

 PDF
PDF Citation
Citation Print
Print



Highlights
· This review highlights the gut microbiome’s potential in predicting glycemic response.
· The gut microbiome affects glucose homeostasis through its metabolites.
· T2D-specific microbiome findings call for further studies on its function.
· The drug-microbiome axis should be considered when studying microbiome effects in T2D.
· Microbiome big data and multi-omics offer AI-driven insights for T2D modulation.
INTRODUCTION
Presently, more than half a billion peoples are living with diabetes worldwide, and the number is expected to rise to nearly 800 million by 2045, with the majority having type 2 diabetes mellitus (T2DM) [1]. T2DM is a multifactorial metabolic disorder characterized by dysregulated glucose homeostasis, impaired insulin secretion, and insulin resistance. Inadequate glucose management in people with T2DM can lead to complications affecting both small and large blood vessels, including retinopathy, neuropathy, nephropathy, and heart diseases [2]. The development of T2DM is influenced by risk factors such as poor dietary habits, lifestyle choices, and genetic factors, which together complicate disease prevention and treatment [3].
Furthermore, advancements in analytical techniques have made it possible for metabolomics to identify and quantify multiple biomarkers simultaneously in a high-throughput manner, offering valuable insights for managing this complex disease [4]. In the last few years, alongside genetic research, metabolomics has emerged as one of the most widely used and effective methods for diagnosing and predicting T2DM. Metabolomics has also been used to investigate the possible pathways affected by medications [4]. The alterations observed in metabolite profiles underscore their role in the pathogenesis of T2DM, emphasizing the need for exploring novel therapeutic targets (Table 1) [5-16].
Similarly, proteomic studies have identified a range of proteins associated with diabetes. However, the comparison of these findings is complicated by inconsistencies in classification. Integrating proteins into interaction networks can provide a clearer perspective. Despite the extensive data, only a limited number of discoveries have been thoroughly validated [17]. These proteins and their interactions are summarized in Table 2 [18-34]. Furthermore, these findings highlight the complexity of diabetes and the need for integrated omics approaches to uncover the underlying mechanisms and potential biomarkers for diagnosis and treatment.
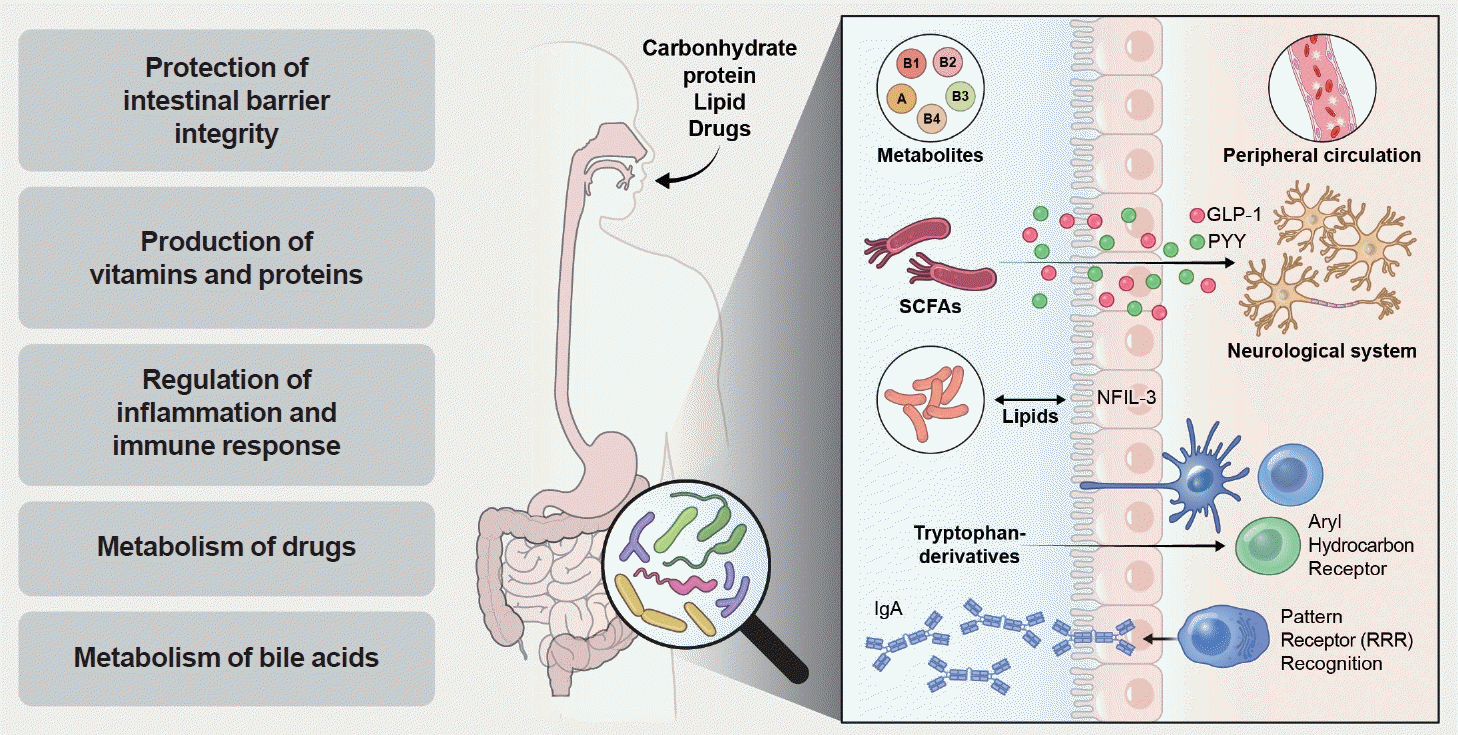
