thereby enhancing stemness and chemoresistance in LUAD cells. This insight suggests that HMMR is likely a significant clinical target for enhancing chemoresistance in LUAD. The research aims to provide theoretical references for the potential of HMMR as a therapeutic target for overcoming chemoresistance in LUAD.
Go to : 
METHODS
Patient samples
This study enrolled patients diagnosed with LUAD by pathology at Affiliated Yueqing Hospital, Wenzhou Medical University from May 2023 to December 2023, and both cancerous and adjacent tissues were collected. The fresh tissue samples obtained were rapidly frozen in liquid nitrogen and stored at –80°C. This research has been granted approval by the Affiliated Yueqing Hospital, Wenzhou Medical University Ethics Committee (IRB No. YQYY202400210), aligning with its ethical standards and the Helsinki Declaration. Each participant has provided written informed consent.
Bioinformatics analysis
The TCGA-LUAD public database was consulted to evaluate HMMR gene expression in both tumor and adjacent normal tissues, with findings shown in violin plots. We also looked into the survival times of patients with high and low HMMR expression in LUAD. GSEA detected the pathways enriched by HMMR, and the Pearson correlation coefficient is employed to analyze the relationship between HMMR and glycolytic genes. The TCGA dataset used is open to public and available for legitimate research use.
Cell culture, reagents, and transfection
Human normal lung epithelial cells (BEAS-2B) and two varieties of human LUAD cells (A549, PC9) were sourced from SunnBio (SNL-203/SNL-089/SNL-152). Cultivation of BEAS-2B and A549 was performed using Dulbecco’s Modified Eagle Medium (DMEM) complete growth medium comprising DMEM base medium, 10% fetal bovine serum (FBS), and 1% penicillin/streptomycin. The PC9 cells were maintained in RPMI-1640 complete growth medium, which includes RPMI-1640 base medium, 10% FBS, and 1% penicillin/streptomycin. Cultivation conditions for all cell lines were set at 37°C in an incubator with 5% CO2, with medium changed every two days.
si-HMMR, oe-HMMR, and their corresponding negative control plasmids were acquired from Ribobio. Lipofectamine 3000 (L3000015, ThermoFisher Scientific) facilitated the transfection of these plasmids into LUAD cells in the logarithmic growth phase with confluence levels of about 80%. The subsequent experimental procedures were carried out 24 h following transfection.
2-Deoxy-D-glucose (2-DG), available from MedChemExpress (HY-13966), acts as a glucose substitute to suppress glycolysis by affecting hexokinase.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
Total RNA was isolated using the Trizol reagent (15596026CN, ThermoFisher Scientific). The mRNA was reverse transcribed into cDNA (RR036Q, TAKARA). Subsequently, TB GreenPremix Ex Taq reagent (RR420A, Takara) was utilized, and the target gene amplification was performed on a real-time quantitative PCR system, yielding the respective Ct values. β-actin was employed as the internal control, with the relative expression levels calculated using the 2–△△Ct method.
The primers used are listed in Table 1.
Western blot (WB)
Cell samples were gathered and chilled on ice, then lysed using RIPA buffer (P0038, Beyotime). After centrifugation, the supernatant was transferred to new EP tubes. Protein was quantified with a BCA protein quantification kit (P0011, Beyotime). The samples were loaded into the SDS-PAGE gel (P1200, Solarbio) for protein electrophoresis and then transferred to a PVDF membrane with a transfer apparatus. The membrane was removed from the transfer setup and incubated in a 5% skim milk solution for 1 h with shaking. The membrane was washed with TBST for 10 min, repeated thrice, before the addition of primary antibodies rabbit anti-CD44, CD133, LDHA, PDK1, MYC, HMMR and β-actin (ab189524/ab222782/ab52488/ab202468/ab32072/ab124729/ab8227, Abcam) for overnight incubation at 4°C. After another round of TBST washes, the membrane was incubated with the secondary HRP Goat Anti-Rabbit IgG (ab6721, Abcam) at room temperature for 1–2 h. TBST was employed again to wash away unbound secondary antibodies. The EZ-ECL chemiluminescence reagents (P0018M, Beyotime) were combined in a 1:1 ratio, applied to the membrane for binding. The membrane was imaged and developed. Protein expression analysis was carried out using ImageJ software.
Spheroid formation assay
Cells from various groups that were in a good condition and at the logarithmic growth phase were subjected to standard digestion. After resuspension and three washes with serum-free medium, they were counted. Diluted to a concentration of 8 × 105 cells/ml, the cells were mixed well and resuspended in serum-free medium with 4 mg/ml insulin (11061-68-0, Sigma), 10 ng/ml FGF (GF003AF, Sigma), 20 ng/ml EGF (E5036, Sigma), and 10 µg/ml B27 supplement (17504044, ThermoFisher Scientific). Cells were cultured at 37°C with 5% CO2, with medium changed every 2–3 days. Spheroid formation of cells was observed and imaged under a microscope after two weeks.
