

 PDF
PDF Citation
Citation Print
Print



Introduction
Gastric cancer is the sixth most common type of malignancy and the third leading cause of cancer-related mortality globally [1]. Gastric cancer, a complex and heterogeneous malignancy, remains a significant global health challenge with diverse molecular subtypes that impact prognosis and treatment outcomes [2]. In 2014, Cancer Genome Atlas Research Network proposed a molecular classification dividing gastric cancer into four subtypes [3]. One of the subtypes that has gained substantial attention in recent years is microsatellite instability–high (MSI-H) subtype with underlying genomic instability, particularly in regions of repetitive DNA sequences known as microsatellites.
MSI-H gastric cancers, characterized by a high frequency of genetic alterations in microsatellites, exhibit distinct clinical and immunological features compared to their microsatellite stable (MSS) counterparts. In comparison with MSS gastric cancer, patients with MSI-H gastric cancer demonstrated a significantly higher survival rate but did not benefit from neoadjuvant or adjuvant chemotherapy [4,5]. In MSI-H gastric cancer, tumor mutation burdens enhance the expression of tumor-specific neoantigens, increasing the chances of recognition by T cell and the infiltration of T cell in the tumor microenvironment [6]. Alongside the high infiltration of T cell, a high prevalence of programmed death-ligand 1 (PD-L1) positivity has also been reported in MSI-H gastric cancer. Due to this characteristic, MSI-H gastric cancer is expected to respond favorably to immune checkpoint inhibitors (ICIs).
In the recent years, ICIs have emerged as a novel therapy for various tumors. Several studies in gastric cancer have demonstrated the effectiveness of ICIs, such as anti–programmed death-1 (PD-1)/PD-L1, in killing tumor cells through the activation of the immune response within the tumor microenvironment [7,8]. Conversely, some studies did not exhibit superiority over chemotherapy for overall survival in patients with human epidermal growth factor receptor 2–negative and combined positive score of one or more [9]. In the context of MSI-H gastric cancer, a phase II trial of pembrolizumab monotherapy reported a response rate of 85.7% in MSI-H patients, suggesting that MSI-H could serve as a reliable biomarker for immunotherapy, in addition to PD-L1 immunohistochemistry [10]. As shown above, the response rate of ICIs in gastric cancer remains still limited. In-depth research on the interaction of tumor cell and immune cell in the tumor microenvironment is imperative for identifying accurate biomarkers that can predict the response to ICIs.
Recent advancements in single-cell RNA sequencing (scRNA-seq) and spatial transcriptome technology have greatly facilitated the molecular characterization of the transcriptome. These methods enable the understanding of cellular heterogeneity at the molecular level and the profiling of subclusters of tumor-infiltrating lymphocytes [11-13]. Despite these advances, the data generated by current scRNA-seq technologies in gastric cancer remain insufficient to provide comprehensive information regarding the characteristics of the immune microenvironment in MSI-H and MSS gastric cancer.
In this study, we conducted scRNA-seq and spatial transcriptome analysis to investigate the molecular characteristics of the immune microenvironment, with a specific focus on CD8+ T cell, T cell exhaustion, and the interaction between tumor and immune cell based on MSI status of gastric cancer.
Go to : 
Materials and Methods
1. Patients
This study included six patients with gastric adenocarcinoma who were diagnosed through endoscopic biopsy at Seoul National University Hospital from July to August 2021. All patients had advanced gastric cancer, as indicated by preoperative endoscopy and computed tomography scans. Surgical treatment was subsequently undertaken for all patients.
2. Microsatellite instability status
MSI status was assessed at five loci (BAT25, BAT26, D2S123, D5S346, and D17S250) according to the National Institutes of Health guidelines. In brief, tumor and paired normal areas were dissected from formalin-fixed, paraffin-embedded blocks, and genomic DNA was extracted. After polymerase chain reaction (PCR) amplification, the PCR products were run on the ABI 3731 automatic DNA sequencer (Applied Biosystems, Foster City, CA) [14]. GC with instability in two or more, one, or no loci were considered MSI-H, microsatellite instability–low, and MSS, respectively.
3. Single-cell RNA sequencing
Tumor and matched normal tissues were obtained immediately after surgical resection of the stomach to minimize cold ischemic time. Although we collected six tumor samples and paired six normal samples, we selected two normal samples with higher cell viability because of the limitation of budget. The samples were processed for library preparation using Chromium Next GEM Single-Cell 3′ Reagent Kits v3.1 (10× Genomics, Pleasanton, CA). After appropriate cell dissociation and viability assessment (cutoff 80%), cell suspensions were diluted to a targeted count of 10,000. The cells were mixed with master mix and loaded onto a chromium Next GEM chip G with Single Cell 3′ v3.1 Gel Beads and Partitioning Oil. RNA transcripts from single cells were uniquely barcoded and reverse transcribed within droplets. The cDNA molecules were pooled and processed for library construction. The libraries were then quantified using quantitative PCR and qualified using Agilent Technologies 4200 TapeStation (Santa Clara, CA). Finally, the libraries were sequenced using the HiSeq platform (Illumina, San Diego, CA).
4. Quality control, preprocessing of sequencing data, and unsupervised clustering
Raw gene expression matrix was generated for each sample using CellRanger v2.0.2. Cells that had less than 300 expressed genes or more than 6,000 expressed genes were removed. Cells in which the fraction of mitochondrial genes exceeded 20% also were removed. We removed doublet cells using the DoubletFinder package with PCs 1 to 30, pN set to 0.25, and pK set to 0.092. We identified 3,000 highly variable genes using the FindVariableFeatures function in the Seurat library for initial clustering. The unique molecular identifier (UMI) count per gene were normalized by the total UMI count in each cell and log transformed with the NormalizedData function in Seurat using 10,000 as the scale factor. The effects of the number of detected UMIs, the fraction of mitochondrial genes, and cell cycles on the gene expression values were corrected by regression using the ScaleData function in Seurat.
Before the clustering, we performed batch correction using Harmony across each single-cell sample, and integrate the gene expression matrix of all samples into a whole matrix. After cell-level quality filtering, we used DubStepR packages for the selection of highly variable genes. The same methods were applied for normalization, scaling, and batch correction. To identify contaminated cell clusters, we performed dimensionality reduction with 50 harmony dimensions, n_neighbors set to 50, and min_dist set to 0.1, followed by clustering with a resolution of 0.2.
