

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
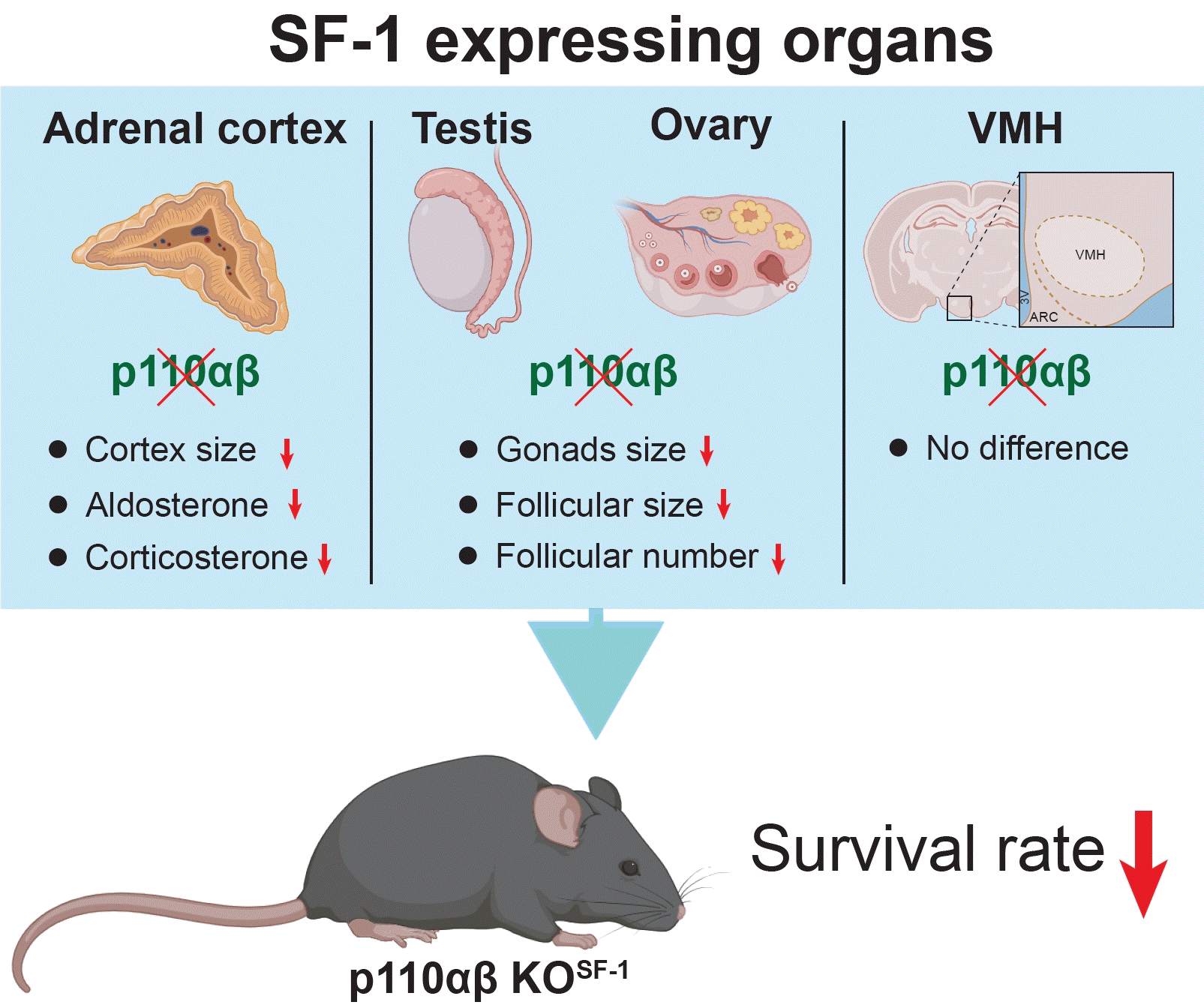
The nuclear receptor steroidogenic factor 1 (SF1, NR5A1) was initially recognized for its role in the development and function of various organs, including the ventromedial hypothalamus (VMH), adrenal glands, and gonads [1,2]. Mutations or deletions in the gene encoding SF1 have been associated with compromised development of these organs [3]. Accordingly, the ablation of SF1 has led to the absence of adrenal glands and abnormal sexual differentiation in both male and female mice [4].
Phosphatidylinositol 3-kinase (PI3K) signaling has been implicated in various pathways governing cellular proliferation and differentiation. Specifically, research suggests that PI3K signaling in neurons of the VMH is key to regulating energy homeostasis [5-7]. Moreover, PI3K is recognized as a direct downstream target of follicle-stimulating hormone, influencing granulosa cell differentiation and follicular maturation [8]. PI3K is categorized into three classes—class I, class II, and class III—based on structural distinctions, particularly in regulatory subunits, and differences in lipid substrate specificity [9,10]. Class IA PI3K, along with its catalytic and regulatory subunits, is essential for cell proliferation, differentiation, and energy metabolism [11-13]. Notably, mutations in the catalytic subunit p110β have been linked to reduced testicular size and impaired function [14]. In contrast, within oocytes, the overexpression of PI3K resulting from the deletion of the phosphatase and tensin homolog is associated with premature ovarian failure [15]. Furthermore, hyperactivation of the PI3K/protein kinase B (Akt)/mammalian target of rapamycin signaling pathway may contribute to the development of primary aldosteronism [16,17]. Previous research indicates that the deletion of the PI3K catalytic subunit p110α can result in altered energy metabolism under conditions of metabolic stress, without impacting the production of steroidogenic hormones [7,18].