CCK-8
In the drug sensitivity test, a CCK-8 reagent kit (C0040, Beyotime) to test cell viability following the manufacturer’s protocol. To elaborate, 24 h after transfection, 5,000 cells per well were dispensed into a 96-well plate and subjected to cisplatin treatments at concentrations ranging from 0 to 100 µM for 48 h at 37°C. The culture medium was then refreshed with a 10% CCK-8 reagent solution for additional 2 h-incubation at 37°C. The absorbance at 450 nm was detected using a microplate reader. The IC50 was calculated to gauge the sensitivity to cisplatin in different groups of cells.
Apoptosis analysis via flow cytometry
Using the Annexin V-FITC Apoptosis Detection Kit (C1062L, Beyotime), cell apoptosis was determined. To begin with, cells that were subjected to apoptotic stimuli were gathered and resuspended in phosphate-buffered saline (PBS). After counting the cells, 1 × 106 cells from each group were prepared, centrifuged, the supernatant was decanted, and the cells were resuspended in 500 µl of Annexin V binding buffer. Annexin V-FITC (5 µl) and propidium iodide (10 µl) were introduced, followed by vortex mixing and a 15-min incubation at room temperature (25°C) in darkness. The cells were then transferred to flow cytometry tubes for analysis of early and late apoptosis stages on a flow cytometer, with FlowJo software used for determining the proportion of apoptotic cells.
Measurement of extracellular acidification rate (ECAR) and oxygen consumption rate (OCR)
The Seahorse XF96 Extracellular Flux Analyzer facilitated the measurement of the OCR and ECAR in LUAD cells from different treatment cohorts. Specifically, cells were seeded at 2.5 × 104 cells per well in Seahorse XF Cell Culture Plates and cultured for 24 h in an incubator at 37°C with 5% CO2. The Seahorse XF Glycolysis Rate Assay Kit and Seahorse XF Cell Mito Stress Test Kit (103344-100/103015-100, Agilent Technologies) were employed to assess OCR and ECAR, respectively, guided by the kits’ manual. The data obtained were used to evaluate the glycolytic activity of the cells.
Glucose consumption, lactate production and ATP production measured by kits
The glucose content detection kit (BC2500, Solarbio) was used, and the working solution was prepared according to the kit’s manual. 1 ml distilled water was added per 5 million cells, and the cells were disrupted by ultrasonic waves (ice bath, 20% power, 3 sec on, 10 sec off, for 30 cycles). The sample was boiled for 10 min in a water bath, cooled, and then centrifuged at 8,000 g at 25°C for 10 min. The supernatant was taken for further use. Reagent I and distilled water were added to the sample in succession, vortexed to ensure homogeneity, and incubated at 37°C for 15 min, with the absorbance determined at a wavelength of 505 nm.
Guided by the L-Lactic Acid (L-LA) Assay Kit (BC2230, Solarbio) manual, 1 ml of extraction solution I was used for 5 × 106 cells, and ultrasonic cell lysis was conducted on ice (300 W, 3 sec on, 7 sec off, for 3 min in total). The sample was centrifuged at 4°C, 12,000 g for 10 min, and 0.8 ml of the supernatant was collected. Then, 0.15 ml of extraction solution II was carefully added, mixed gently to avoid bubbles, and centrifuged again at 4°C, 12,000 g for 10 min. The supernatant was set aside for testing, and absorbance was determined at 570 nm after complete interaction with the reagents.
As per the guidelines of the ATP Content Colorimetric Assay Kit (BC0300, Solarbio), 5 million cells were subjected to 0.9 ml of extraction reagent I, homogenized in ice water bath at 60 Hz for 90 sec, boiled for 2 min, and then cooled with running water. The mixture was centrifuged at 4°C and 10,000 g for 10 min, and the supernatant was collected and kept on ice for analysis. Enzymatic reaction: The working solution was added for a 30-min incubation. After adding Reagent V, the mixture was centrifuged at 10,000 g for 5 min, and the resulting supernatant was prepared for subsequent testing. Color development: The supernatant was transferred into microplate wells, to which the color developer working solution was subsequently added, mixed for 5 sec, and then left to stand at room temperature for 2 min. After that, reagent VIII was added, mixed for 5 sec, and allowed to stand for 5 min at room temperature. The OD values at 636 nm were determined using a microplate reader.
Statistical analysis
Each experiment was triplicated, and data are shown as mean ± standard deviation. Student’s t-test was applied for assessing differences between pairs of groups, and ANOVA for multiple groups. Statistical analysis was conducted using SPSS 26.0 (SPSS Software) and GraphPad Prism 8.0 (GraphPad Software) facilitated the charting. In addition, * indicates a significant difference with p < 0.05.
