

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Pulmonary fibrosis (PF) is a progressive, age-related lung disease that can lead to structural remodeling, impaired gas exchange, respiratory failure and death [1]. The causes of PF are complex, and the median survival after diagnosis is extremely short [2]. Currently, the only agents recommended by evidence-based guidelines are pirfenidone and nintedanib, but they cannot effectively improve patients' quality of life and inhibit disease progression [3-7]. Therefore, further research into the mechanisms of PF and effective therapies is particularly important.
Oxidative stress (OS) is involved in the process of PF through three main pathways: directly causing damage to alveolar epithelial cells and promoting apoptosis; increasing the aggregation of inflammatory cells, overexpressing tumor necrosis factor-α (TNF-α), interleukin (IL) and nuclear transcription factor-κB (NF-κB) and releasing large amounts of inflammatory factors; and directly stimulating the production of transforming growth factor-β1 (TGF-β1) [8-11]. TGF-β1 is a key inducer of PF progression and stimulates the production of reactive oxygen species (ROS) through the activation of the Smad 2/3 and mitogen-activated protein kinase (MAPK) pathways [12]. In turn, high levels of ROS promote the expression of TGF-β1, which induces the transformation of fibroblasts into myofibroblasts and accelerates the development of PF [13]. MAPKs are members of the serine-threonine kinase superfamily and consist of three MAPK subfamilies: the extracellular signal-regulated kinase (ERK), p38 MAPK and c-Jun-n-terminal kinase (JNK) [14]. Chien et al. [15] showed that the phosphorylation levels of p38, ERK and JNK were significantly elevated in the lung tissue of mice with bleomycin-induced PF, suggesting that p38, ERK and JNK are involved in the progression of PF. Wang et al. [16] reported that peptide DR8 ameliorated bleomycin-induced PF through inhibition of TGF-β1/MAPK pathway activation and attenuation of oxidative damage. Therefore, we suggest that inhibiting TGF-β1/MAPK pathway activation may be an effective treatment for PF.
TGF-β1 also induces alveolar epithelial cell injury by inhibiting adenosine monophosphate-activated protein kinase (AMPK) activity. AMPK is a 'switch' that regulates energy metabolic homeostasis in the body and is one of the key factors contributing to many fibrotic diseases, including PF [12,17]. AMPK has an inhibitory effect on the inflammatory response in lung diseases, and its mechanism may be related to the regulation of the expression of downstream genes, such as silent information regulator 1 (sirtuin, SIRT1), peroxisome proliferator-activated receptor-gamma coactivator-1α (PGC-1α), and p53 [18]. Heme oxygenase-1 (HO-1) is a rate-limiting enzyme that catalyzes the degradation of heme, and several studies have confirmed the protective role of heme-induced HO-1 in acute lung injury [19], pulmonary hypertension [20] and chronic obstructive pulmonary disease [21]. AMPK/HO-1 activation plays a critical role in several physiological processes, including anti-inflammatory, antioxidant, antiapoptotic and vasculoprotective effects [22,23]. In addition, Yang et al. [24] reported that hemin-induced HO-1 ameliorated carbon tetrachloride-induced liver fibrosis in rats by regulating peroxisome proliferation-activated receptor (PPARγ) and NF-κB expression. Cho et al. [25] reported that rosiglitazone-induced HO-1 inhibits lipopolysaccharide-mediated lung inflammation by modulating the PKCα/AMPKα/p38MAPKα/SIRT1/PPARγ pathway. Therefore, in the present study, hemin was used to induce HO-1, and the expression and phosphorylation levels of proteins related to the TGF-β1/MAPK and AMPK/SIRT1/PGC-1α/HO-1/NF-κB pathways were detected in the lung tissue of mice with bleomycin-induced PF. Additionally, the protective effect of hemin on PF and the possible underlying mechanisms were investigated.
METHODS
Animals
Male C57BL/6 mice (weight 20–25 g) were purchased from Changsha Tianqin Biotechnology [Certificate number SCXK (XIANG) 2019-0014]. The animals were maintained on a 12-h day/12-h night cycle at a temperature of 20°C–24°C. Mice were allowed to eat and drink ad libitum. All the experimental procedures were approved by the Animal Experimental Ethics Committee of Wannan Medical College (LISC-2020-157).