The current literature indicates that PI3K, particularly its catalytic subunits, plays a functional role in SF1-expressing organs. These include the VMH, adrenal glands, and gonads. However, the deletion of only p110α in SF1-expressing cells has a small impact on peripheral tissues [7,19]. Given that class IA of PI3K comprises three distinct isoforms [20], we hypothesized that ablating both p110α and p110β in SF1-expressing cells may yield relevant insights. Our findings indicate that mice with a double knockout (KO) of these subunits, termed p110αβ KOSF1 mice, exhibited a decreased survival rate and impaired development of the adrenal cortex and gonads compared to their wild-type (WT) counterparts. However, no significant changes were observed in VMH development or SF1 levels. Overall, these data demonstrate the developmental importance of PI3K catalytic subunits in SF1-expressing cells.
Go to : 
METHODS
Animal and experimental protocols
Mice were housed under a 12-hour light/dark cycle, with room temperature controlled between 22°C and 24°C. They were fed a normal chow diet (3.82 kcal/g; Research Diets, New Brunswick, NJ, USA) and received water ad libitum. Genotyping was performed by extracting toe DNA and analyzing it using the REDExtract-N-Amp Tissue PCR Kit (Sigma-Aldrich, St. Louis, MO, USA). All animal experiments and surgical procedure were approved by the Institutional Animal Care and Use Committee (IACUC) of the Avison Biomedical Research Center, Yonsei University (Animal Protocol #2018-0056). The experimental design ensured a minimum of three mice per genotype, while aiming to minimize the number of animals suffering in the experiments. Mice with p110αβ KO in SF1-expressing cells were generated by crossing SF1-Cre mice [21] with floxed p110α and p110β mice [22,23].
Blood serum collection
To assess basal corticosterone and aldosterone levels, mice were separated and allowed to stabilize. Blood was collected from the tail the following day between 2:00 PM and 3:00 PM. Subsequently, mice that had been singly housed were grouped for 30 minutes, after which blood was drawn to determine corticosterone levels under stress conditions, as previously described [7].
Hormone measurement
All blood samples were maintained at room temperature for 4 hours before being centrifuged for 10 minutes at 12,000 rpm and 4°C. Subsequently, the blood serum was transferred to 1.5-mL microcentrifuge tubes and stored at −60°C until hormone concentration analysis using commercial enzyme-linked immunosorbent assay kits (corticosterone and aldosterone kits; Abcam, Cambridge, UK).
Postprandial glucose measurement and glucose tolerance test
To assess postprandial glucose levels, food was withdrawn from the animal cages for 2 hours. Then, blood samples were drawn from the tail vein for analysis. Glucose concentrations were determined using the glucose oxidase method with a Contour TS glucometer (Ascensia Diabetes Care UK Limited, Newbury, UK).
For the glucose tolerance test, mice were fasted overnight to minimize variability in baseline blood glucose levels, with water provided ad libitum. The experimental mice were housed individually and allowed to stabilize for 1 hour the following day before being administered 1 g/kg of glucose dissolved in saline. Subsequently, tail blood samples were collected at 0, 20, 40, 60, and 120 minutes post-administration. Glucose levels were measured using a Contour TS glucometer (Ascensia Diabetes Care UK Limited).
Histological analysis
For brain sample preparation, whole brains were collected following a cardiac perfusion procedure using a 10% neutral buffered formalin solution (Sigma-Aldrich). The brains were then dehydrated overnight in 20% sucrose and sectioned using a CM1850 clinical cryostat (Leica Microsystems, Wetzlar, Germany) to a thickness of 20 µm. For peripheral samples, adrenal glands and gonads were dehydrated in ethanol, cleared in xylene, and embedded in paraffin blocks following standard protocols. The paraffin blocks were then sectioned using an RM2145 rotary microtome (Leica Microsystems) to a thickness of 5 µm. To assess the general anatomy of the tissues, brain structures were visualized by Nissl staining, while peripheral tissues were stained with hematoxylin and eosin (H&E). The number of neurons and SF1-positive cells were quantified using ImageJ (National Institutes of Health, Bethesda, MD, USA) and Adobe Photoshop CS6 (Adobe, San Jose, CA, USA). The size and number of follicles in the mouse ovary were also measured using ImageJ.