Go to :
RESULTS
High expression of HMMR in LUAD correlates with worsened patient outcomes
We downloaded and analyzed HMMR expression data from the TCGA-LUAD database, which showed a remarkable upregulation of HMMR in LUAD tissues compared to normal tissues (Fig. 1A). Leveraging bioinformatics, we assessed the correlation between HMMR expression levels and survival, discovering that patients with lower HMMR expression had a notably higher survival rate than those with higher HMMR expression (Fig. 1B), which is suggestive of a link between HMMR expression and poor patient prognosis. Following this, we collected LUAD tissues and corresponding paraneoplastic tissues from patients, finding a pronounced increase in HMMR mRNA levels in the cancerous tissues via qRT-PCR (Fig. 1C). In addition, HMMR expression levels gradually increased with cancer progression expression (Fig. 1D). The qRT-PCR examination of HMMR mRNA in normal epithelial cells (BEAS-2B) and LUAD cell lines (A549, PC9) further corroborated these findings (Fig. 1E). Overall, HMMR is overexpressed in LUAD, and this overexpression is associated with poor patient outcomes.
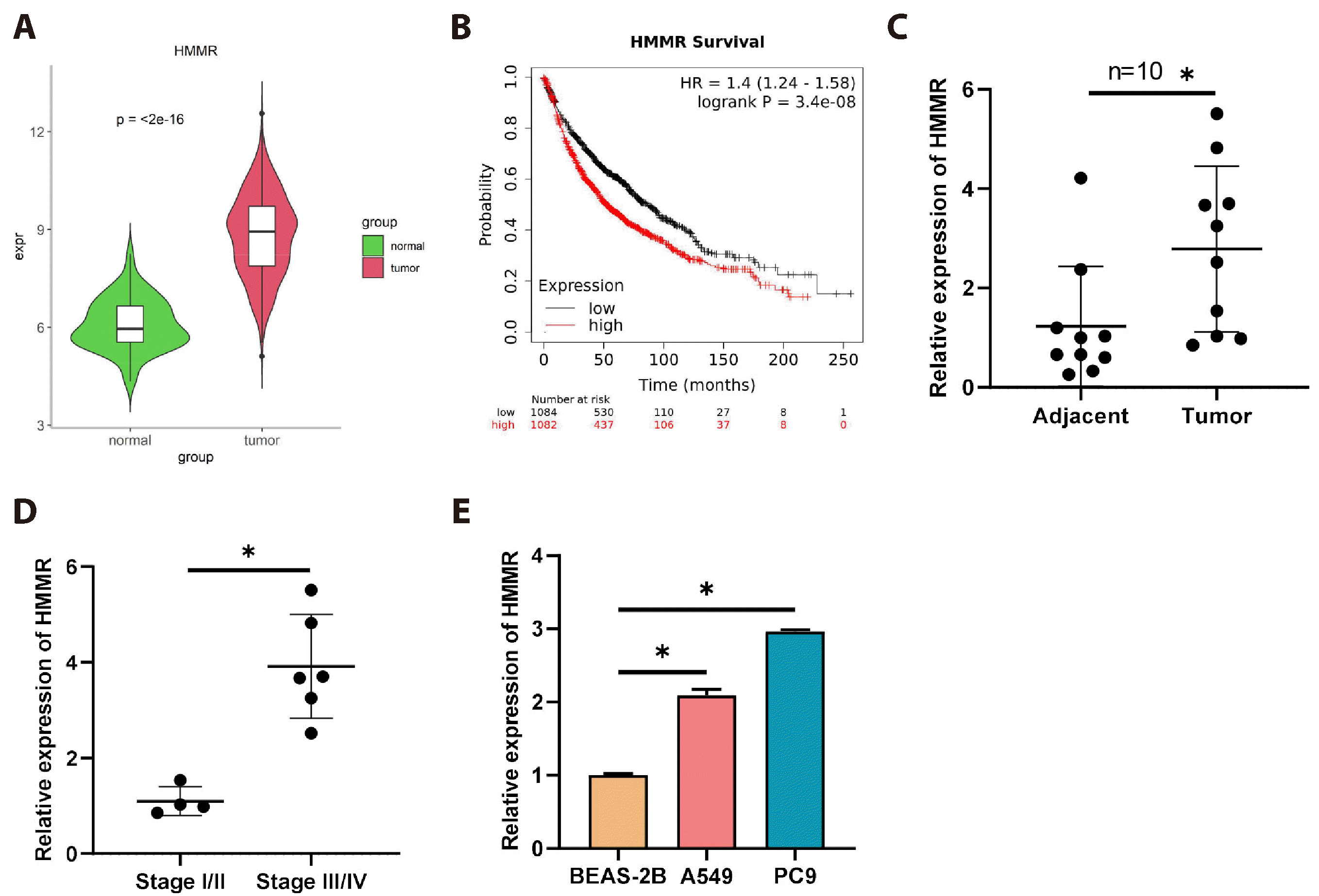 | Fig. 1High expression of hyaluronan-mediated motility receptor (HMMR) in lung adenocarcinoma (LUAD) correlates with worsened patient outcomes.(A) TCGA database analysis of HMMR expression in normal lung tissues (N = 59) versus LUAD tissues (N = 539). (B) K-M curve analysis of overall survival time in patients with high/low HMMR expression (N = 1,082/1,084). (C) qRT-PCR detection of HMMR mRNA expression in cancerous and adjacent tissues (N = 10/10). (D) qRT-PCR detection of HMMR mRNA expression in tumor tissues of stage I/II (N = 4) versus stage III/IV (N = 6) LUAD patients. (E) qRT-PCR evaluation of HMMR mRNA levels in normal human lung epithelial cells and LUAD cells (N = 3). Values are presented as mean ± SD. * signifies p < 0.05.