Grouping and design of animal experiments
Male C57BL/6 mice were randomized into control, bleomycin and bleomycin + hemin groups, with 15 mice in each group. In accordance with the methods of Ma et al. [26], mice were anesthetized, 3 mg/kg bleomycin was injected intratracheally into the trachea of mice in the bleomycin and bleomycin + hemin groups to establish the PF model, and saline was administered to the control group. Mice in the bleomycin + hemin group received intraperitoneal injections of hemin at a dose of 30 mg/kg (once every other day) for 7 days prior to bleomycin stimulation and were continuously treated with hemin for 21 days after bleomycin injection. The preparation, dosage and method of administration of hemin were based on previous studies [19,27,28]. After the last administration of the drug, the mice in each group were weighed, blood was collected from the eyeballs, the animals were sacrificed, the lung tissue was weighed, and the lung coefficient was calculated (lung weight/body weight; mg/g).
Materials
Bleomycin sulfate (9041-93-4, purity ≥ 90%) and hemin chloride (16009-13-5, purity ≥ 97%) were purchased from Hefei Bomei Biotechnology. The α-SMA antibody was purchased from Beijing Bioworld Technology; the GAPDH antibody was purchased from Shanghai Beyotime Biotechnology; the collagen I antibody was purchased from Affinity Biosciences; and TGF-β1, AMPK, p-AMPK, SIRT1 and PGC-1α antibodies were purchased from Abcam; and p38, p-p38, ERK1/2, p-ERK1/2, JNK, p-JNK, NF-κB p65, p-NF-κB p65 and HO-1 antibodies were purchased from Cell Signaling Technology.
Histopathological examination
Fresh lung tissues were fixed in 4% paraformaldehyde for 48 h and then dehydrated in gradient ethanol before being processed into wax blocks. The wax blocks were cut into 5 μm thick sections, and the sections were deparaffinized in xylene and then placed in gradient ethanol and double-distilled water and stained according to the instructions of the HE (Fuzhou Phygene Science and Technology) and Masson (Hefei Bomei Biotechnology) staining kits. ImageJ software was used for image analysis of Masson's staining results (Broken Symmetry Software).
Determination of the activity or content of malondialdehyde (MDA), superoxide dismutase (SOD) and catalase (CAT) in lung tissues
An appropriate amount of lung tissue was removed, saline or hydrolysate was added, and the mixture was centrifuged (4°C, 3,000 rpm, 10 min). The supernatant was extracted, after which the MDA, SOD and CAT kits (Nanjing Jiancheng Bioengineering Institute) were used to detect the activity or content of MDA, SOD and CAT in the lung tissue.
Measurement of the serum levels of IL-6 and TNF-α
The serum of mice in each group was collected, and the levels of IL-6 and TNF-α in the serum of mice in each group were detected by using ELISA (IL-6 and TNF-α; Wuhan Elabscience Biotechnology) kits, which were operated strictly according to the instructions.
Immunohistochemical detection of α-SMA and collagen I expression in lung tissue
Sections were deparaffinized in xylene and then placed in gradient ethanol, and double distilled water, and immunohistochemistry experiments were performed using a concentrated SABC-POD (rabbit IgG) kit (Wuhan Boster Biological Technology) according to Huang et al. [29]. The sections were immersed in sodium citrate solution for antigen retrieval (heat in a microwave oven to 98°C–100°C and then on a low heat for 10 min). The sections were washed with ultrapure water, followed by 3% hydrogen peroxide to inactivate peroxidase (room temperature, 10 min). Then, the goat serum (37°C, 1 h) was added, and α-SMA and collagen I primary antibodies (1:150) were subsequently added, after which the sections were placed in a refrigerator at 4°C overnight. The sections were then removed and placed in an oven (37°C, 1 h) to allow the temperature to recover, after which secondary antibodies were added to the sections dropwise (37°C, 1 h), followed by SABC added dropwise (37°C, 1 h) and staining using an HRP-DAB kit (Beijing Tiangen Biochemical Technology). The images were analysed and processed using ImageJ software.
Western blotting
Appropriate amounts of lung tissue were removed, RIPA lysis buffer was added, homogenized and allowed to stand for 2 h, followed by centrifugation (4°C, 12,000 rpm, 10 min). The supernatant was extracted, and the protein concentration was determined using a BCA kit (Shanghai Beyotime Biotechnology). A 10% SDS polyacrylamide gel was prepared, followed by sample loading, electrophoresis (80-mV constant pressure electrophoresis until the target proteins were separated). The proteins were subsequently transferred to a PVDF membrane (a constant current of 200 mA, 1–2 h), after which the membranes were immersed in 5% skim milk or BSA (4°C, 1 h). The membranes were subsequently incubated with TGF-β1, p38, p-p38, ERK1/2, p-ERK1/2, JNK, p-JNK, AMPK, p-AMPK, HO-1, PGC-1α, SIRT1, NF-κB p65, p-NF-κB p65 and GAPDH primary antibodies (1:1,000, 4°C, overnight), and the next day, the membranes were incubated with the corresponding secondary antibodies (4°C, 2 h), washed with TBST buffer, incubated with fluorescent liquid and exposed. The bands were analyzed using ImageJ software.