Brain sections were initially mounted onto Fisherbrand Superfrost Plus slides (Thermo Fisher Scientific, Waltham, MA, USA), while paraffin sections were deparaffinized. Subsequently, all sections were subjected to antigen retrieval by boiling in a 0.01 M sodium citrate solution at pH 2.0. To block nonspecific binding, sections were treated with 3% bovine serum albumin (Sigma-Aldrich) and then incubated with primary antibodies—rabbit anti–SF1 at a dilution of 1:10,000 [24] and rabbit anti-tyrosine hydroxylase (TH) at a dilution of 1:1,000 (Merck Millipore, Burlington, MA, USA)—overnight at 4°C. For further analysis, sections were incubated with either biotinylated antibody (Vector Laboratories, Newark, CA, USA) or Alexa 488 anti-rabbit (Invitrogen, Carlsbad, CA, USA), and detection was performed using a TCS SP5 or ICC50E device (Leica Microsystems).
Data analysis
Data are presented as mean±standard error of the mean from multiple samples. The Student t test, two-way analysis of variance, and GraphPad Prism version 8.0 (GraphPad Software Inc., La Jolla, CA, USA) were used to evaluate differences. P values less than 0.05 were considered to indicate statistical significance.
Go to :
RESULTS
Impact of p110αβ deletion in SF1 cells on survival and glucose homeostasis
The deletion of the p110α and p110β subunits of PI3K in SF1-expressing cells, referred to as p110αβ KOSF1, substantially impacted survival and glucose metabolism. Both male and female p110αβ KOSF1 mice exhibited significantly reduced survival rates from an early age, with only 36.8% of male and 50% of female mice surviving beyond 36 weeks (Fig. 1A, B). Weekly body weight monitoring revealed that male p110αβ KOSF1 mice began to show significant weight loss at 12 weeks of age, while female p110αβ KOSF1 mice maintained a body weight similar to that of WT controls (Fig. 1C, D). Further analysis revealed that, despite comparable body weights at earlier stages, male p110αβ KOSF1 male mice exhibited elevated glucose levels and impaired glucose tolerance, suggesting a disruption in glucose homeostasis (Fig. 1E-G). These findings underscore the critical roles of p110α and p110β in survival, development, and energy balance.