|
High HMMR expression enhances the stemness and drug resistance of LUAD cells
Existing literature has shown that HMMR correlates with tumor stemness [26] and chemoresistance [27], yet the connection between HMMR expression and these attributes in LUAD remain unexplored. This study is aimed at filling this gap. As our initial experiments have revealed high HMMR expression levels in the PC9 LUAD cell line, we transfected these cells with si-NC and si-HMMR to establish HMMR knockdown models, and the transfection efficiency was measured by qRT-PCR (Fig. 2A). Following the transfection, spheroid formation experiments was conducted to examine the stemness of cells in the various treatment groups. The data showed that HMMR knockdown remarkably impeded the spheroid formation efficiency of PC9 cells as compared to the control group (Fig. 2B, C). Additionally, WB detection of the stem cell markers CD44 and CD133 showed that their expression was inhibited in HMMR knockdown PC9 cells relative to the control group (Fig. 2D). The results collectively implied that the suppression of HMMR reduced the stemness of LUAD cells. To investigate how HMMR influences the resistance of LUAD cells to cisplatin, transfected PC9 cells with si-NC and si-HMMR were exposed to cisplatin, followed by a CCK-8 assay to measure and determine the IC50 values. The findings indicated lower IC50 values for cells with HMMR knockdown versus the control group (Fig. 2E). Furthermore, flow cytometry assessed apoptosis in si-NC and si-HMMR-transfected PC9 cells, showing that HMMR knockdown markedly promoted apoptosis compared to the control (Fig. 2F). Collectively, these results suggested that elevated HMMR expression fostered stemness and resistance in LUAD cells.
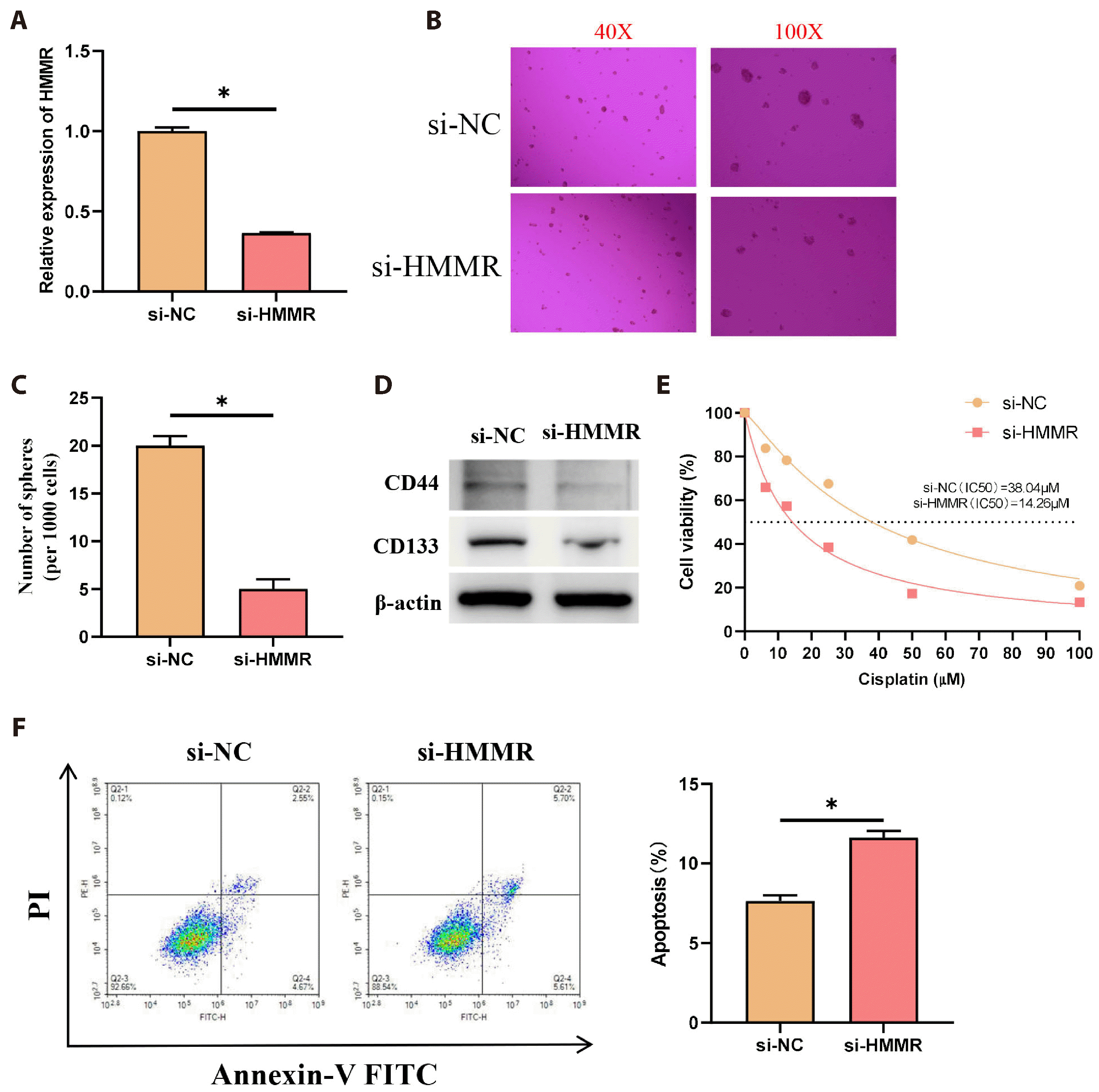 | Fig. 2High hyaluronan-mediated motility receptor (HMMR) expression enhances the stemness and drug resistance of lung adenocarcinoma (LUAD) cells.(A) si-NC and si-HMMR were transfected into PC9 lung adenocarcinoma cells, and transfection efficiency was confirmed by qRT-PCR (N = 3). (B, C) Spheroid formation detected stemness among different treatment groups (N = 3). More sphere formation represents more stemness of the cell. (D) WB examined the expression of stem cell markers CD44 and CD133. (E) The CCK-8 assay calculated the cisplatin IC50 values for each group (N = 3). Lower IC50 values indicate greater cisplatin sensitivity. (F) Apoptosis was evaluated using flow cytometry (N = 3). Values are presented as mean ± SD. WB, Western blot; PI, propidium iodide. * signifies p < 0.05.
|