Statistical analysis
The data are presented as the means ± standard deviations. Statistical analysis was performed using GraphPad Prism 8 software (version 8.0; GraphPad Software, Inc.). One-way analysis of variance (ANOVA) was performed for multiple comparisons between groups. The Tukey’s test was used for pairwise comparisons. The survival rates were recorded, estimated by the Kaplan–Meier method and were compared by a log-rank test. p < 0.05 was considered to indicate statistical significance.
RESULTS
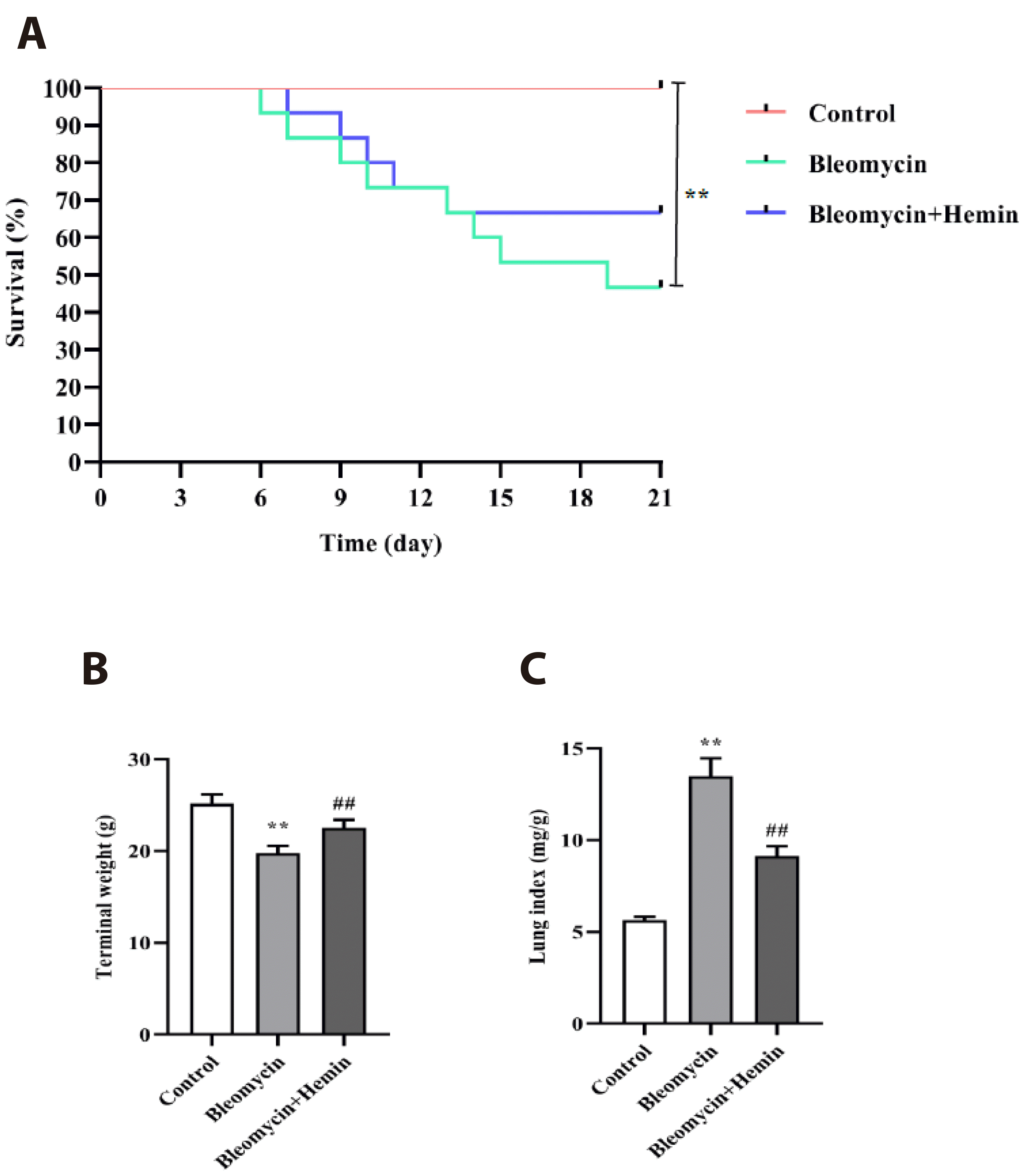
Effects of hemin on the survival rate, terminal body weight and lung indices of mice with PF
Compared with those in the control group, the mice in the bleomycin group had significantly lower survival rates (p < 0.01) (Fig. 1A) and terminal body weights (p < 0.01) (Fig. 1B) and a significantly greater lung index (p < 0.01) (Fig. 1C). The difference in survival among the mice in the bleomycin + hemin group was not statistically significant (p > 0.05), although there was an increase in survival. Pretreatment with hemin resulted in a marked increase in the final body weight (p < 0.01) and a marked decrease in the lung indices of the mice (p < 0.01).