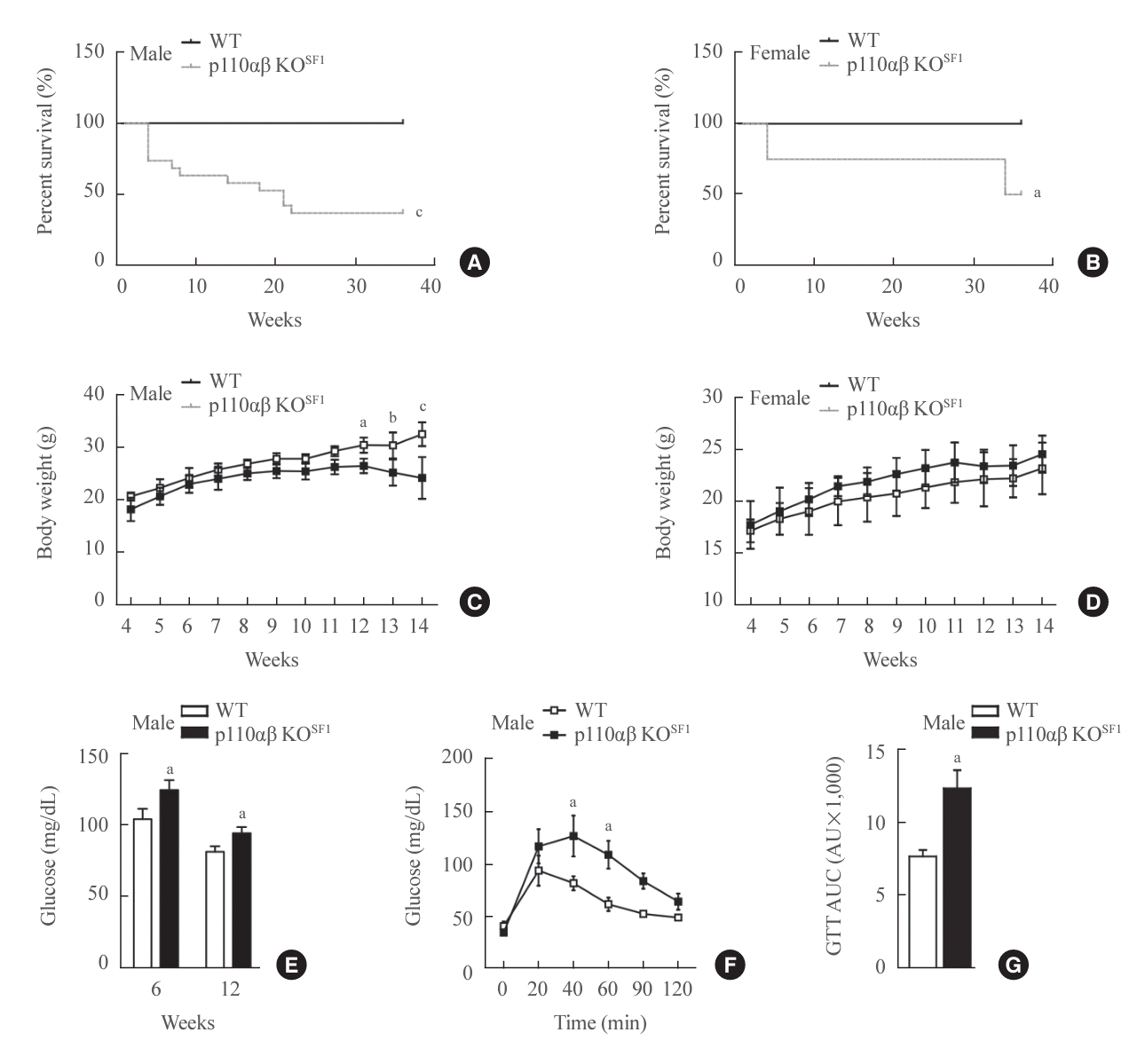 | Fig. 1.Phosphatidylinositol 3-kinase isoforms p110α and p110β in steroidogenic factor 1 (SF1)-expressing organs play key roles in the survival and normal development of mice. (A, B) Survival rates of (A) male and (B) female mice over a 36-week observation period (n=12 males and n=9 females for the wild-type [WT] group and n=19 males and n=8 females for the p110αβ knockout [KO]SF1 group). (C, D) Body weights of WT and p110αβ KOSF1 mice in both (C) males (n=4 for the WT group and n=6 for the p110αβ KOSF1 group) and (D) females (n=4 for each group). (E) Postprandial glucose levels in male mice at 6 and 12 weeks of age. (F, G) Glucose tolerance test (GTT) for male mice aged 6 to 10 weeks (n=7 for each group) (F) and the area under the curve (AUC) for the GTT (G). Data are presented as mean±standard error of the mean. AU, arbitrary unit. aP<0.05, bP<0.01, and cP<0.001 vs. WT group.
|
Unaltered VMH structure and SF1 expression in p110αβ KOSF1 mice
Given the central role of the VMH in regulating energy homeostasis and body weight [6,7,25,26], we investigated whether the absence of p110α and p110β affected the structural integrity and function of the VMH in p110αβ KOSF1 mice. Nissl staining revealed no noticeable differences between p110αβ KOSF1 and WT mice in the number of neurons or the histological morphology of the VMH structure, nor in other brain regions such as the dorsomedial hypothalamus or arcuate nucleus (Fig. 2A-E). Furthermore, immunohistochemical analysis of SF1 expression [27-29] within the VMH indicated no significant differences in the number of SF1-positive cells between the p110αβ KOSF1 and WT groups (Fig. 2F-H). These findings suggest that the loss of p110α and p110β does not impact the structural development of the VMH or SF1 protein expression in this area.
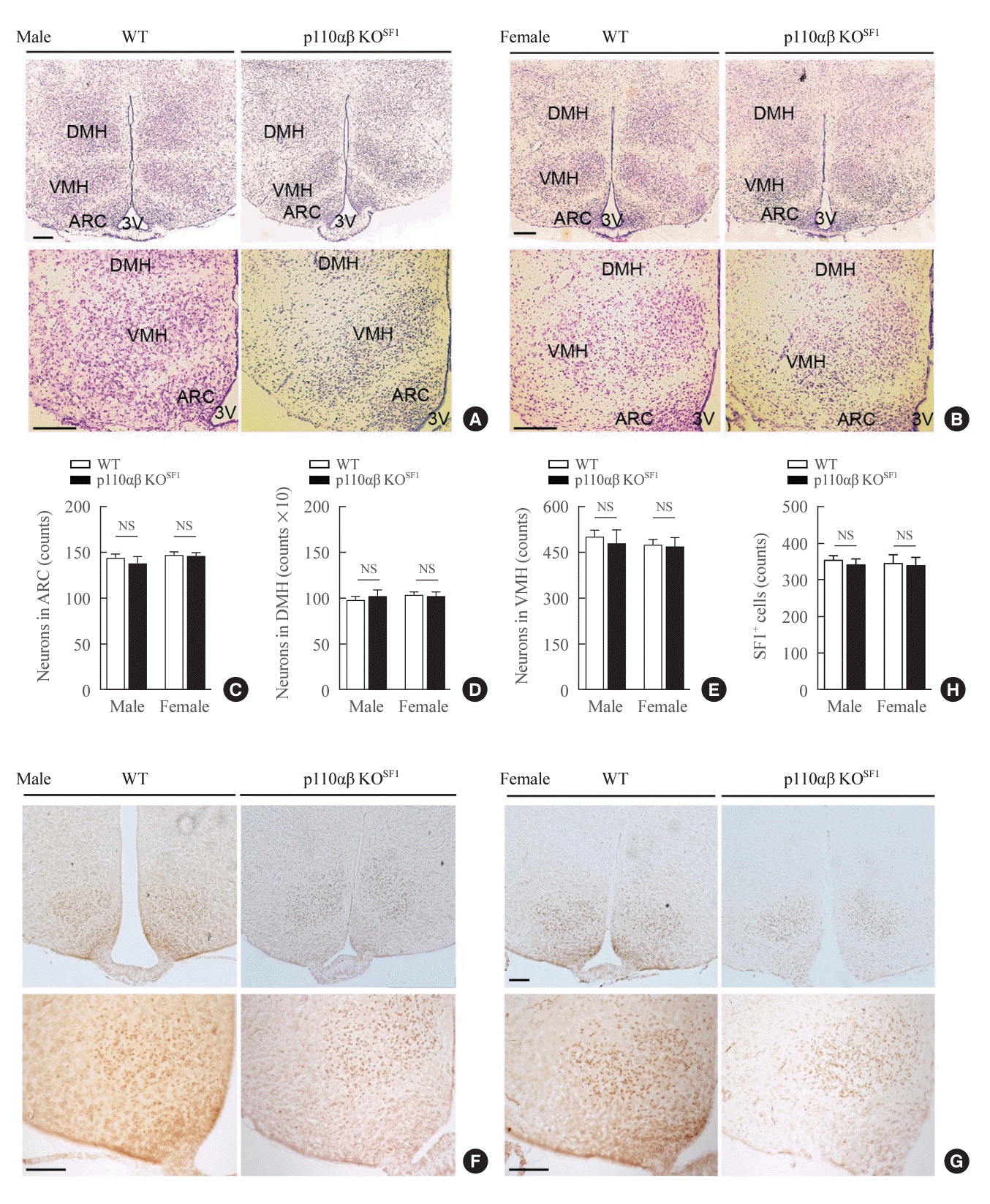 | Fig. 2.Conditional deletion of p110α and p110β in steroidogenic factor 1 (SF1) neurons does not significantly impact the formation of the ventromedial hypothalamus (VMH) or the expression of SF1. (A, B) Representative images of Nissl staining in the hypothalamus region of (A) male (n=2 for wild-type [WT] and n=1 for p110αβ knockout [KO]SF1) and (B) female (n=2 for each group) mice at 12 weeks of age. (C-E) Quantitative data indicating the numbers of neurons in the (C) arcuate nucleus (ARC), (D) dorsomedial hypothalamus (DMH), and (E) ventromedial hypothalamus (VMH). (F, G) Representative SF1 immunohistochemical staining in the brains of (F) male (n=2 for WT and n=1 for p110αβ KOSF1) and (G) female (n=1 for WT and n=2 for p110αβ KOSF1) mice. (H) Quantitative data indicating the numbers of SF1-positive cells in the VMH. The original magnification is ×100 (upper panel) and ×200 (lower panel). Scale bars represent 200 µm. Data are presented as mean±standard error of the mean. 3V, third ventricle; NS, not significant.