5. Spatial transcriptome
Representative gastric cancer tumor samples were collected for the spatial transcriptomic sequencing. Samples were cut into 5×5 mm2 pieces, embedded in optimal cutting compound media and quickly frozen on dry ice. The frozen tissues were cryosectioned at 10 μm thickness by using the Thermo Scientific CryoStar NX50 cryostat and were placed in the capture area frames on the 10× Visium Spatial slides. Each sample slide was stained with H&E (hematoxylin Dako #S3309, eosin, Dako #CS701, bluing buffer #CS702, Dako, Glostrup, Denmark) and the brightfield images were captured via Leica whole-slide scanner at 10× resolution. Following tissue permeabilization, reverse transcription and cDNA amplification were processed by using Reagent Kit (#PN-1000184, PN-1000193, 10× Genomics). Visium spatial libraries were constructed using Visium spatial library construction kit (#PN-1000184, 10× Genomics) according to the manufacturer’s protocols. Finally, the libraries were sequenced using the Illumina NovaSeq6000 at least 100,000 reads per spot via pair-end 150 bp reading strategy.
6. Trajectory analysis
Trajectory analysis was conducted using Monocle 3.0, accessible at http://cole-trapnell-lab.github.io/monocle3/. Utilizing the final annotated CD8+ T cell subtype Uniform Manifold Approximation and Projection (UMAP), we employed Monocle 3 to construct learned graphs and determine cell order, without setting the partition_qvalue. To identify genes that change in expression over pseudotime, the graph_test function within Monocle 3 was used, enabling us to pinpoint significant genes false discovery rate < 10–50.
7. Infer copy number variations analysis
To identify malignant cell in the epithelial cell clusters, we utilized inferCNV (version 1.14.2), available at https://github.com/broadinstitute/inferCNV. We analyzed the count matrix designating normal sample pit cell and chief cell as the reference. The parameters set included a cutoff of 0.1, data scaling, denoising, a noise filter of 0.12, subcluster analysis mode, an hidden markov model type of i6, and the ward.D2 hierarchical clustering method.
8. Cell to cell interaction analysis
Interaction between CD8+ T cell including effector memory T cell (Tem), exhausted T cell (Tex), proliferative exhausted T cell (pTex) and malignant cell was analyzed using NicheNet [15]. For NicheNet analysis, we set CD8+ T cell as a sender (as the regulator cell) and malignant cell as a receiver (as the target cell). We used the provided ligand-target pairs and ligand-receptor pairs as inputs and we performed differential NicheNet analysis between MSI-H and MSS gastric cancer. We used the gene set which are provided in NicheNet (https://github.com/saeyslab/nichnetr). To identify important interaction signal, we also utilized CellChat between CD8+ T cell and malignant cell [16]. We used the gene set provided by CellChat for reference (https://github.com/sqjin/cellchat).
9. Defining CD8+ T cell existing spot in spatial transcriptome
Due to the low resolution of the Visium technology that couldn’t distinguish CD8+ T cell and malignant cell in the spot level, we calculated CD8+ T cell score using SEAGAL (ver. 2.4) python package. For the Tem we used GZMA, GZMB, IFNG, and PRF1 gene as signature genes, we used PDCD1, LAG3, TIGIT, and CTLA4 for Tex score. Next, we defined each Tem and Tex co-localized spot with the malignant cell when the score was over 0.2.
10. Calculation of malignant-CD8+ T cell interaction score
To quantify interaction between malignant cell and adjacent CD8+ T cell co-localized spot, we calculated the number of adjacent spots identified as CD8+ T cell co-localized spot. This count was the divided by seven which was the number of adjacent spot and core spot of Visium to derive a malignant-CD8+ T cell interaction score.
11. Bulk sequencing data deconvolution
We utilized CIBERSORTx to quantify the proportions of immune cell types within tumor microenvironment [17]. For signature matrix of 22 immune cell types, we used CIBERSORTx provided signature matrix. Then we compute the fractions of immune cell types in GSE66229 microarray data.
12. Survival analysis in public data
In GSE66229 dataset, we defined MSI status according to MLH1 imunohistochemical result that was provided by original article [18]. Then, we excluded epithelial-mesenchymal transition (EMT) subtype of gastric cancer from the original dataset due to EMT subtype showed distinct characteristics of MSI or MSS samples. Cox proportional hazard analysis in MSI-H and MSS samples was performed using lifeline library of python. For maximal survival differences according to IFNG, GZMB, LAG3, TIGIT, and CTLA4 expression in MSI-H samples, we used MaxStat R package. Overall survival of IFNG, GZMB, LAG3, TIGIT, and CTLA4 expression high and low patients was analyzed by log-rank tests with Kaplan-Meier curves. A p < 0.05 indicated statistical significance.
Go to :
Results
1. Higher proportion of CD8+ T cell in MSI-H gastric cancer
Droplet-based scRNA-seq was performed on tissue sample obtained from six patients. Clinicopathological features of patents were described in Table 1. After quality control and doublet removal, 45,087 cells from six tumor samples and two normal samples were included in the dataset. After multi-sample batch-effect correction and data normalization, unsupervised cell clustering was performed using the UMAP embedding 45,087 cells. The resulting UMAP plot highlights distinct clusters through color differentiation (Fig. 1A). To gain insights into the identity of each cell type, we superimposed gene expression levels of established markers onto the UMAP plot, providing a view of clusters (Fig. 1B). Further elucidating the composition of the identified clusters, a dot plot demonstrates proportions and average expression levels of marker genes across 10 cell types (Fig. 1C). The single cell atlas comprised of epithelial cell (n=3,188; EPCAM, CDH1), endothelial cell (n=1,270; PLVAP), stromal cell (n=1,326; THY1), CD8+ T cell (n=19,287; CD8A), CD4+ T cell (n=6,764; CD4), B cell (n=5,441; MS4A1), plasma cell (n=2,961; TNFRSF17), mast cell (n=1,354; KIT), neutrophil (n=1,485; CXCL8), and myeloid cell (n=2,011; CD163) (Fig. 1A and C). The UMAP plots demonstrate the differences in cell count across clusters within MSI-H, MSS, and normal samples (Fig. 1D). Examining the broader context, stacked bar plots illustrate the distribution of clusters among samples and demonstrate the fraction of cells within MSI-H, MSS, and normal samples (Fig. 1E and F). In MSI-H samples, a significantly higher proportion of tumor infiltrating CD8+ T cell was observed.