Effect of hemin on morphological and pathological changes in the lung tissue of mice with PF
As shown in Fig. 2A, the lung tissue of mice in the control group was light pink in color, smooth, soft and elastic to the touch, with no nodules or hemorrhage points, and that of mice in the bleomycin group was grayish white, rough, hard and inelastic to the touch. The nodules were distinct and abundant with multiple hemorrhages points. The lung tissue of mice in the bleomycin + hemin group was slightly darker in color, slightly rougher, with reduced elasticity, and with a small number of nodules and hemorrhage points in some areas.
After HE and Masson staining (Fig. 2B–D), we found that the alveolar structure of the lung tissue of the mice in the control group was complete and clear, with thin alveolar walls, normal spacing, no edema or inflammatory cell exudation, and only a small amount of scattered blue collagen fibers. The lung tissue structure of the mice in the bleomycin group was severely damaged, with thickening of the alveolar wall, an increase in the width of the septa, and the appearance of large, dark blue collagen fibers between the lung interstitium. The bleomycin + hemin group showed varying degrees of reduction in all of the above pathological impairments.
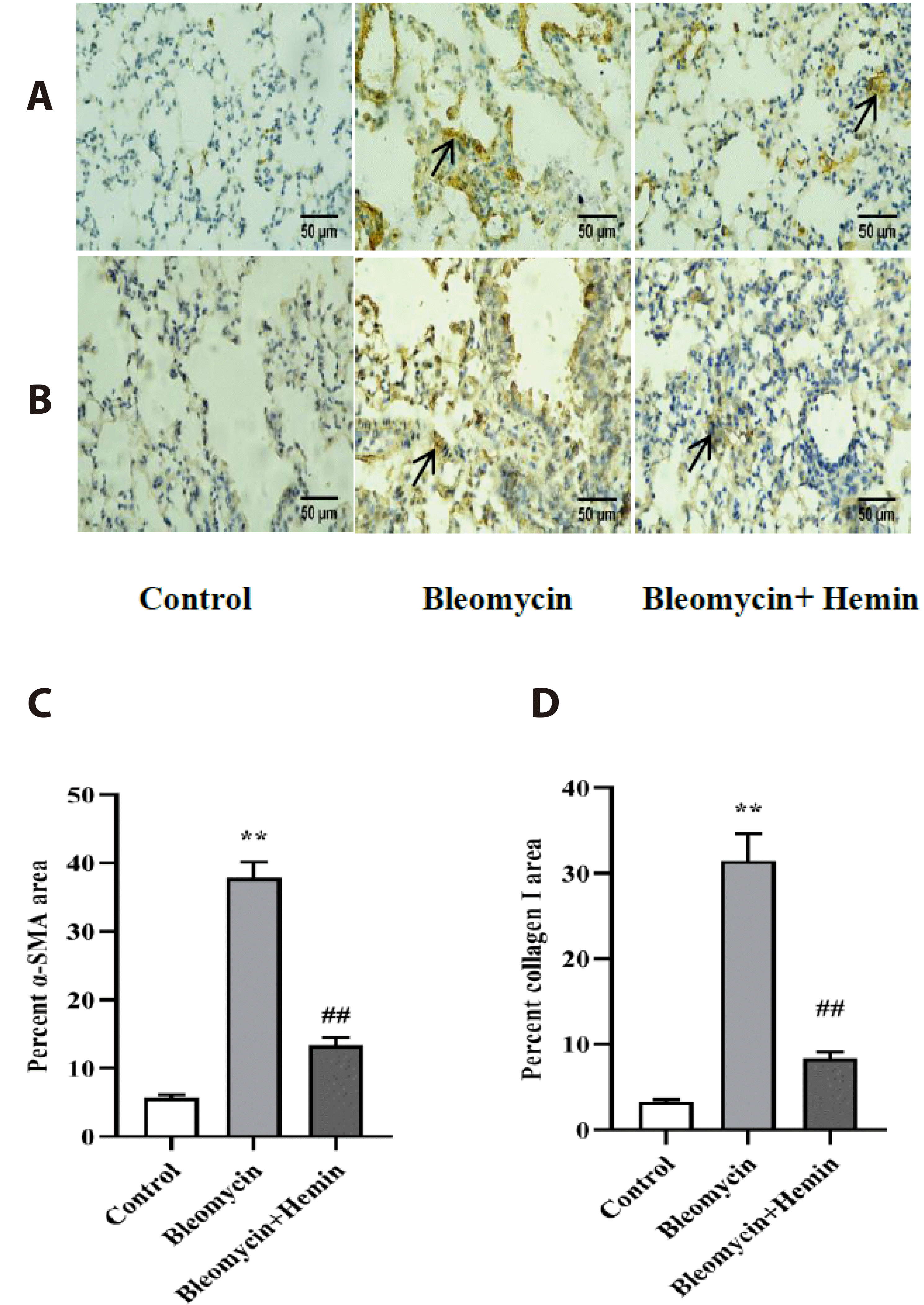
Effect of hemin on the expression of the PF marker proteins α-SMA and collagen I
Immunohistochemistry revealed only a small number of scattered brownish-yellow α-SMA- and collagen I-positive cells in the control group (Fig. 3). In the bleomycin group, the expression of SMA and collagen I was obviously increased (p < 0.01). Compared with those in the bleomycin group, the expression of α-SMA and collagen I was significantly lower in the hemin pretreatment group (p < 0.01).