|
Altered adrenal cortex development and hormone production in p110αβ KOSF1 mice
SF1 is a key factor in the function, formation, and regulation of the adrenal cortex [30]. In response to the altered glucose metabolism in p110αβ KOSF1 mice (Fig. 1) and the absence of changes in VMH development, we proceeded to examine the structure and function of the adrenal gland. Surprisingly, both male and female p110αβ KOSF1 mice exhibited severe atrophy of these glands (Fig. 3A). The adrenal glands of female animals were harvested and stained with H&E. We observed a dramatic reduction in the size of the adrenal cortex, with the adrenal gland mass consisting primarily of the adrenal medulla (Fig. 3B). To further investigate this phenomenon, we performed immunofluorescence and immunohistochemical analyses using anti-TH (as TH is selectively expressed in the adrenal medulla [31,32]) and anti-SF1 assays. The staining results corroborated the observations from the H&E staining, with SF1-expressing cells forming a thin layer around the normally developed adrenal medulla (Fig. 3C, D). Since the adrenal cortex is crucial for the synthesis of steroidogenic hormones, the abnormal development of this structure in p110αβ KOSF1 mice prompted us to assess the production of adrenal hormones such as aldosterone and corticosterone. As anticipated, both male and female p110αβ KOSF1 mice exhibited significantly lower aldosterone levels compared to WT animals (Fig. 3E, F). Similarly, under basal conditions, corticosterone levels were markedly reduced in both male and female p110αβ KOSF1 mice (Fig. 3G). Specifically, aldosterone levels in p110αβ KOSF1 mice were reduced 3-fold in male and 1.8-fold in female animals, while corticosterone levels were reduced 5.7-fold in males and 13.5-fold in females. Additionally, the male p110αβ KOSF1 mice exhibited impaired responses to stress conditions (Fig. 3H). Overall, the deletion of p110α and p110β in SF1-expressing cells within the adrenal cortex resulted in impaired development and function of the adrenal cortex, without noticeably affecting the development of the adrenal medulla.