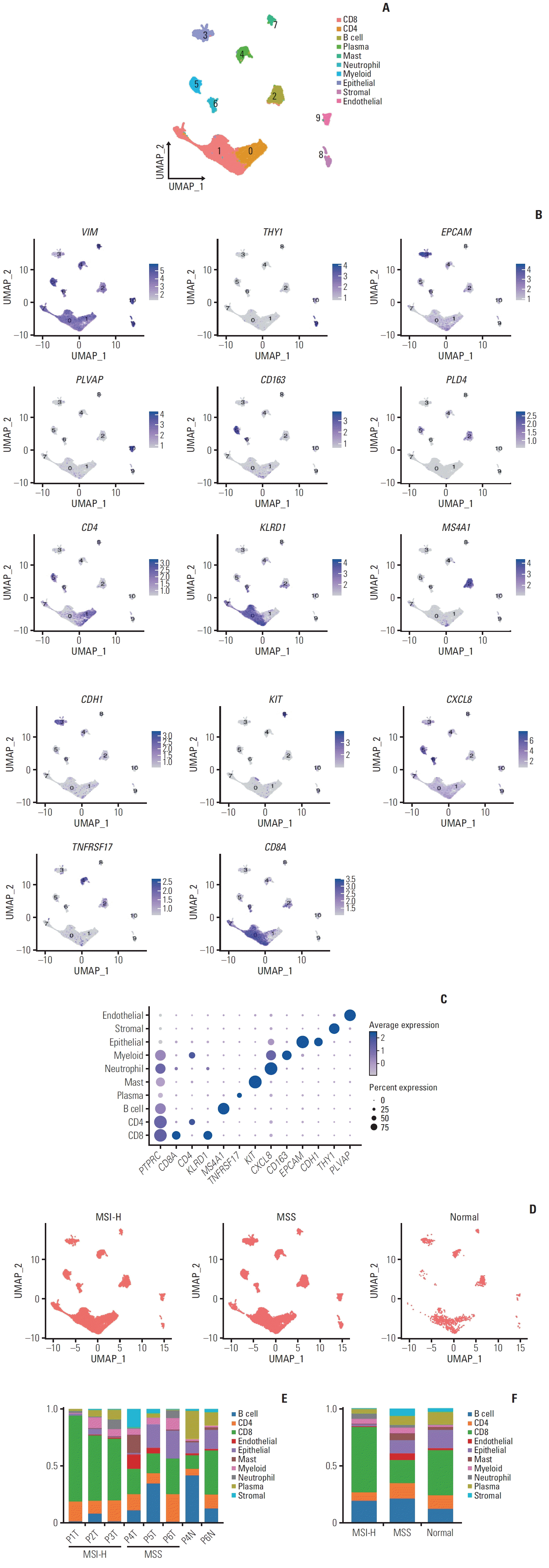 | Fig. 1.Single-cell atlas of microsatellite instability–high (MSI-H) and microsatellite stable (MSS) gastric cancer. (A) Uniform Manifold Approximation and Projection (UMAP) embeddings 45,087 cells. Clusters are highlighted in color. (B) The gene expression levels of known markers specific to each cell type are depicted on the UMAP plot. (C) Dot plot showing the proportions and average expression levels of marker genes for 10 cell types. (D) UMAP plot shows cells by MSI status. (E) Stacked bar plot shows the fraction of clusters by the samples. (F) Stacked bar plot shows the fraction of cells among MSI-H, MSS, and normal samples.
|
Table 1.
Clinicopathological features
EBV, Epstein-Barr virus; HER2, human epidermal growth factor receptor 2; L, lower third; LI, lymphatic invasion; M, middle third; MD, moderately differentiated; MSI, microsatellite instability; MSI-H, microsatellite instability–high; MSS, microsatellite stable; PD, poorly differentiated; PI, perineural invasion; VI, vascular invasion.
![]()
2. Effector function characteristics of exhausted T cell in MSI-H gastric cancer
After multi-sample batch-effect correction and data normalization, unsupervised cell clustering was performed, utilizing UMAP to embed 18,150 CD8+ T cell. This analysis identified nine distinct CD8+ T cell clusters, each characterized by unique gene signatures (Fig. 2A). A dot plot elucidates the proportions and average expression levels of marker genes, contributing to an understanding of CD8+ T cell subtypes (Fig. 2B). The single cell atlas comprised of naïve CD8+ T cell, activated naïve CD8+ T cell, effector memory CD8+ T cell (Tem), exhausted CD8+ T cell (Tex), proliferative exhausted CD8+ T cell (pTex), proliferative CD8+ T cell, mucosal-associated invariant T (MAIT) cell, FCER1G+ natural killer T (NKT) cell, and FCGR3A+ NKT cell (Fig. 2A). Violin plots show the expression of TOX, TOX2, CTLA4, and CXCL13 in Tex and pTex (Fig. 2C). The stacked bar plot reveals the distribution of CD8+ T cell subtypes among MSI-H, MSS, and normal samples (Fig. 2D). In MSI-H gastric cancer, Tem, Tex, pTex, and proliferative CD8+ T cell exhibited significantly higher proportions, whereas MAIT cell and NKT cell showed significantly higher proportions in MSS gastric cancer. In Tex and pTex, the exhaustion marker LAG3 was significantly upregulated in MSI-H gastric cancer; however, CXCL13 and TOX were significantly upregulated in MSS gastric cancer (Fig. 2E). Also, the effector function marker IFNG, GZMB, GZMH, and GZMK were significantly upregulated in Tex and pTex of MSI-H gastric cancer (Fig. 2F). The trajectory analysis, originating from naïve CD8+ T cell, demonstrates the differentiation of CD8+ T cell into Tex and pTex as pseudotime progressed (Fig. 2G). Furthermore, it was observed that despite the progression of exhaustion, the expression of IFNG remained elevated in MSI-H gastric cancer. Although the expression of GZMK and GZMH decreased as pseudotime progressed, the expression of these genes was higher in MSI-H gastric cancer (Fig. 2H).