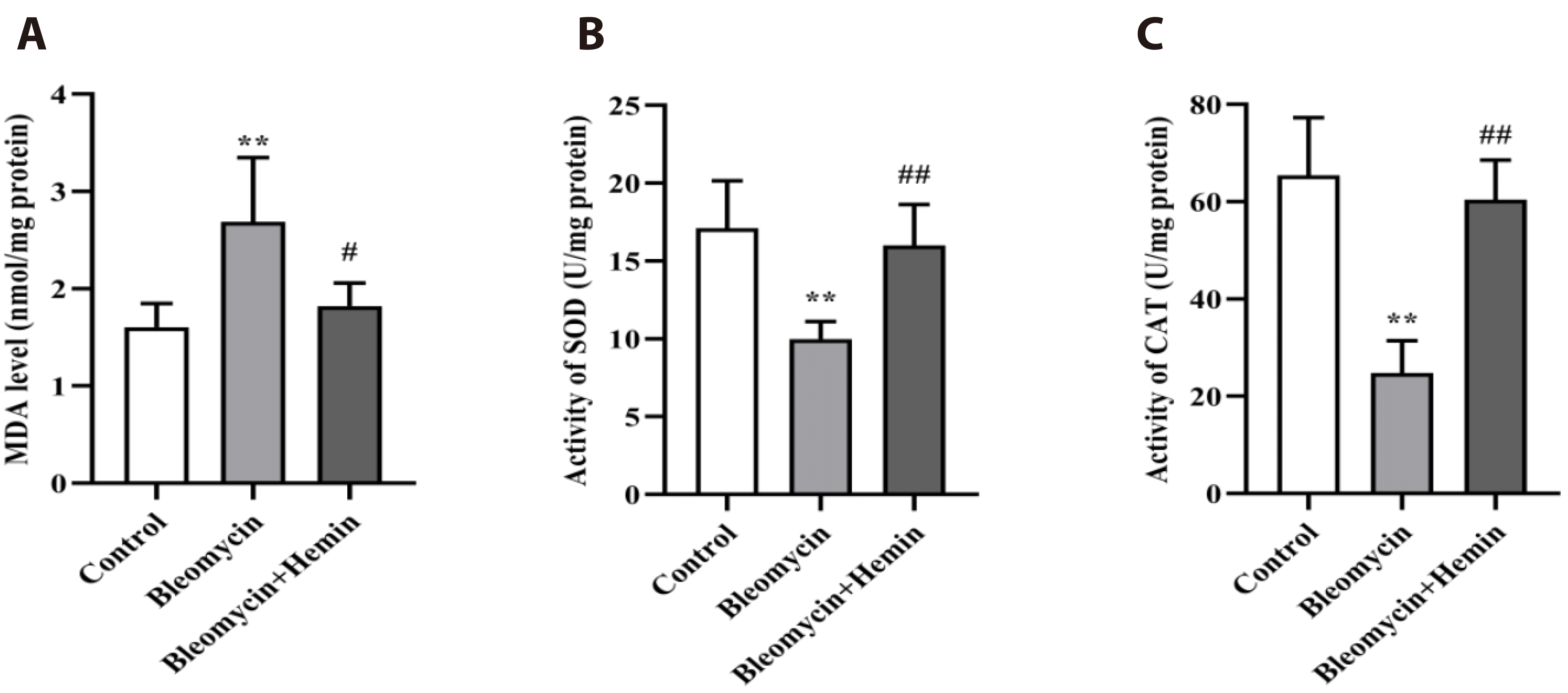
Effect of hemin on the MDA, SOD and CAT contents or activity in the lung tissue of mice with PF
MDA (p < 0.01) content was significantly increased, and SOD and CAT (p < 0.01) activity were markedly decreased in the lung tissue of the bleomycin group compared to those of the control group (Fig. 4). Hemin pretreatment significantly inhibited the bleomycin-induced increase in MDA (p < 0.05) production and the decrease in SOD and CAT (p < 0.01) activity.
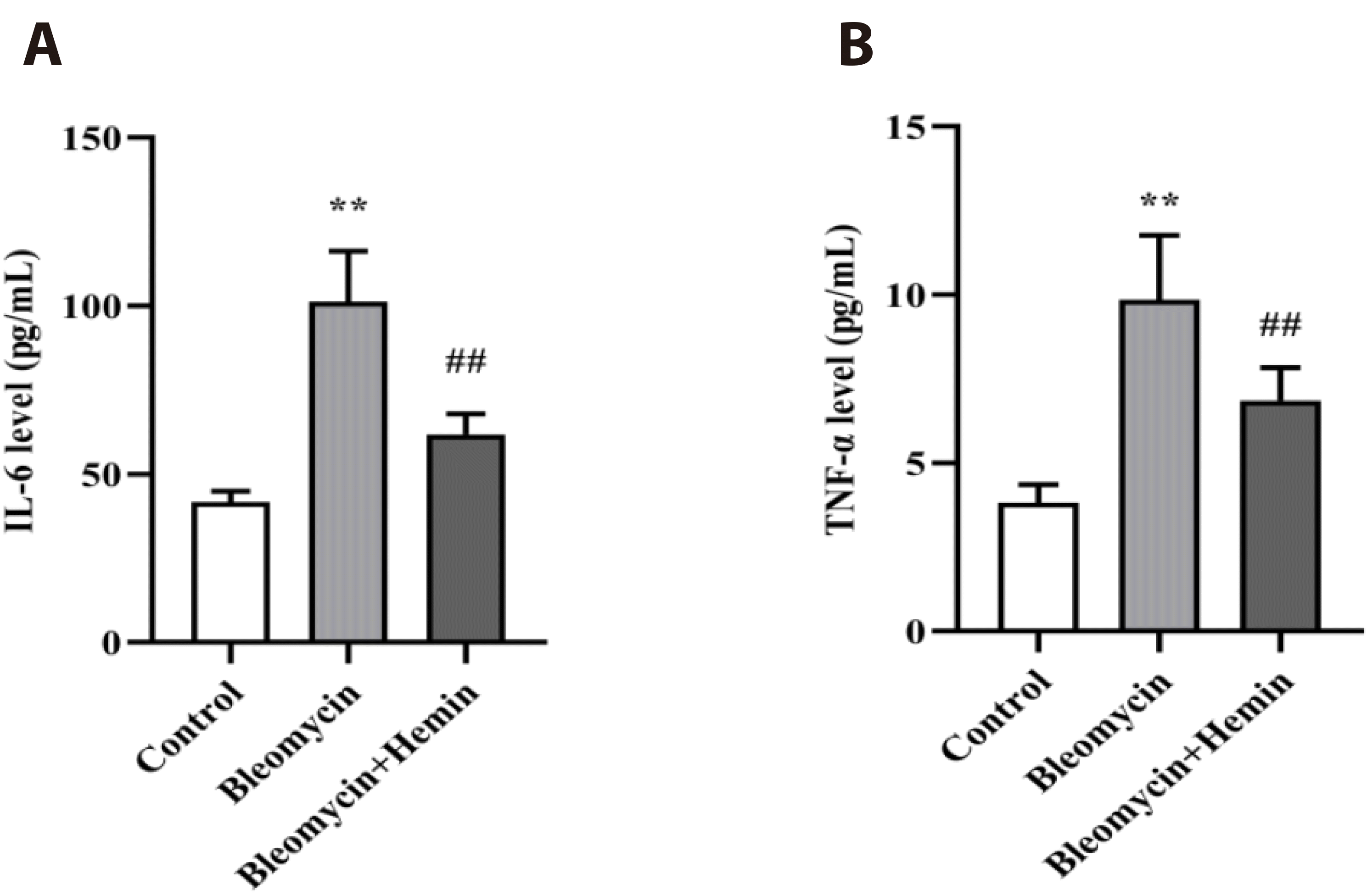
Effect of hemin on serum levels of IL-6 and TNF-α in mice with PF
As shown in Fig. 5, bleomycin stimulation significantly elevated the serum levels of IL-6 and TNF-α (p < 0.01) compared to those in the control group. The serum concentrations of IL-6 and TNF-α (p < 0.01) were markedly lower in the bleomycin + hemin group than in the bleomycin group.