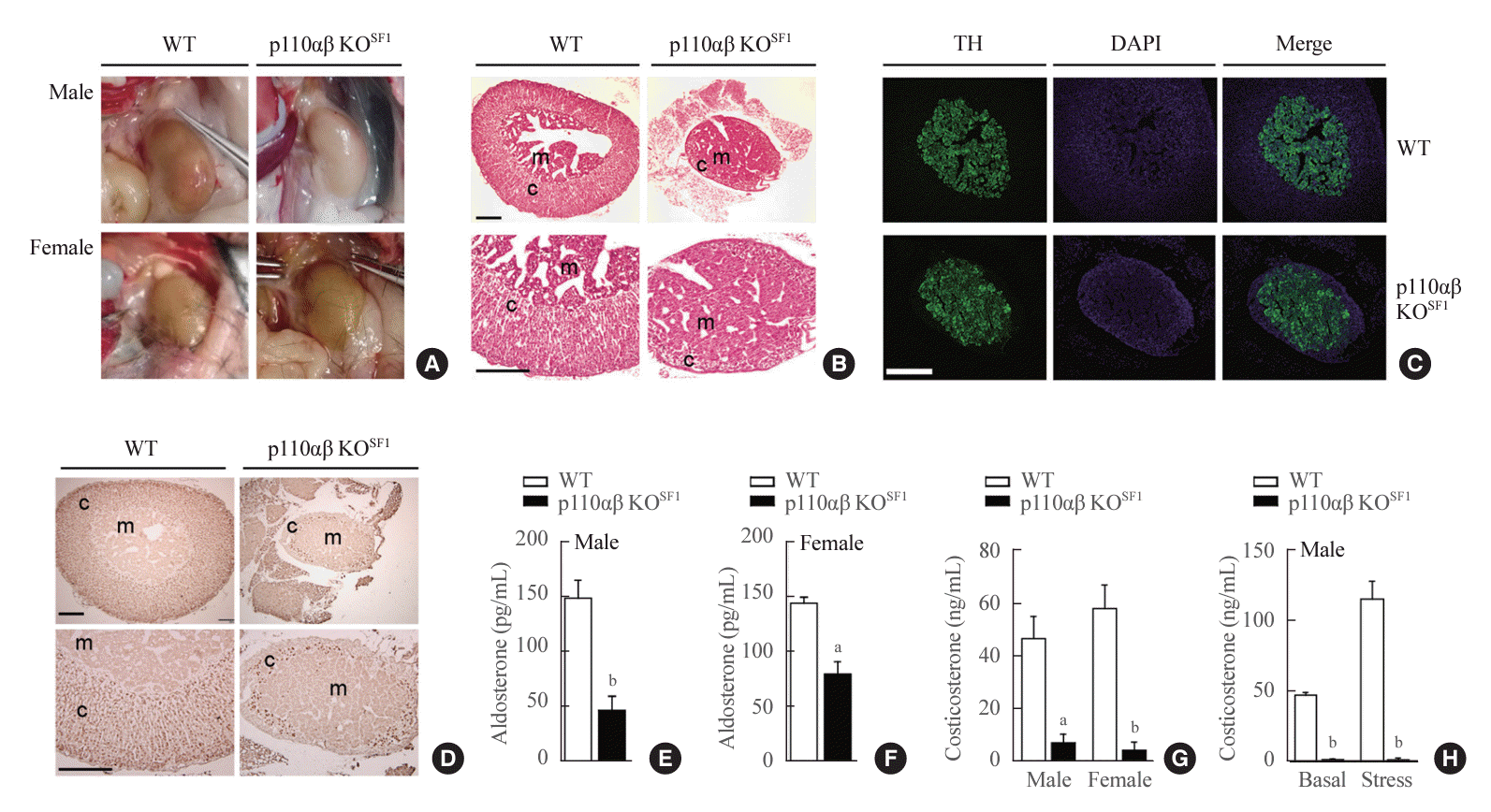 | Fig. 3.Ablation of p110α and p110β in cortical layers leads to impaired development of the adrenal cortex. (A) Representative image of adrenal glands dissected from wild-type (WT) and p110αβ knockout (KO)SF1 mice, with male specimens in the upper panel and female specimens in the lower panel. (B) Representative image of hematoxylin and eosin-stained adrenal glands from female mice. The original magnification is ×100 (upper panel) and ×200 (lower panel). Scale bars represent 200 µm. (C) Representative image of tyrosine hydroxylase (TH) immunofluorescent staining in the adrenal medulla of female mice. Original magnification, ×100. Scale bars represent 250 µm. (D) Representative image of steroidogenic factor 1 (SF1) immunohistochemical staining in the adrenal cortex of female mice. The original magnifications are ×100 (upper panel) and ×200 (lower panel). Scale bars represent 200 µm. (E, F) Aldosterone levels in 12-week-old (E) male (n=7 for each group) and (F) female (n=3 for the WT group and n=8 for the p110αβ KOSF1 group) mice. (G) Corticosterone levels in 8-week-old male (n=4 for the WT group and n=6 for the p110αβ KOSF1 group) and female (n=4 for the WT group and n=5 for the p110αβ KOSF1 group) mice under basal conditions. (H) Corticosterone levels in 12-week-old male mice (n=3 for the WT group and n=4 for the p110αβ KOSF1 group) under basal and stress conditions. Data are presented as mean±standard error of the mean. DAPI, 4´,6-diamidino2-phenylindole; c, adrenal cortex; m, adrenal medulla. aP<0.01 and bP<0.001 vs. WT group.