HMMR expression mediates glycolytic signaling
To delve into the underlying mechanisms through which HMMR influences LUAD stemness and drug resistance, we performed the signaling pathways enriched by HMMR-related genes using GSEA single-gene enrichment The data demonstrated the pathway enrichment for HMMR in GLYCOLYSIS GLUCONEOGENESIS (Fig. 3A). Correlation analysis also showed a positive association between HMMR and expression of glycolytic genes (PKM, LDHA, SLC2A1, PDK1, MYC) (Fig. 3B). We thus proposed that HMMR expression could be orchestrating glycolytic signaling. Firstly, cells were allocated into si-NC and si-HMMR groups, where WB was employed to examine the expression of LDHA, PDK1, and MYC proteins, revealing that HMMR knockdown inhibited LDHA, PDK1, and MYC protein expression compared to the control levels (Fig. 3C). Later, the Seahorse XF96 system was applied for ECAR and OCR detection, which showed that HMMR knockdown considerably decreased ECAR and augmented OCR relative to the control group (Fig. 3D, E). Metabolic assays were then used to determine glucose consumption, lactate synthesis, and ATP levels in different groups. It was found that HMMR knockdown notably suppressed glucose consumption, lactate synthesis, and ATP levels in comparison to the control group (Fig. 3F-H). In conclusion, HMMR expression could facilitate aerobic glycolysis in LUAD cells.
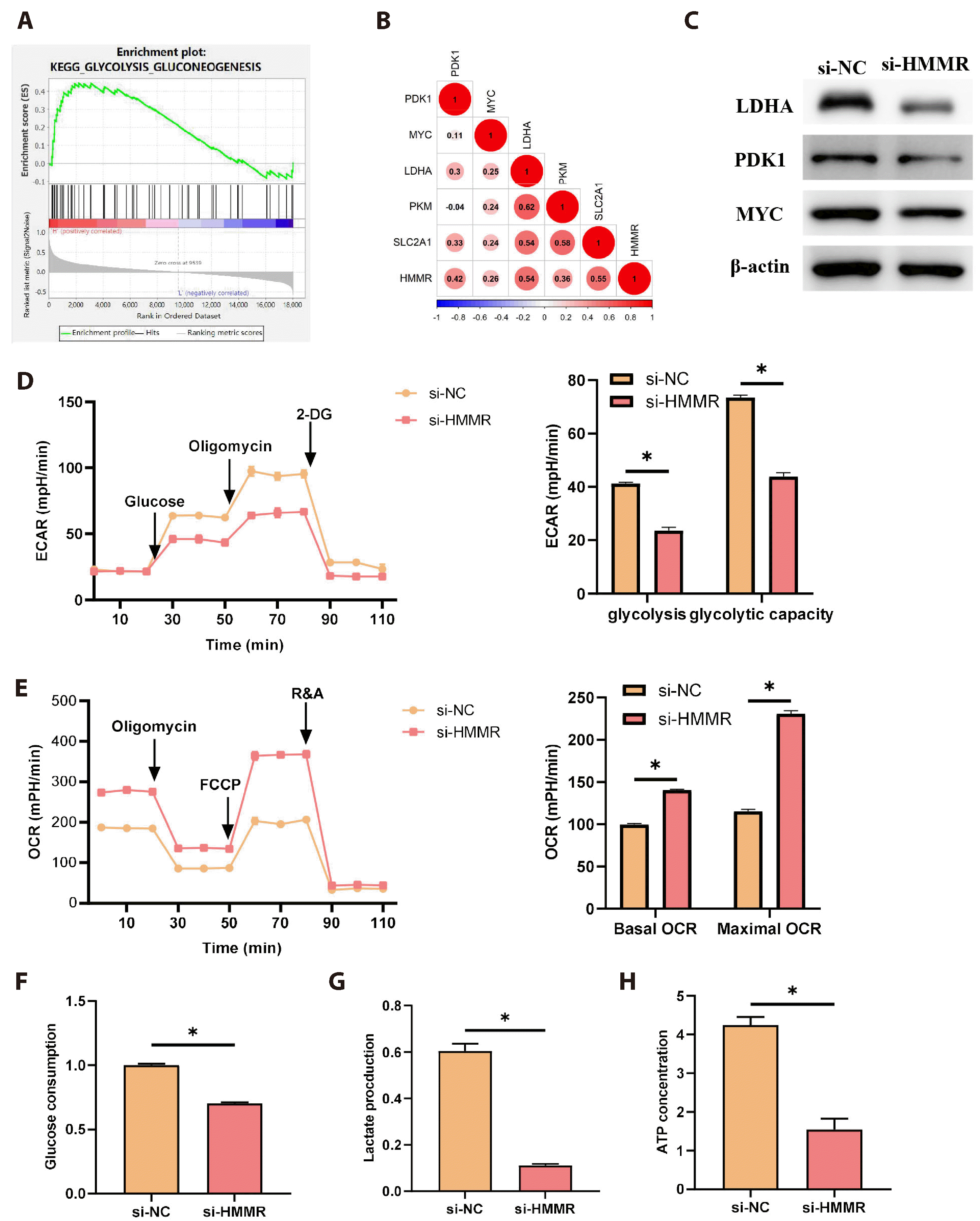 | Fig. 3Hyaluronan-mediated motility receptor (HMMR) expression mediates glycolytic signaling.(A) GSEA analysis of signaling pathways enriched for genes highly associated with HMMR. (B) Pearson analysis of the correlation between HMMR and key genes of glycolysis (PKM, LDHA, SLC2A1, PDK1, MYC). A larger value means a stronger correlation. (C) WB analysis of LDHA, PDK1, and MYC protein expression in si-NC and si-HMMR groups. (D) ECAR measurement via the Seahorse XF96 Extracellular Flux Analyzer (N = 3). ECAR levels are positively correlated with glycolysis. (E) OCR measurement via the Seahorse XF96 (N = 3). OCR levels are negatively correlated with glycolysis. (F) Glucose consumption measured in cell supernatants of different groups with a specific assay kit (N = 3). (G) Lactate synthesis quantified in the cell groups with a specific assay kit (N = 3). (H) ATP yield evaluated in the cell groups with a specific assay kit (N = 3). Values are presented as mean ± SD. WB, Western blot; ECAR, extracellular acidification rate; OCR, oxygen consumption rate; 2-DG, 2-Deoxy-D-glucos. * signifies p < 0.05.
|