Effect of hemin on the TGF-β1/MAPK pathway in lung tissue from mice with PF
Western blotting revealed that bleomycin stimulation significantly increased the expression of TGF-β1 (p < 0.01) and the phosphorylation of p38, ERK1/2 and JNK (p < 0.01) in mouse lung tissues compared with those in the control group. Hemin pretreatment markedly inhibited the bleomycin-induced increase in TGF-β1 expression (p < 0.05) and p38 (p < 0.05), ERK1/2 (p < 0.01) and JNK phosphorylation (p < 0.05) (Fig. 6 and Supplementary Fig. 1).
Effect of hemin on the AMPK/SIRT1/PGC-1α/HO-1/NF-κB pathway in lung tissue from mice with PF
As shown in Fig. 7 and Supplementary Fig. 2, the expression of SIRT1, PGC-1α and HO-1 (p < 0.01) and the phosphorylation level of AMPK (p < 0.01) in the lung tissues of the mice in the bleomycin group were significantly lower than those in the control group, while the phosphorylation level of NF-κB p65 (p < 0.01) was significantly greater than that in the control group. Hemin pretreatment partially increased SIRT1 (p < 0.05), PGC-1α (p < 0.01) and HO-1 (p < 0.01) expression and AMPK (p < 0.01) phosphorylation and decreased NF-κB p65 (p < 0.01) phosphorylation.
DISCUSSION
PF is a devastating chronic pulmonary disease [30]. Epidemiological studies have shown that the incidence of PF is approximately 0.9 to 13/100,000 [31]. Dove's study showed a 9.8% increase in PF mortality in the United States over the past 10 years [32]. The pathogenesis of PF is complex and diverse, existing therapeutic drugs have the disadvantages of poor efficacy and many side effects [33], and the search for safe and effective anti-PF drugs has become an important research topic for a wide range of scholars. Therefore, the aim of the present study was to investigate the protective role of hemin in bleomycin-induced PF and its possible mechanisms to provide a new option for the treatment of PF.
The bleomycin-induced PF model is widely used in experimental studies, and its pathological process can be divided into three phases: early (inflammatory), intermediate (pro-fibrotic) and late (fibrotic). Bleomycin may cause patchy parenchymal inflammation of variable intensity, epithelial cell damage with reactive hyperplasia, basement membrane damage, interstitial damage and intra-alveolar fibrosis by provoking DNA strand breaks and oxidative damage [34-36]. The present study showed that bleomycin stimulation resulted in a marked decrease in survival and terminal body weight; a significant increase in the lung index; substantial inflammatory cell infiltration in the alveolar lumen and interstitium; and increased collagen deposition in mice, in agreement with the findings of Yang et al. [37]. After hemin treatment, these changes were significantly improved, and the degree of PF was mitigated.