Given the complexity of T2DM pathogenesis and the insights gained from studying metabolites and proteins associated with the disease, recent research has increasingly focused on the role of the gut microbiome in regulating host metabolism and glucose homeostasis [35]. Moreover, since metabolic syndrome is a critical risk factor for T2DM, the gut microbiome has been extensively linked with the metabolic syndrome and T2DM in humans [35]. After the birth, all individuals harbour specific and stable gut microbiome that supports host functions. For example, right after the birth, commensal microbes colonize in the human gut to support the digestion of various nutrients that host cannot break down, such as resistant fibers. In the meantime, gut microbiota supports the development and function of many organs and tissues, including immune system and brain as well (e.g., neurogenesis, microglial maturation, and myelination) [36]. In addition, microbiome takes part in the regulation of metabolic homeostasis, by actively engaged in the control over the whole-body metabolism, including the regulation of gut hormone secretions. Some reports identified that yo-yo effects after weight reductions can be improved by modulating human gut microbiome [37], thereby being a good agent to modulate host metabolic phenotype (Fig. 1) [38,39]. Therefore, understanding of gut microbiome can be a key to prevent and treat many different metabolic diseases.
Molecular functions of human gut microbiome are encoded in their genetic repertoire and has been known to outnumber human (approximately 100 times more). However, identification of individual microbial genomes has been challenged due to the limited culture conditions of most commensal microbes. Interestingly, recent advances in next-generation sequencing has helped us to investigate the full genetic repertoire of human gut microbiome without cultivation. For example, de novo assembly of gut metagenome identified individual bacterial genomes by in silico approaches, called metagenome-assembled genomes (MAGs), even up to 92,143 MAGs in our gut microbiome known so far [40,41]. Such analyses also identified that human gut microbiome provided numerous carbohydrate-processing enzymes [42], and produce antibiotics-like substances for the protection of host to pathogen invasions [43].
To better characterize the composition and function of human gut microbiome, large collective efforts has been put into the construction of microbiome big data generation. For example, at the first phase of human microbiome project (HMP), the unique composition of human microbiome in all the body part has been explored [44]. At the second phase of HMP, iHMP project, by combining host multiomics data with gut microbiome meta-omics, functional roles of human microbiome has been extensively studied [45].
In line with large-scale microbiome big data analysis, many personalized nutrient projects with diverse microbiome data generation has been established [46]. For example, individual postprandial glucose responses of 800 normal subjects recorded with their diets, clinical parameters, and microbiome compositions as well [47]. In this project, microbiome information has been used as a proxy of predicting glucose responses and also guided the design of personalized diet to reduce blood glucose levels effectively [47,48].