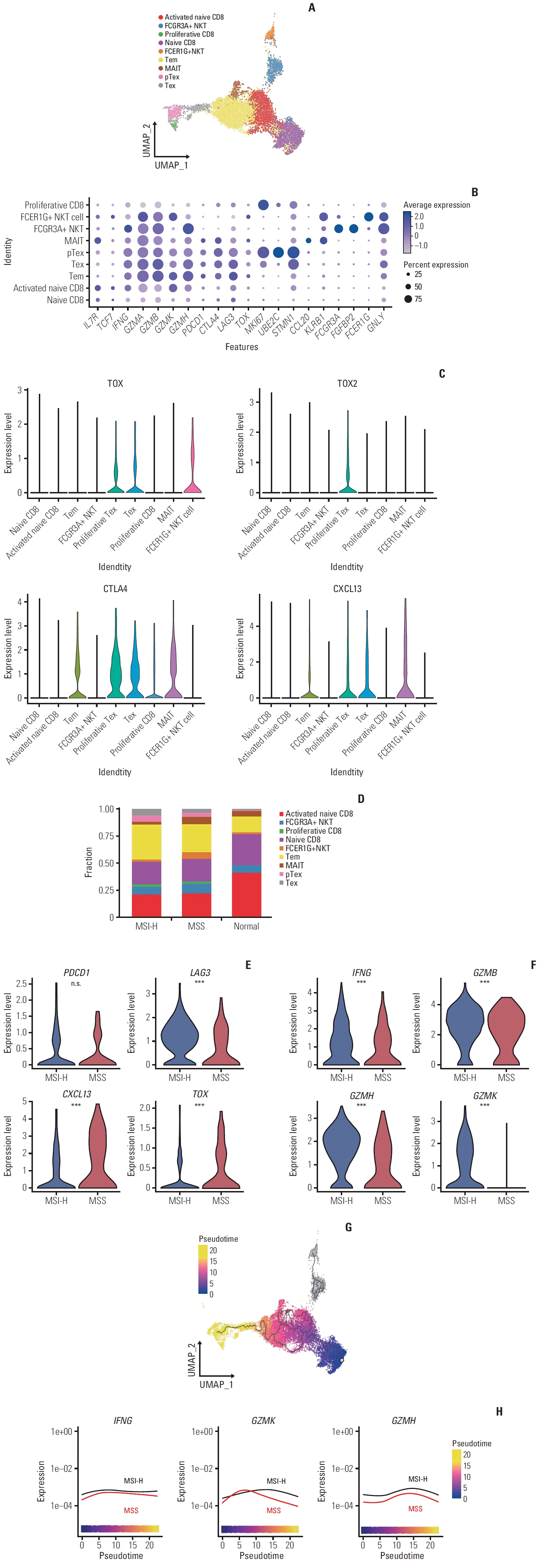 | Fig. 2.Sub-cellular analysis of CD8+ T cell. (A) Uniform Manifold Approximation and Projection (UMAP) plot embeddings 18,150 cells of CD8+ T cell. Clusters are highlighted in color. (B) Dot plot shows the proportions and average expression levels of marker genes for CD8+ T cell subtypes. MAIT, mucosal-associated invariant T; NKT, natural killer T; pTex, proliferative exhausted CD8+ T cell; Tem, effector memory CD8+ T cell; Tex, exhausted CD8+ T cell. (C) Violin plot shows the expression of exhaustion markers in T cell subtypes. (D) Stacked bar plot shows the fraction of CD8+ T cell subtypes between microsatellite instability–high (MSI-H), microsatellite stable (MSS) and normal samples. The color scale is the same as in A. (E) Violin plots compared marker genes of exhaustion in exhausted CD8+ T cell and proliferative exhausted CD8+ T cell between MSI-H and MSS samples (Wilcoxon rank sum test; ***p < 0.001). (F) Violin plots compared marker genes of effector function in exhausted CD8+ T cell and proliferative exhausted CD8+ T cell between MSI-H and MSS samples (Wilcoxon rank sum test; ***p < 0.001). (G) UMAP plot demonstrates trajectory analysis, with colored by inferred pseudotime. The trajectory of the NKT cells is represented in grey color, which means the independent trajectory. (H) Line plot illustrating the gene expression of IFNG, GZMK, and GZMH over the inferred pseudotime trajectory in CD8+ T cell.
|
3. Sub-cellular analysis of epithelial cell
A comprehensive analysis of epithelial cell was conducted to unravel the cellular subtype according to the molecular characteristics (Fig. 3A). We can identify the expression levels of epithelial marker genes across clusters, including TFF1 and MUC5AC for pit cell, LIPF for chief cell, APOB and ALDOB for enterocyte, and CEACAM5 and CCND2 for malignant cell (Fig. 3B). The chromosomal landscape, revealing inferred large-scale copy number variations (CNV) within the epithelial sub-clusters, identified clusters 0, 1, 3, 4, 6, and 8 as malignant cell clusters (Fig. 3C). Based on the gene expression patterns observed in the dot plot and the inferred CNV within the sub-clusters of epithelial cells, we have annotated sub-clusters on the UMAP (Fig. 3D).