HMMR enhances stemness and drug resistance of LUAD cells by promoting glycolytic metabolism
To substantiate that HMMR facilitates glycolysis, thus augmenting the stemness and drug resistance of LUAD cells, we set up distinct cell groups: oe-NC with PBS, oe-HMMR with PBS, and oe-HMMR with 2-DG (a glycolytic inhibitor). Considering the low HMMR expression in the A549 cell line as confirmed by prior experiments, we transfected A549 cells with oe-NC and oe-HMMR to establish an overexpression model of HMMR and validated the transfection efficiency via qRT-PCR (Fig. 4A). Afterward, 2-DG was applied to the cells for ECAR and OCR testing, with the data revealing that HMMR overexpression notably increased ECAR and decreased OCR when compared to the control group, and the addition of 2-DG reversed the effects induced by HMMR overexpression (Fig. 4B, C). Assay kits permitted the quantification of glucose consumption, lactate production, and ATP synthesis in different cell groups. The results demonstrated that HMMR overexpression led to a substantial increase in glucose utilization, lactate synthesis, and ATP output when compared to the control group, with these effects being reversed by 2-DG (Fig. 4D-F). WB results also show that compared to the control group, overexpression of HMMR significantly promotes the expression of glycolysis -related proteins LDHA, PDK1, and MYC, which is reversed by the addition of 2-DG (Fig. 4G). This evidence pointed out that HMMR overexpression accelerated glycolytic metabolism in LUAD cells. To determine the link between HMMR-induced glycolysis and the stemness and drug resistance of LUAD cells, we performed spheroid formation assays to test stemness in the different groups, finding that HMMR overexpression notably increased the spheroid formation efficiency of A549 cells, and this increase was counteracted by 2-DG (Fig. 4H). WB analysis of stemness indicators CD44 and CD133 showed that their expression was markedly enhanced by HMMR overexpression, and this enhancement was mitigated by 2-DG treatment (Fig. 4I). Additionally, cisplatin treatment followed by CCK-8 detection showed that overexpression of HMMR led to a marked increase in both the survival rate of LUAD cells and the IC50 value compared to the control group, which was counteracted by 2-DG addition (Fig. 4J). The subsequent flow cytometry analysis of apoptosis among the groups demonstrated that HMMR overexpression notably suppressed cell apoptosis, and this suppression was reversed by 2-DG (Fig. 4K). Overall, HMMR enhanced the stemness and drug resistance of LUAD cells by promoting glycolytic metabolism.