The emerging role of gut microbiota in diabetes management is shown by its potential in diagnosing and predicting the risk of T2DM. Understanding individual microbiomes through big data analysis will offer a promising new avenue for developing novel preventive strategies for various chronic metabolic diseases, such as obesity, diabetes, and fatty liver diseases. A specific metagenomic linkage group associated with an increased risk of T2DM has been identified, emphasizing the connection between microbial profiles and the disease [49]. Key microbial risk markers include altered microbial diversity, reduced populations of butyrate-producing bacteria, elevated levels of opportunistic pathogens, and increased microbial functions such as sulphate resistance and oxidative stress [11].
This review summarizes the recent advances in understanding the gut microbiome’s role in metabolic phenotypes and glycaemic control. After discussing the roles of metabolites and proteins in T2DM, we focus on how the gut microbiome influences metabolism and glucose homeostasis. We also explore the use of artificial intelligence (AI) and machine learning to predict physiological changes and improve early diagnosis and prevention of metabolic diseases.
GUT MICROBIOME AS A MODULATOR OF HOST ENERGY STATUS AND GLYCAEMIC RESPONSES
Gut microbiome plays a key role in host energy balance. Roughly for the host, through the digestion of many different types of dietary polysaccharides [50,51]. In previous study, germ-free mice has lower “energy harvest” from the diet, compared to conventionally-raised mice due to the lack of the microbiome [52], and re-colonization of microbiome from obese mice resulted in significant increase of total body fat [53]. Likewise, transferring faecal microbiome from human obese twins significantly increased the adiposity of mouse with faecal microbiota transplanted [54], thereby confirming that gut microbiome as a key modulator of host energy balances. Furthermore, population studies based on human genetics and gut microbiome showed that many metabolic phenotypes were actually explained by microbiome factors, rather than host genetic factors, such as body mass index, glycaemic status, and cholesterol levels [55]. Indeed, based on large-scale plasma metabolomics studies, together with human microbiome analyses, human metabolic make-up (i.e., metabolome) can be explained by gut microbiome, up to 46% of their variations [56], and outperformed than genetics and diet factors [57]. Therefore, understanding the role of human gut microbiome is a key to understand host energy metabolism changes.
Usually, gut microbiome generated fermentation products as an energy input, including short-chain fatty acids (SCFAs) [58,59], and such microbial metabolites serve as a major energy source of host cells, providing 60% to 70% of their energy supply of colonocytes (Fig. 1) [60]. SCFAs can also trigger G protein coupled receptor (GPCR) signaling by binding to GPR41, GPR43, and GPR109A [61-65], affecting the wide range of tissue physiology, including intestine, kidney and nervous system. Notably, SCFAs can act on GPCRs expressed by enteroendocrine cells, stimulating the secretion of glucagon-like peptide 1 (GLP-1), peptide YY (PYY), and ghrelin release, and subsequently can increase the insulin biosynthesis, fat storage and regulate appetite and satiety [66]. Additionally, SCFAs exerted their effects through multiple mechanisms, including activation of free fatty acid receptors 2 and 3 (FFAR2/3). These receptors, which were identified as novel therapeutic targets [67], were widely expressed in various cell types in both humans and mice, serving as sensors for SCFAs and regulating a range of physiological and cellular processes [67].
To confirm the effects of SCFAs on GLP-1 secretion and glucose intolerance, some studies generated ffar2 and ffar3-deficient mice and observed the decreasing trends of GLP-1 section and glucose intolerance at the in vitro and in vivo levels [67], confirming its regulatory roles on host energy balances and glycaemic status. However, there are some debates about effects of SCFAs to the metabolic phenotypes, e.g., acetate; thus, its effects should be carefully studied in different metabolic context.