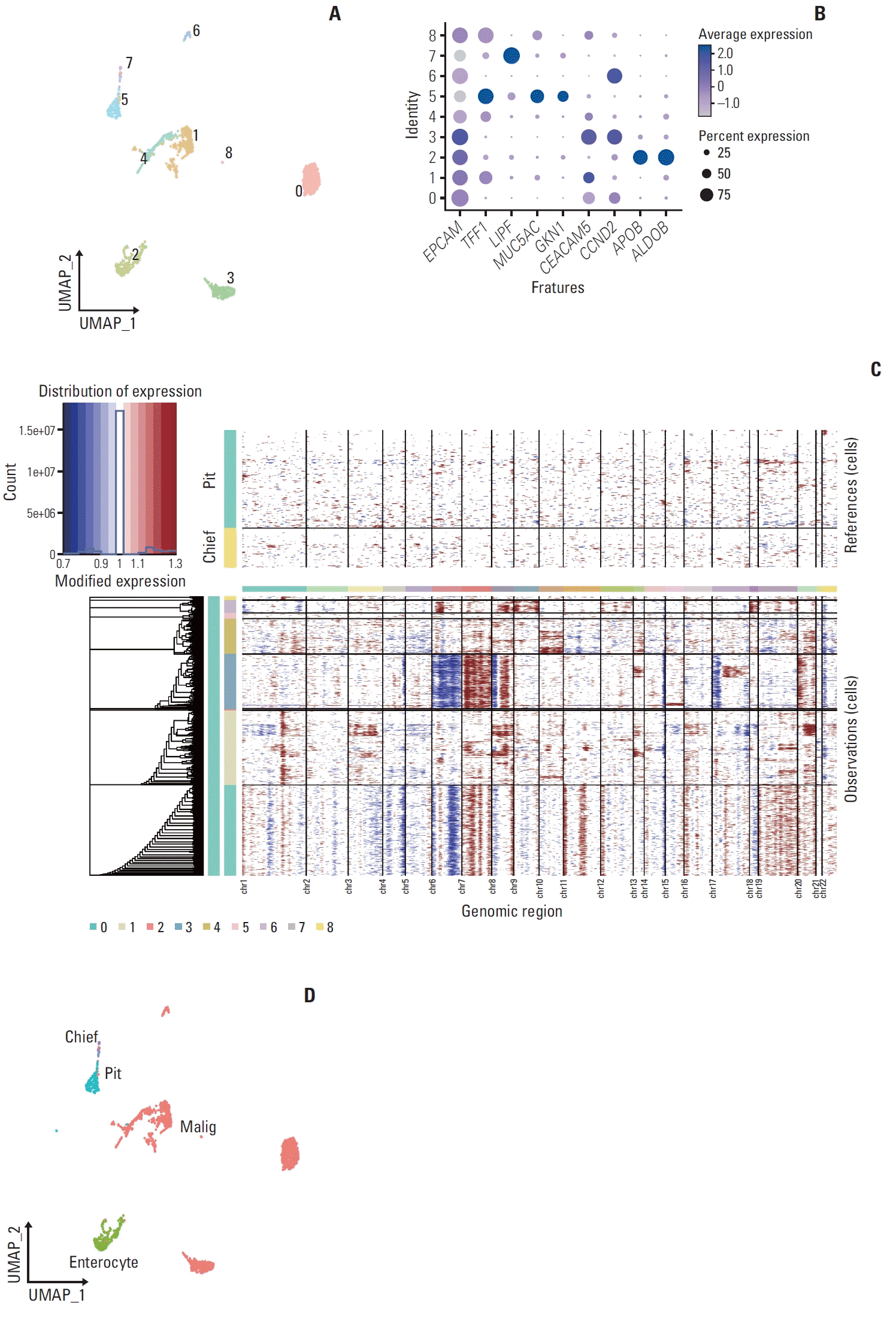 | Fig. 3.Sub-cellular analysis of epithelial cell. (A) Uniform Manifold Approximation and Projection (UMAP) plot embeddings 3,066 cells of epithelial cell. Clusters are highlighted in color. (B) Dot plot illustrates the proportions and average expression levels of epithelial marker genes such as TFF1 and MUC5AC for pit cell; LIPF for chief cell; APOB and ALDOB for enterocyte; CEACAM5 and CCND2 for malignant cell across the clusters. (C) A chromosomal landscape shows the inferred large-scale copy number variations within the epithelial sub-clusters, with annotation tracks on the left correlating to the respective clusters and chromosome numbers indicated at the bottom. (D) UMAP plot of 3,066 epithelial cells are marked for sub-cellular analysis, with cells color-coded by type: pit cell, chief cell, enterocyte, and malignant cell.
|
4. Retained interferon-γ signaling pathway of exhausted T cell in MSI-H gastric cancer
To examine the intercellular communication between malignant cell and CD8+ T cell, receptor-ligand interactions were identified. The scaled expression of IFNG and GZMB ligands was observed to be retained in Tex and pTex compared to Tem in MSI-H gastric cancer. However, this distinctive pattern was not observed in MSS gastric cancer (Fig. 4A). Subsequently, the inferred interferon γ (IFN-γ) signaling pathway networks calculated by CellChat demonstrate the enriched network between Tex, pTex and malignant cell in MSI-H gastric cancer and decreased network between Tex, pTex and malignant cell in MSS gastric cancer (Fig. 4B and C).
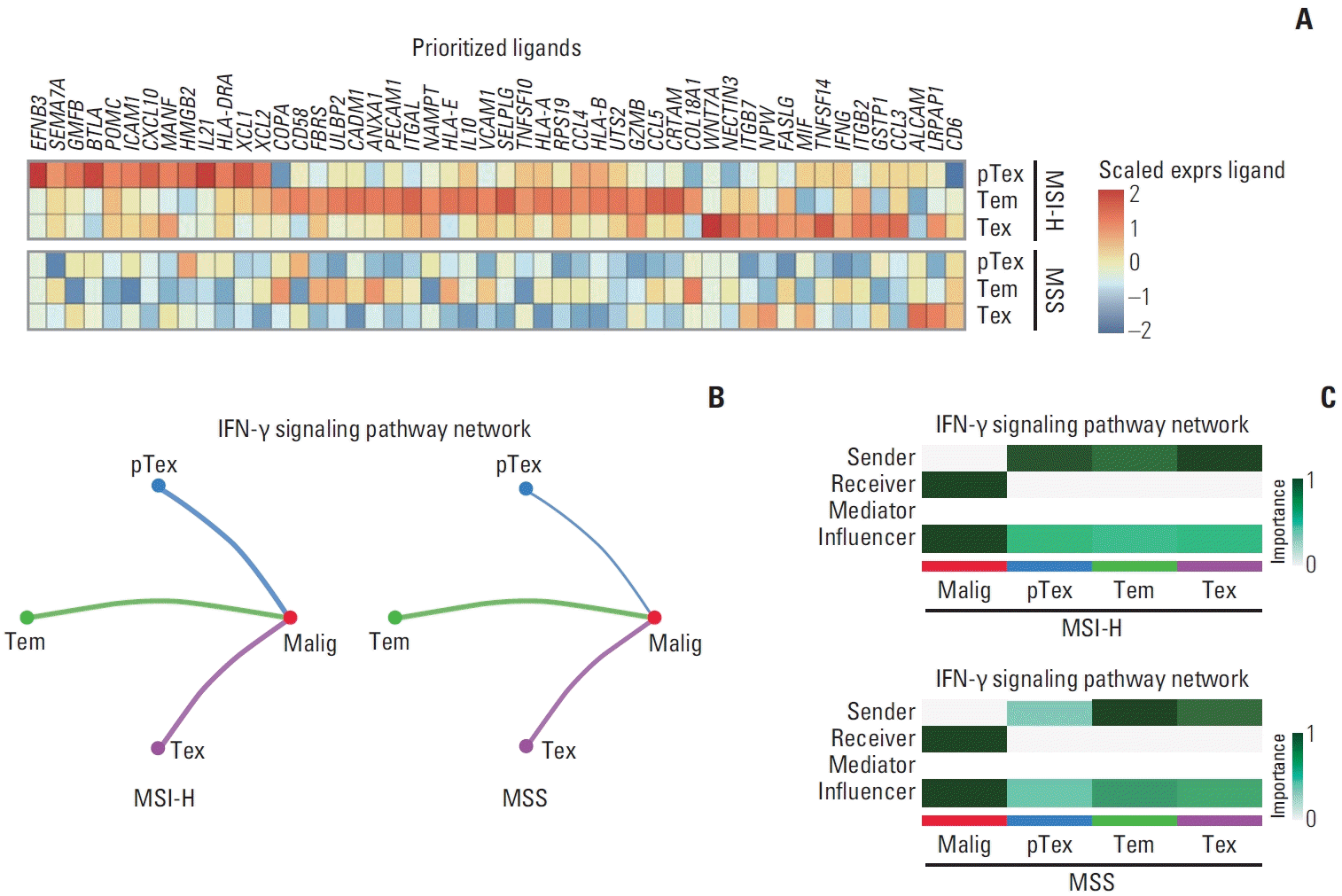 | Fig. 4.Cell-to-cell interaction analysis. (A) Heatmap of expression of prioritized ligands from NicheNet between microsatellite instability–high (MSI-H) and microsatellite stable (MSS) gastric cancer. Heatmap is colored by scaled expression of the ligands (top). (B) The inferred interferon γ (IFN-γ) signaling networks calculated by CellChat in MSI-H (left) and MSS samples (right). Edge width represents the communication probability. (C) The heatmap displays the relative importance of each cell group within the IFN-γ signaling network for MSI-H and MSS gastric cancer, based on four computed network centrality measures. pTex, proliferative exhausted CD8+ T cell; Tem, effector memory CD8+ T cell; Tex, exhausted CD8+ T cell.
|
5. Higher IFN-γ response between adjacent exhausted T cell and malignant cell in MSI-H gastric cancer
We further conducted spatial transcriptomic analysis to verify molecular characteristics with spatial distribution. The spatial transcriptomic data shows the expression of EPCAM and CD8A at each spot. Overall, in MSI-H gastric cancer, it was observed that the distribution of CD8A expression spots is diffuse and prominently elevated compared to MSS gastric cancer (Fig. 5A and B). This suggests a significant presence of tumor infiltrating CD8+ T cell in MSI-H gastric cancer.