α-SMA is a marker of myofibroblast activation and accelerates the development of PF by participating in the synthesis of extracellular matrix components such as collagen I, collagen III and fibronectin. TGF-β1 is one of the most important pro-fibrotic proteins, and its downstream MAPK pathway, as a key signaling pathway in vivo, mediates cellular processes such as inflammation, fibrosis and cancer progression and regulates the expression of multiple inflammatory mediators and cytokines [15,38]. The MAPK pathway has been confirmed to be closely associated with lung fibroblast activation and extracellular matrix deposition [39,40]. Studies have shown that increased ROS levels lead to activation of the MAPK pathway, which is associated with PF [41,42]. Dong et al. [43] reported that in H2O2-induced human alveolar epithelial A549 cells, MDA production was excessive, and SOD and GSH-Px activities were reduced, accompanied by activation of the MAPK pathway, whereas Sarcodon aspratus reduced apoptosis and alleviated oxidative damage to cells by decreasing the phosphorylation levels of ERK, JNK and p38. Yang et al. [44] reported that berberine has a protective effect on bleomycin-induced PF, and the underlying mechanism may be related to inhibition of the p38 MAPKa (pT180/Y182) sites, reduction of inflammatory cell aggregation and cytokine secretion, attenuation of the inflammatory response, downregulation of TGF-β1 and α-SMA expression, reduction of collagen production and inhibition of the conversion of lung fibroblasts into myofibroblasts. The present study showed that there was a significant increase in MDA production and a marked decrease in SOD and CAT activity; moreover, the expression of TGF-β1 and the phosphorylation of p38, ERK1/2 and JNK were markedly elevated, and the expression of α-SMA and collagen I increased in the lung tissue of the bleomycin group mice. However, hemin pretreatment notably inhibited MDA overexpression, increased SOD and CAT activities, decreased the expression and phosphorylation levels of proteins related to the TGF-β1/MAPK pathway, and decreased the positive cellular expression of α-SMA and collagen I, suggesting that hemin probably ameliorated bleomycin-induced PF by inhibiting TGF-β1/MAPK pathway activation, attenuating oxidative damage and reducing extracellular matrix deposition.
AMPK is a recognized cellular bioenergetic sensor and metabolic regulator. AMPK protects the liver, heart, lung and kidney from fibrosis [45]. SIRT1 is a histone deacetylase that exerts anti-inflammatory effects by inhibiting the production of several proinflammatory cytokines by inducing the activation of PGC-1α and the downregulation of NF-κB [46,47]. PGC-1α is a primary regulator of mitochondrial biogenesis, oxidative phosphorylation and mitochondrial antioxidant defense and is responsible for maintaining a metabolic steady state. PGC-1α and NF-κB regulate each other during inflammation and play an important role in the vicious cycle of OS [48]. HO-1 is a membrane-bound enzyme encoded by the HMOX-1 gene and can be strongly induced by a variety of factors, including hemin, UV light and heat shock [49-53]. HO-1 acts both as a key cytoprotective molecule against oxidative damage and as a key anti-inflammatory protein that can attenuate NF-κB-mediated inflammatory responses [54,55]. The literature has shown that Sanghuangporus sanghuang acts by regulating the TLR4/NF-κB/MAPK, Keap1/Nrf2/HO-1, CaMKK/AMPK/Sirt1 and TGF-β/SMAD3 pathways and exerts anti-inflammatory, antioxidant and antiapoptotic effects, thereby relieving bleomycin-induced PF in mice [15]. Park et al. [56] reported that hemin prevents UV-induced skin cancer, and the underlying mechanism may be related to activation of the AMPK/HO-1 pathway. Ali et al. [57] reported that hemin ameliorates ovarian ischemia‒reperfusion injury by regulating the HO-1 and p-JNK/p-NF-κB p65/iNOS pathways, increasing antioxidant capacity and inhibiting the inflammatory response. In the present study, after bleomycin stimulation, the expression of SIRT1, PGC-1α and HO-1 and the phosphorylation of AMPK were significantly reduced, and the phosphorylation of NF-κB p65 and the serum IL-6 and TNF-α levels were significantly increased in mouse lung tissues; however, hemin pretreatment significantly inhibited the bleomycin stimulation-induced changes in these indices. These results suggest that hemin possibly inhibits OS and the inflammatory response by modulating the AMPK/SIRT1/PGC-1α/HO-1/NF-κB pathway, thereby ameliorating bleomycin-induced PF.
In conclusion, hemin likely alleviated bleomycin-induced PF by regulating expression and phosphorylation of proteins related to the TGF-β1/MAPK and AMPK/SIRT1/PGC-1α/HO-1/NF-κB pathways, inhibiting oxidative stress and the inflammatory response, and reducing extracellular matrix deposition; however, the exact mechanism involved requires further investigation.
SUPPLEMENTARY MATERIALS
Supplementary data including two figures can be found with this article online at https://doi.org/10.4196/kjpp.2024.28.6.559

 XML Download
XML Download