Other energy metabolites from gut microbial fermentation, such as succinate, formate, and lactate, can serve as key biosynthetic precursors of whole-body metabolism and its excessive amounts can lead to metabolic dysregulations. For example, succinate can serve as a precursor metabolite for SCFA production and intestinal gluconeogenesis, thereby improving glucose homeostasis. In addition, succinate can be taken up by brown adipose tissues and promoted thermogenesis to improve metabolic phenotype [68]. However, excessive amounts of succinate can also promote the colonization of pathobionts and induce inflammatory conditions, by polarizing M2 macrophages and activating other immune cells. Likewise, formate itself can be used for the nucleotide synthesis, but its excessive amounts can promote intestinal inflammation when it is oxidized [69] and also drives colorectal cancer progression by increasing cancer stemness [70]. Significantly, formate was identified as a potential biomarker for diabetic peripheral neuropathy, with lower levels linked to mitochondrial dysfunction and changes in gut microbiota. This indicated its role in regulating blood sugar and metabolic processes [71].
Microbially-produced lactate (e.g., Streptococci and Lactobacilli), a minor metabolite in the healthy colon, can be consumed by other lactate-utilizing bacteria to produce SCFAs and help epithelial development, but its accumulations can promote intestinal inflammation [72,73]. Therefore, fostering metabolic balances between host and microbiome can be a key to regulate whole-body metabolic homeostasis.
In summary, the gut microbiome plays a significant role in regulating the energy balance and glycaemic control of the host by interacting with metabolic pathways and producing essential metabolites (Table 3).
THE ROLE OF MICROBIOME IN THE PATHOGENESIS OF TYPE 2 DIABETES MELLITUS
T2DM is a complex metabolic disorder characterized by dysregulated glucose metabolism, which arises from a multifactorial interplay of genetic, environmental, and behavioural elements [74,75]. There was a strong evidence demonstrating the influence of microbiota on glucose metabolism in both preclinical animal models of T2DM and healthy animals. As a result, there has been a considerable focus on the clinical significance of the microbiome in order to enhance our understanding and address the challenges associated with T2DM. Various studies reported the associations between different taxa and T2DM. In individuals diagnosed with T2DM, a decreased in beneficial butyrate-producing strains such as Faecalibacterium prausnitzii and Roseburia intestinalis, along with an increased in opportunistic pathogens like Escherichia coli and Clostridium ramosum was observed [74,76]. Indeed, in randomized clinical study of T2DM patients, ones supplemented with isoenergetic high fiber diets showed improved glucose homeostasis by selectively promoting SCFAs producers, such as F. prausnitzii and Lachnospiraceae spp. [77]. Broadly, certain taxa at the genus level, such as Bacteroides and Bifidobacterium, consistently demonstrated a protective association with T2DM, while Actinobacteria showed a similar pattern at higher taxonomic levels [78]. On the other hand, taxa such as Lactobacilli exhibited effects that were specific to certain species or strains. This phenomenon could be attributed to the diversity within a particular genus residing in the human gut, where a greater number of strains of a genus led to the observation of more strain-specific effects [79]. Moreover, a recent comprehensive study revealed geographical variations in the microbial composition associated with similar metabolic outcomes [80]. Several studies have highlighted notable differences in the microbiota composition between individuals with and without T2DM [79,81].
Based on many mechanistic and observational studies, it was proposed that the role of microbiota in the onset and progression of T2DM was mediated through its metabolites. Once absorbed by the host, these metabolites were processed by endogenous enzymes and interacted with specific receptors, affecting various physiological functions. Table 3 summarized how these metabolites influenced different organs and contributed to the onset and progression of metabolic syndrome and T2DM [35,82-114].