 | Fig. 5.Epithelial cell and CD8+ T cell in spatial transcriptomic data. (A) Spatial transcriptomic data visualized with color coding based on the gene expression including EPCAM (left) and CD8A (right) in microsatellite instability–high (MSI-H) gastric cancer. (B) Spatial transcriptomic data visualized with color coding based on the gene expression including EPCAM (left) and CD8A (right) in microsatellite stable (MSS) gastric cancer.
|
To assess the interaction between Tem and Tex with malignant cell, co-localization scores for Tem and Tex were evaluated using cor.test and visualized in the spatial transcriptome (Fig. 6A and B). The fraction of co-localized spots for Tem and Tex was found to be significantly higher in MSI-H gastric cancer (Fig. 6C). Based on the spatial distribution of malignant cell and Tem, Tex spots, we calculated interaction scores (Fig. 6D). Significantly higher interaction scores were observed in MSI-H which indicating the abundant presence of Tem and Tex cells in close proximity to malignant cell in MSI-H gastric cancer (Fig. 6E). Finally, we evaluated the IFN-γ pathway between the adjacent Tex and malignant cell, which demonstrates significantly higher interaction in MSI-H gastric cancer. (Fig. 6F).
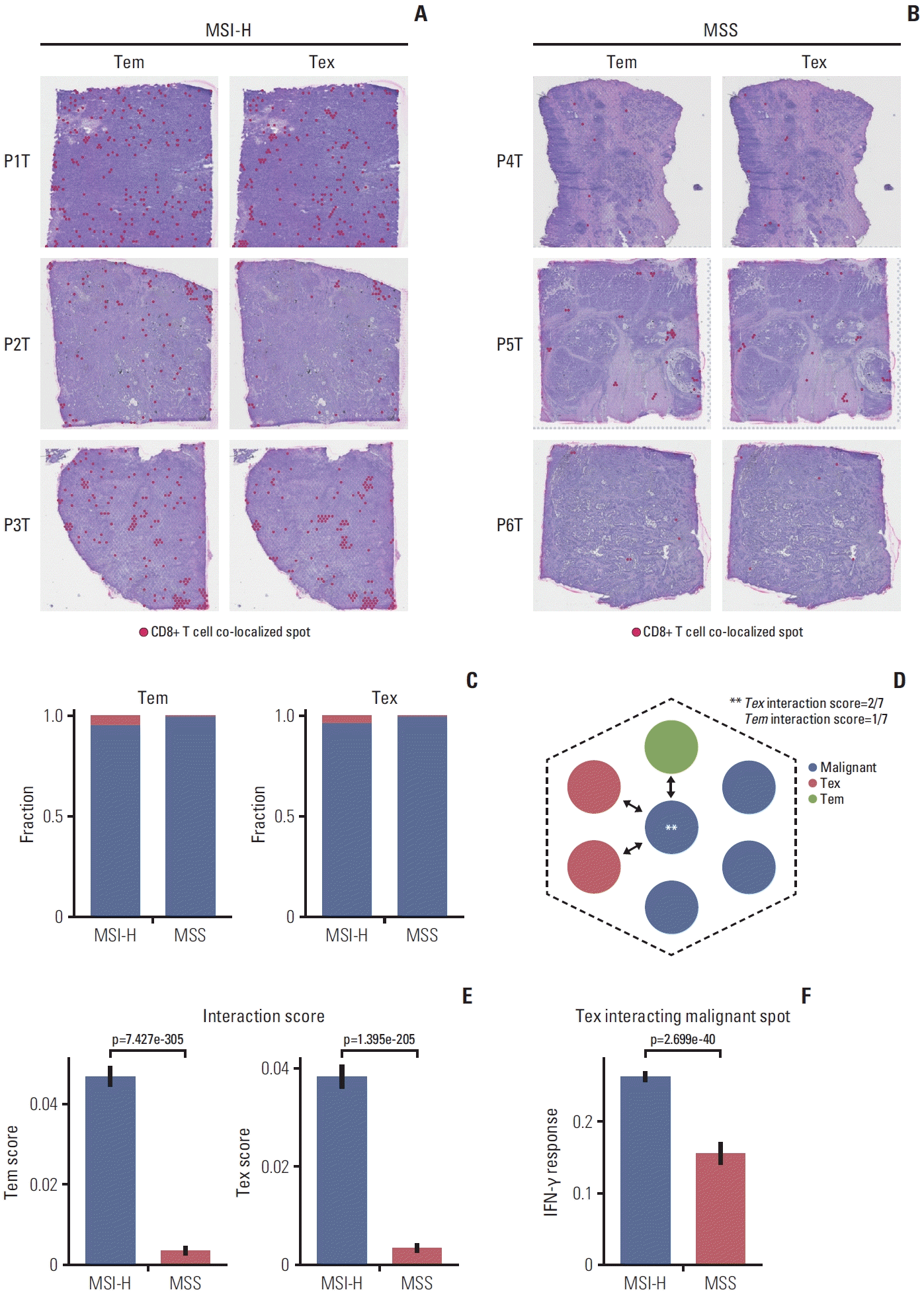 | Fig. 6.Spatial architecture of CD8+ T cell. (A) Effector memory CD8+ T cell (Tem) and exhausted CD8+ T cell (Tex) co-localized spots were marked on the spatial images in microsatellite instability–high (MSI-H) gastric cancer. (B) Tem and Tex co-localized spots were marked on the spatial images in microsatellite stable (MSS) gastric cancer. (C) Bar plot shows the fraction of Tem co-localized spot (left) and Tex co-localized spot (right) in integrated 3 MSI-H and 3 MSS spatial transcriptome samples. (D) Schematic representation of malignant cell - CD8+ T cell interaction score (i.e., malignant cell - Tex interaction score is 2/7 and malignant cell - Tem interaction score is 1/7). (E) Box plot shows malignant cell - Tem interaction scores (left) and malignant cell - Tex (right) in integrated three MSI-H and three MSS spatial transcriptome samples. (F) Box plot shows Hallmark IFN-γ response score from MSigDB in the exhausted CD8+ T cell interacting malignant spots in integrated three MSI-H and three MSS spatial transcriptome samples.
|
6. Validation using bulk level transcriptome data
Further examination was performed by reanalyzing bulk transcriptomic ACRG data (GSE66229) using CIBERSORTx deconvolution, revealing a significantly higher presence of CD8+ T cells in MSI-H compared to MSS gastric cancer (Fig. 7A). In the heat map, it was observed that cytotoxic and exhaustion markers are highly expressed in MSI-H gastric cancer (Fig. 7B). The scatter plot between IFNG and marker genes of exhaustion illustrates a higher coefficient value in MSI-H gastric cancer, indicating elevated expressions of IFNG at the same level of CXCL13 and LAG3 expression (Fig. 7C). Survival analysis demonstrates the significant better survival in MSI-H patients according to the expression of effector function and exhaustion marker genes (Fig. 7D and E).