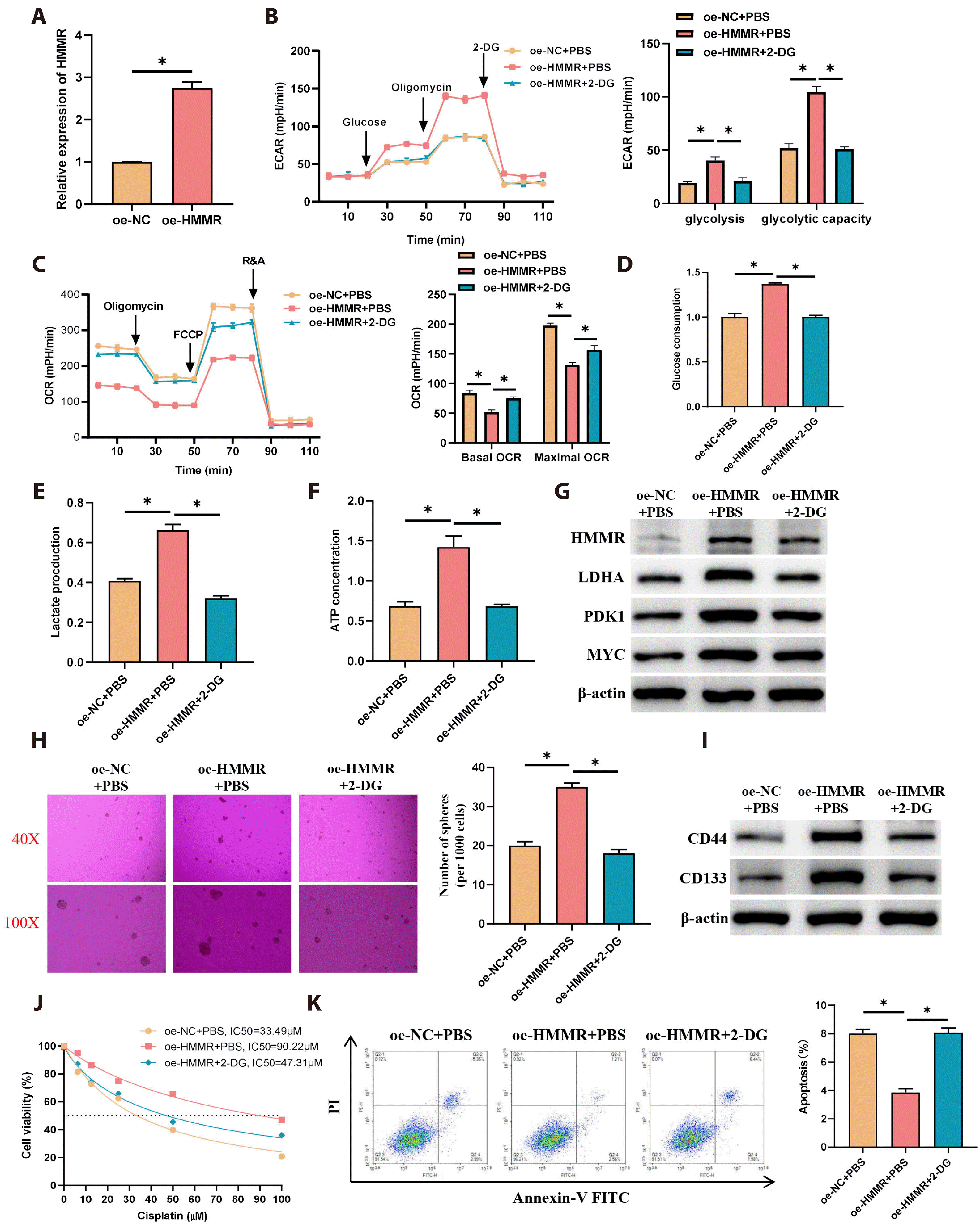 | Fig. 4Hyaluronan-mediated motility receptor (HMMR) enhances stemness and drug resistance of lung adenocarcinoma (LUAD) cells by promoting glycolytic metabolism.(A) Transfection of A549 cells with oe-NC and oe-HMMR, with qRT-PCR assessing transfection efficiency (N = 3). (B) Cells were divided into groups as oe-NC + PBS, oe-HMMR + PBS, oe-HMMR + 2-DG, with ECAR analyzed by the Seahorse XF96 (N = 3). (C) OCR measured with the Seahorse XF96 (N = 3). (D) Glucose consumption in supernatants of the groups assessed by a specific assay kit (N = 3). (E) Lactate production in the groups measured by an assay kit (N = 3). (F) ATP production in the groups evaluated with a specific assay kit (N = 3). (G) WB detection of HMMR and glycolysis-related protein (LDHA, PDK1, MYC) expression. (H) Spheroid formation assays for stemness in different groups of cells (N = 3). (I) WB detection of stem cell markers CD44 and CD133 expression. (J) CCK-8 assay for cisplatin IC50 in each group (N = 3). (K) Flow cytometry for apoptosis assessment in cells (N = 3). Values are presented as mean ± SD. PBS, phosphate-buffered saline; 2-DG, 2-Deoxy-D-glucos; ECAR, extracellular acidification rate; OCR, oxygen consumption rate; WB, Western blot; PI, propidium iodide. * signifies p < 0.05.