Based on these observations, it was proposed that the involvement of the microbiota in T2DM was due to (1) decreased intestinal barrier integrity, (2) reduced production of SCFAs, (3) altered bile acid metabolism, and (4) increased production of branched-chain amino acids (BCAAs) [74,115]. For example, some gut bacteria, such as Bacteroides vulgatus and Prevotella copri can biosynthesize BCAAs, augmenting circulating levels of human plasma BCAAs [116]. It is also known that metabolic endotoxaemia caused by decreased barrier integrity and the production of metabolic endotoxins, such as lipopolysaccharide [80] and imidazole propionate [117] can promote T2DM pathogenesis. Interestingly, recent studies identified that Akkermansia muciniphila can play a protective role in increasing barrier integrity and inhibiting glucosidase activity [79], and also its secreted proteins found to improve glucose homeostasis [118,119]. Therefore, modulating gut microbial community toward beneficial community can be a key to treat T2DM pathogenesis. In addition to known microbial functions on T2DM pathogenesis, there are many alternative hypotheses that gut microbiomes can regulate glucose metabolism by other non-canonical metabolites. For example, peripheral serotonin, mediated by the gut microbiome, can influence glucose homeostasis [120], pharmacological and genetic inhibitions of gut-derived serotonin induced host glucose handling. Other tryptophan catabolites, such as indole-3-propionic acid (IPA) and tryptamine, also found to play a beneficial effect on diabetes, significantly reducing fasting blood glucose and insulin levels in mouse experiments [121]. Therefore, identification of beneficial microbial metabolites can be new avenue of improving glucose metabolism in diabetic patients.
Nevertheless, in order to make the most of these observations, it is crucial to take into account the impact of different factors, such as interactions between drugs and the microbiome, on these results. In the next part, ‘Hidden Factors of Gut Microbiome Signatures—Drug-Microbiome Interactions,’ we will explore the influence of drugs on the microbiome and how these interactions can impact the effectiveness of diabetes treatments. Knowledge of these dynamics is essential for improving our strategy in handling T2DM.
HIDDEN FACTORS OF GUT MICROBIOME SIGNATURES—DRUG-MICROBIOME INTERACTIONS
Multiple molecular mechanisms of gut microbiota contribution to metabolic disease and T2DM were recently reviewed elsewhere [122]. The microbiota actively influenced inflammation, interacts with dietary components, impacts gut permeability, regulates glucose and lipid metabolism, modulates insulin sensitivity, and maintains overall energy homeostasis in the mammalian host [79]. In recent years, gut microbiome found to be modulated by host-direct drugs, metformin being a key example of such drug-microbiome interactions [123-125]. Indeed, in a large-scale association study, among single-drug formulations, metformin were found to have a largest number of associations with microbiome composition and pathways, together with proton pump inhibitors and laxatives [126].
The gut microbiome had played a vital role in drug and xenobiotic metabolism, thereby impacting the effectiveness and response of antidiabetic medications [127]. Many orally administered drugs and xenobiotics were processed in the intestines by microbial enzymes before being absorbed into the bloodstream. As a result, the metabolic capacity of the gut microbiome influenced the absorption, activity, and toxicity of these drugs, rendering them pharmacologically active, inactive, or even toxic, thereby affecting the metabolic functions of the host (Fig. 1). This phenomenon is known as the drug-microbiome-metabolism axis [127].
Some study also suggested that health benefits of metformin could be attributed to the specific changes it induced in the gut microbiota. Human studies were consistently demonstrated that metformin administration leads to increased abundance of E. coli and decreased levels of Intestinibacter bartlettii [124, 126,128]. Single-arm studies and a short-term trial provided evidence indicating that metformin exerts an influence on the composition of gut microbiota [128-130]. Similarly, individuals with T2DM exhibit changes in their gut microbiota, characterized by decreased levels of butyrate-producing genera such as Roseburia, Subdoligranulum, and Clostridiales. Interestingly, the administration of metformin has been shown to reduce or even reverse these alterations in the population of Firmicutes, providing potential therapeutic benefits [124]. Experimental findings in mice provided evidence that the modulation of gut microbiota, specifically through an increase in the population of Akkermansia spp., played a role in the antidiabetic effects of metformin. This finding introduced a novel mechanism by which metformin exerted its therapeutic impact in individuals with T2DM [131]. It was concluded that antidiabetic drugs not only affect the gut microbiome but are also reciprocally/bidirectionally influenced by it. Several additional antidiabetic drugs are summarized in the table, all impacting the composition of the microbiota (Table 4) [14,124,130,132-143]. Understanding and predicting these interactions in T2DM required the use of advanced methods, such as machine learning and AI, due to their complexity.