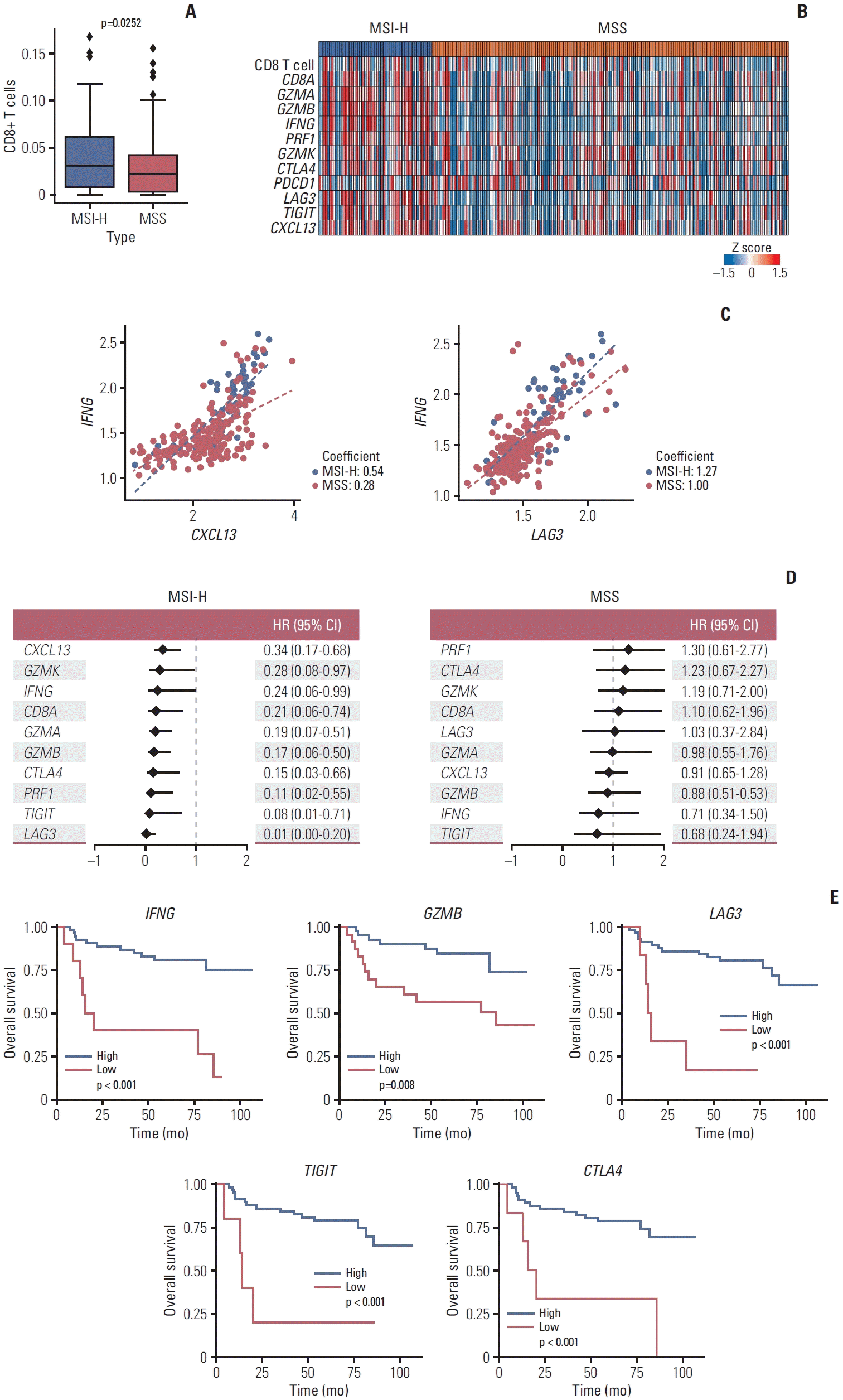 | Fig. 7.Validation using bulk level transcriptome data. (A) Box plot shows fraction of CD8+ T cells calculated by CIBERSORTx between microsatellite instability–high (MSI-H) and microsatellite stable (MSS) samples in ACRG cohort (GSE66229). (B) Heatmap of normalized gene expression including cytotoxic CD8+ T cell marker genes and exhausted CD8+ T cell marker genes in ACRG cohort. The heatmap is colored by z score across by the samples. (C) Scatter plot between IFNG and exhausted CD8+ T cell marker including CXCL13 (left) and LAG3 (right). The scatterplot is colored by MSI status and dash lines in each plot represents linear regression line. Coefficient value of linear regressions is marked on the plot. (D) Univariate Cox proportional hazard analysis of effector memory CD8+ T cell marker genes and exhausted CD8+ T cell marker genes in MSI-H patients (left) and MSS patients (right). CI, confidence interval; HR, hazard ratio. (E) Kaplan-Meier curves showing significant difference between gene expression high group and low group which is calculated by MaxStat R package in MSI-H patients. The plot is colored by the gene expression groups.
|
Go to :
Discussion
In the KEYNOTE-059 trial, patients with MSI-H gastric cancer experienced a higher overall response rate compared to patients with non-MSI-H gastric cancer (MSI-H vs. non-MSI-H; 57.1% vs. 9.0%) [19]. The increased response of MSI-H cancer to immunotherapy can be attributed to several factors. Firstly, these tumors have a higher mutational burden, leading to the production of more neoantigens, which can be recognized by the immune system [20]. Additionally, MSI-H gastric cancer is characterized by high gene expression for CD8+ T cell and IFN-γ signatures, indicating the immunogenicity. A high prevalence of PD-L1 positivity has also been reported in MSI-H gastric cancer, along with a significant infiltration of tumor-infiltrating lymphocytes [6]. However, the molecular characteristics of the tumor-infiltrating CD8+ T cell subtypes in MSI-H and MSS gastric cancer have not been evaluated yet.
Our current study provided a comprehensive molecular characterization of tumor-infiltrating CD8+ T cell in MSI-H and MSS gastric cancer. We identified nine distinct CD8+ T cell subtypes. Among them, Tem, Tex, pTex, and proliferative CD8+ T cell exhibited significantly higher proportions in MSI-H gastric cancer whereas MAIT cell and NKT cell showed significantly higher proportions in MSS gastric cancer. Interestingly, pTex shows a higher expression of proliferation markers (MKI67, UBE2C, STMN1) compared to Tex, although both subtypes exhibit expression of the exhaustion markers. A recent study demonstrates that pTex is beneficial to the survival outcomes