|
Go to :
DISCUSSION
LUAD, identified as a highly invasive malignant tumor, is a great menace to human health and even life [28]. Although chemotherapy remains the foremost therapeutic choice for advanced stages of LUAD, the emergence of chemoresistance in patients leads to unfavorable prognoses, thus lessening the clinical efficacy of treatments [29-31]. There is an urgent necessity to explore the intrinsic mechanisms of chemoresistance in LUAD and to develop innovative therapeutic strategies for overcoming chemoresistance and enhancing patient outcomes. The main purpose of this study is to elucidate the potential role of HMMR in LUAD chemoresistance. Numerous studies have showed that HMMR, when overexpressed, substantially accelerates tumor growth and metastasis [32,33]. Jiang et al. [34] and Li et al. [35] have confirmed that HMMR is considerably upregulated in LUAD, and its elevated expression is associated with increased cancer cell proliferation, migration, and invasion, as well as a negative impact on patient prognosis. Our investigation corroborates these findings. Additionally, there is research substantiating the correlation between HMMR expression and chemoresistance. For instance, HMMR can interact with FOXM1 to activate autophagy, which in turn enhances the level of chemoresistance in osteosarcoma [36]. In addition, HMMR was shown to activate the TGF-β/Smad2 signaling pathway to induce epithelial mesenchymal transition and cell stemness thereby conferring chemoresistance to gastric cancer cells [37]. In this work, we further validated the critical function of HMMR overexpression in cisplatin resistance.
Cancer stem cells, a rare but malignant cell population [38,39], have stem-like properties such as high proliferation, self-renewal, and continuous differentiation, and play a crucial role in tumor metastasis, relapse, and resistance to chemotherapy [39,40]. The overexpression of PLOD2 has been found to enhance cancer stem cell-like features in advanced laryngeal squamous cell carcinoma, fostering resistance to chemotherapy [41]. Zhang et al. [37] have shown that HMMR overexpression in gastric cancer confers cancer stem cell-like properties, thereby inducing chemoresistance. Our study has revealed that HMMR knockdown markedly inhibits cisplatin resistance in LUAD and attenuates stemness. Cancer stem cells typically depend on the reprogramming of metabolism to sustain their stem-like properties. Overexpression of FOXQ1, for example, has been shown to boost stemness in pancreatic cancer through the facilitation of aerobic glycolysis [42]. Similarly, in pancreatic ductal adenocarcinoma, P4HA1 is known to modulate cancer stemness via glycolysis [43]. Research by Kang et al. [44] highlights that HSP90 promotes the stemness of glioma by stimulating glycolysis, a process that can be counteracted by 2-DG. These findings imply a link between tumor cell stemness and glycolytic activity. These results are consistent with the findings of this study in LUAD cells. This study demonstrated that HMMR could promote the stemness characteristics of tumor cells and chemoresistance levels by activating glycolysis. We hypothesize that HMMR regulates the level of glycolytic metabolism through downstream molecular mechanisms, thereby conferring cisplatin resistance to tumor cells by influencing energy supply [45].
Glycolysis, a cancer identifier, critically impacts the growth and chemotherapeutic resilience of LUAD cells [46,47]. Gong et al. [48] revealed that the suppression of glycolysis activation can mitigate the cisplatin resistance in LUAD cells (A549) under hypoxic conditions. He et al. [49] further validated that PRDM14 can suppress the cisplatin sensitivity of LUAD cells by enhancing glycolysis. Therefore, it is evident that inhibiting glycolysis can reduce the proliferative capacity of LUAD cells and their resistance to chemotherapeutic agents. In our study, overexpressed HMMR in LUAD was highly enriched in the glycolytic signaling, reinforcing the notion that HMMR contributes to the cisplatin resistance and the inhibition of cell apoptosis in LUAD cells by orchestrating the glycolytic signaling cascade.
However, this study still has some limitations. First, due to the limited number of clinical samples, this study did not have sufficient control over variables such as age and smoking status. This limitation may result in a less comprehensive understanding of the clinical value of the HMMR target. Secondly, we were unable to identify the specific molecular mechanisms by which HMMR mediates changes in glycolytic metabolism levels, which requires us to dig deeper into possible upstream and downstream regulatory genes in subsequent studies. In addition, targeting HMMR to address chemoresistance may lead to potential side effects, including affecting stem cell proliferation [20,50] and neuronal development [51,52]. The resolution of these issues will provide a more reliable experimental basis for targeting HMMR for the treatment of LUAD.
Go to :
 XML Download
XML Download