USING MACHINE LEARNING TO PREDICT METABOLIC STATUS AND GLYCAEMIC RESPONSES, WITH INSIGHTS FROM THE GUT MICROBIOME
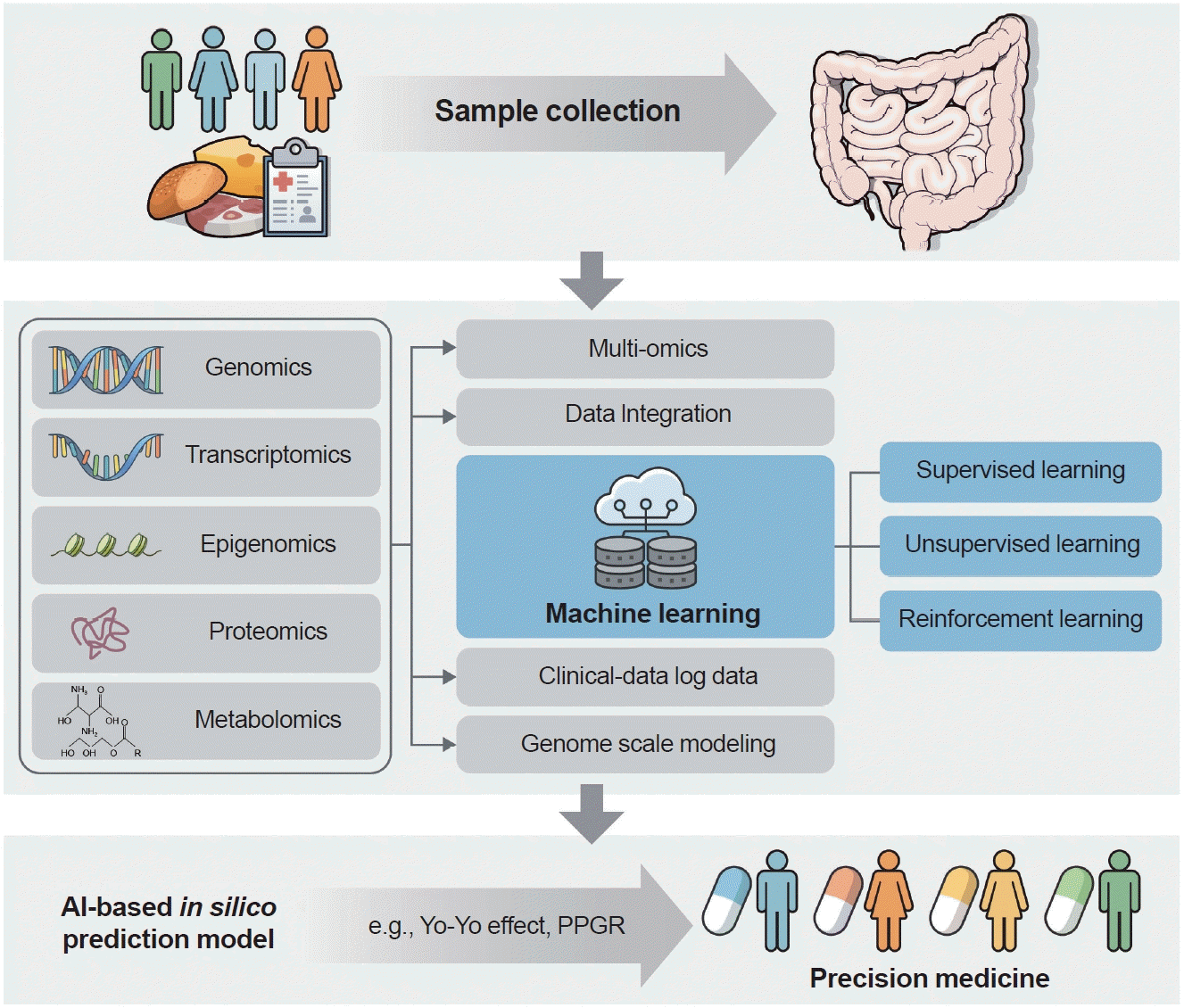
The advancements in multiomics research on gut microbiota have paved the way for personalized microbiota-based treatments in patients with T2DM. Integrating host multiomics data with microbiome information holds the potential to enable the development of tailored therapeutic approaches for T2DM (Fig. 2). In recent years, microbiome big data with deep phenotyping enabled the in silico prediction of host physiology, including the prediction of postprandial glycaemic responses (PPGRs), weight-regain (i.e., yo-yo) effect after diet challenges, and microbiome aging clock. A cornerstone study for AI-based in silico prediction model is glycaemic response prediction model by Eran Segal group [47]. In this study, authors found that glycaemic responses can be predicted based on individual microbiome and phenotype information and this helped to guide personalized nutrition. For examples, prediction models rank the meals that are associated with low PPGRs or high PPGRs. In prediction arm, healthy participants followed the “good” diet for meals of low PPGRs or “bad” diet for meals of high PPGRs for 1-week and in expert arm, they followed the diets recommended by clinical dietitians. Interestingly, PPGRs founds to be lower for ones followed “good” diets from the prediction model, compared to those from the clinical dietitians. In follow-up study of diabetic subjects, similar prediction models recommended “good” and “bad” diets, and those who followed “good” diets had lower PPGRs, compared to ones followed Mediterranean diets.
In other study from deep phenotyped subjects with diet and microbiome information (The Personalized Nutrition Project [PNP] and Personalized Responses to Dietary Composition Trial-1 [PREDICT-1] study) [144], authors identified gut microbial species associated with specific diets that we are normally eating, and also associated with favourable outcomes of cardio-metabolic and postprandial markers. Among the 30 microbial species found to predict favourable health outcomes, specific microbes like P. copri [5] and Blastocystis spp. [145] have been observed to correlate with enhanced postprandial glucose metabolism, while others are indicative of lipid and peptide fluctuations post-meal [144]. These finding suggested a new paradigm of designing diets based on their own microbiome make-ups.