|
Impaired reproductive organ development in p110αβ KOSF1 mice
In addition to the VMH and adrenal glands, the gonads express high levels of SF1 [33,34]. Consequently, we investigated the formation of testicular and ovarian tissues. Notably, the testicular size of p110αβ KOSF1 mice was substantially reduced compared with that of WT mice (Fig. 4A), as evidenced by a decrease in the vertical testicular dimension (Fig. 4B). Furthermore, the testicular weight of the p110αβ KOSF1 mice was only around half that of WT mice (Fig. 4C). Similarly, the ovaries of female p110αβ KOSF1 mice displayed clearly impaired development relative to those of WT female mice, including smaller size (Fig. 4D). Additionally, we noted a significant reduction in the number of follicles at each stage of follicular development in female p110αβ KOSF1 mice, although the size of the follicles remained unchanged (Fig. 4D). These data indicate the critical roles of p110α and p110β within SF1-expressing cells in the development of the testes and ovaries. In summary, these results underscore the integral nature of p110α and p110β in the regulation of key physiological processes, including survival, metabolic homeostasis, adrenal gland function, and reproductive organ development, through their activity in SF1-expressing cells.
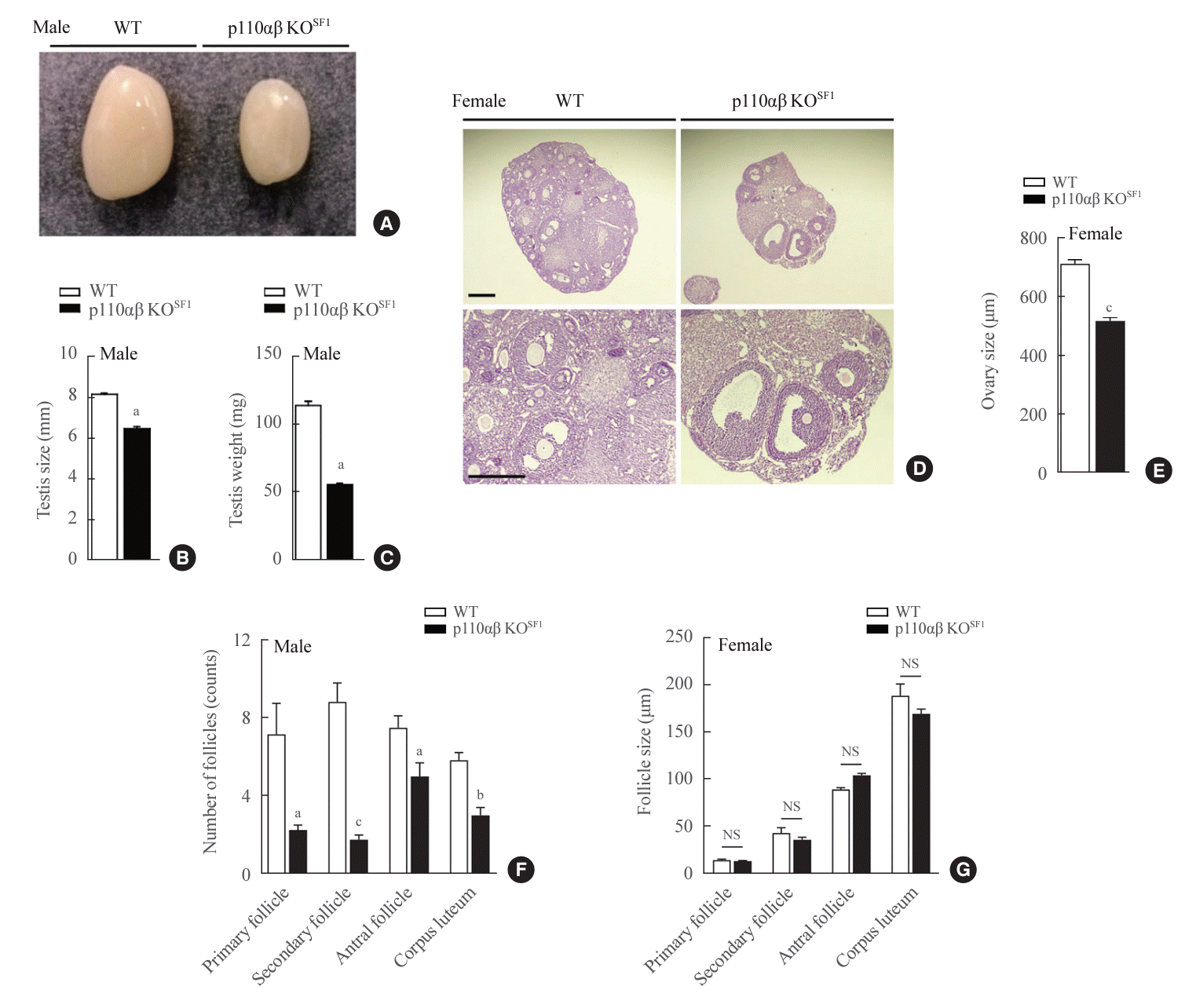 | Fig. 4.Absence of p110α and p110β in steroidogenic factor 1 (SF1)-expressing cells leads to severe testicular and ovarian hypotrophy. (A) Representative image of testes dissected from wild-type (WT) and p110αβ knockout (KO)SF1 mice. (B) Average size and (C) weight of testes from WT (n=5) and p110αβ KOSF1 (n=3) male mice. (D) Representative image of hematoxylin and eosin-stained ovaries. The original magnification is ×100 (upper panel) and ×200 (lower panel). Scale bars represent 200 µm (upper panel) and 100 µm (lower panel). (E) The size of ovaries from WT (n=2) and p110αβ KOSF1 (n=1) female mice. (F) The number and (G) size of follicles in WT (n=2) and p110αβ KOSF1 (n=1) female mice. Data are presented as mean±standard error of the mean. NS, not significant. aP<0.05, bP<0.001, and cP<0.0001 vs. WT group.