of patients with human papillomavirus–positive head and neck squamous cell carcinoma [21]. The pTex was found to aggregate in the antigen-presenting cell areas, where T cell activation and proliferation signaling pathways were activated. The pTex represents an independent branch of Tex cell and might act as a T cell pool, providing modest but persistent antitumor immunity through its prolonged cell survival and highly specialized cytotoxic capacity. In our study, both Tex and pTex demonstrate higher expression levels of IFNG and GZMB in MSI-H gastric cancer, implying cytotoxic activity.
The IFN-γ, encoded by the IFNG gene, plays a pivotal role in activating cellular immunity, subsequently stimulating the antitumor immune response [22,23]. Within the tumor microenvironment, IFN-γ acts as a cytotoxic cytokine alongside granzyme B and perforin, initiating apoptosis in tumor cell [24]. Simultaneously, it facilitates the synthesis of immune checkpoint inhibitory molecules, contributing to other immune-suppressive mechanisms [25]. The advent of ICIs, including PD-1, PD-L1, and CTLA-4, brought hope for patients. Upon binding to PD-1, these antibodies trigger CD8+ T cell to release IFN-γ, activating its receptor and thereby increasing interleukin-12 production in the tumor microenvironment. The concentration of IFN-γ and the induction of the IFNG gene could potentially serve as biomarkers for predicting patient responses to immunotherapy [26]. In this study, we observed the retained expression of IFNG in Tex and pTex within MSI-H gastric cancer. Furthermore, cell-to-cell interaction analysis and spatial transcriptome analysis unveiled the persistence of the IFN-γ pathway between Tex, pTex, and malignant cell. The spatial transcriptome analysis specifically highlighted the IFN-γ pathway between neighboring Tex and malignant cell, showcasing a significantly elevated interaction in MSI-H gastric cancer. This differs from the traditional concept that effector function decreases as T cell exhaustion progresses, and it is believed to be attributed to the unique molecular biology of MSI-H gastric cancer.
Although our study has yielded remarkable findings, it is not without its limitations. First, our results, which are based on genetic analysis and inferred cell networking, require further functional validation. Second, while we have identified the effector function characteristics of exhausted T cells in MSI-H gastric cancer, the mechanism by which ICIs enhance the survival of patients with MSI-H gastric cancer remains uncertain.
In summary, our study demonstrates novel findings that IFN-γ signaling pathway is retained in exhausted T cell of MSI-H gastric cancer. Our integrated analysis provides a holistic perspective on the immune microenvironment in MSI-H gastric cancer, and we anticipate future work to unravel the mechanism of how ICIs work in exhausted T cell of MSI-H gastric cancer.
Go to :
 XML Download
XML Download