In addition, some study identified that metabolic phenotype can be stored as “microbiome memory” and such memory can be used for developing machine learning models to predict weight regains after diet challenges, called yo-yo effects [146], which can be also linked to the role of microbiome in host glycaemic control, influencing satiety signaling and energy metabolism. Interestingly, authors of this study performed memory wipe-out by antibiotics treatments and such yo-yo effects were significantly reduced, thereby implicating that such information can be a good proxy to design personal metabolic phenotypes. Therefore, developing accurate individual microbiome-based prediction models will be a good platform to guide and evaluate microbiome-based therapeutics and also dietary recommendations to tailored to each person [147], shaping better health benefits in near future.
CONCLUSIONS
This review explored the complex relationships among T2DM, metabolites, proteins, and the gut microbiome, with a focus on glycaemic responses. The integration of multiomics, precision medicine, machine learning, and personalized dietetics provides a revolutionary approach to healthcare, particularly for complicated illnesses such as diabetes. Researchers have obtained substantial insights into the complex relationships between the gut microbiome, metabolic health, and dietary factors by utilizing cutting-edge technology and interdisciplinary collaboration. This comprehensive approach enables the development of personalized therapies based on an individual’s genetic, physiological, and dietary profiles, as well as personalized medications. The relationship between microbiota and T2DM is due to weak intestinal barrier integrity, decreased SCFA production, altered bile acid metabolism, and increased BCAA production. Customized diabetes therapy is critical, especially considering the potential for metformin and other diabetic medicines to exacerbate dysbiosis. Our recommendation emphasizes the need of integrating personalized dietary suggestions with precise medicine administration to improve glucose management, restore gut microbial balance, boost SCFA production, maintain bile acid levels, and protect intestinal barrier integrity. These interventions can effectively slow disease progression and improve overall health outcomes for individuals with diabetes.
Despite limitations such as a lack of user-friendly reporting for clinicians in multiomics research, accomplishing the full potential of this approach will require continued research, innovation, and collaboration across healthcare disciplines. Embracing this comprehensive approach has the potential to revolutionize diabetes management and bring in a new era of personalized healthcare that emphasizes individualized nutrition and optimal health outcomes.
 XML Download
XML Download