|
Go to :
DISCUSSION
PI3K and its subunits play fundamental roles in energy homeostasis and gonadal development [5,35]. In SF1 neurons within the VMH, the deletion of insulin receptors inhibits PI3K activity in the context of obesity triggered by a high-fat diet [36,37]. Furthermore, the ablation of the catalytic subunit p110α in SF1 neurons results in hypersensitivity to obesity induced by such a diet [7]. Notably, knockdown of p110α can lead to a compensatory rise in other p110 isoforms, particularly p110β [38]. In this report, we demonstrate that the loss of both PI3K catalytic subunits (p110α and p110β) in SF1-expressing tissues leads to changes in whole-body metabolism, particularly glucose metabolism and adrenal steroidogenic hormone production, and is associated with impaired development of the adrenal cortex and gonads.
The conditional KO of p110α and p110β was linked to a higher mortality rate in both male and female p110αβ KOSF1 mice (Fig. 1A, B). Consistent with these findings, PI3K has been identified as a target for cancer therapy. Previous research has demonstrated that inhibition of PI3K increases gemcitabine-induced apoptosis [39] and that the knockdown of p110α and p110β leads to reduced cell proliferation and increased apoptosis [40]. However, in our study, p110αβ KOSF1 male mice exhibited a decrease in body weight gain after 12 weeks, along with improved postprandial glucose levels and impaired glucose tolerance (Fig. 1C-G). These effects were not seen in mice with a single KO of the p110α subunit maintained on a chow diet [7]. Considering the changes in energy homeostasis, we examined the structure of the VMH, but we found no significant changes in VMH development or SF1 protein levels (Fig. 2).
Consequently, we investigated another critical endocrine organ: the adrenal gland. SF1 is expressed in the adrenal cortex from the 13.5-day post-coitum fetus [41] and is essential for the development and function of this organ. The ablation of p110α and p110β in SF1-expressing cells within the cortical layers led to the impaired formation of these layers, but not the adrenal medulla (Fig. 3A-D). Indeed, a previous study demonstrated that a specific KO of Sonic hedgehog in SF1-expressing cells reduced the thickness of the adrenal cortex by 50% to 70% [42]. However, the loss of p110α and p110β resulted in a notably thin adrenal cortex, as depicted in Fig. 3B-D. This thinning may explain surgical challenges that we encountered in the p110αβ KOSF1 mice, both male and female. Upon observing the compromised development of the adrenal cortex, we measured steroidogenic hormones to evaluate adrenal function. As anticipated, we observed a marked reduction in the production of aldosterone and corticosterone, even under stress conditions (Fig. 3E-H). To further investigate the impacts of p110α and p110β on various cell types in the adrenal cortex, detailed experiments are required to distinguish the remaining cells. However, the severe atrophy of the adrenal cortex may pose challenges to such discrimination.
Prior to the differentiation of the adrenal cortex and gonads, these tissues originate from the same source, known as the adrenogonadal primordium [43]. The development of this primordium is dependent on the expression of SF1 [44]. Consequently, we explored gonadal development. In the testes, SF1 is present in both Leydig (interstitial) and Sertoli (cord) cells [41,45]. In the ovaries, SF1 is expressed in theca and granulosa cells [45-47]. Due to the extensive expression of SF1 in the gonads, the loss of p110α and p110β in SF1-expressing cells resulted in severe testicular and ovarian atrophy in p110αβ KOSF1 male and female mice, respectively (Fig. 4). A previous study demonstrated that whole-body mutation of p110β leads to reduced testicular size, fertility, and fecundity in mutant male mice, while female mice exhibited no significant changes [14].
The present findings underscore the potential developmental role of PI3K in organs expressing SF1. PI3K and its subunits are not only pivotal in cell differentiation, proliferation, and energy metabolism but also play a critical role in the formation of the adrenal cortex and gonads [25]. In the absence of the catalytic subunits p110α and p110β, the adrenal cortex does not form properly, leading to reduced production of steroidogenic hormones. This deficiency is a key factor contributing to the lower survival rates observed in p110αβ KOSF1 mice. These findings suggest that PI3K signaling in cells expressing SF1 may exert differential effects on the development of the VMH, adrenal cortex, testis, and ovary. Such insights provide a direction for future studies to elucidate the molecular mechanisms underlying these distinct effects.
Go to :
 XML Download
XML Download