

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Hepatocellular carcinoma (HCC) is the third-greatest source of cancer mortality worldwide [1]. The main threats to liver cancer are alcohol, aflatoxins, hepatitis, pollutants, and industrial toxic chemicals [2]. The most important environmental carcinogen is diethylnitrosamine (DEN) which is dispersed in tobacco smoke, whisky, and processed meat [3]. DEN comprises of the N-nitrosamines broadly used in industries, which are hazardous to human beings as DNA adducts [4]. DEN is generally used as a carcinogen, and it forms neoplastic lesions and proliferation in the hepatic tissue of experimental animals, particularly rodents [5,6]. DEN exerts hepatic free radicals, which enhance the requirement for antioxidants, and consequently cause oxidative stress in HCC [7]. Oxidative stress and inflammation are the dual motivating forces behind HCC development [8]. So, a bioactive agent with potent antioxidant activity is required for the management of the early-stages of HCC.
Apoptosis in the liver is related to advanced inflammation and fibrosis [9]. The latest studies propose the regulation of proliferation and apoptosis involved in mitochondria through the P13K/AKT signaling pathway [10]. The conventional mode of radiotherapy and chemotherapy are found to be less effective in patients with liver cancer. It has been deliberated that chemoprevention is the paramount approach for reducing the harmful prognosis of liver carcinoma [11]. Anti-carcinogenic effects exerted by chemopreventive agents include preventing the creation of carcinogenic metabolites, detoxifying carcinogens, scavenging reactive oxygen species (ROS), restraining cell proliferation, and triggering apoptosis [12].
Medicinal plants are extensively used for bioactive compounds to cure several diseases, including liver cancer. Goniothalamin (GTN) is the unique genera of Annonaceae, with almost 160 species disseminated all over Southeast Asia [13]. GTN is a styryl-lactone that unveils assorted pharmacological actions comprising anti-inflammatory, anti-cancer, apoptotic, and immunosuppressive potential [14,15]. Conversely, little is known about how GTN might affect liver cancer that is linked to inflammation. To the best of our knowledge, no specific research on the HCC effects of GTN on the hepatic cancer has been published. In the current study, liver cancer was utilized to evaluate the liver's marker enzymes, antioxidant state, lipid peroxidation levels, and the expression of the PI3K/Akt signaling pathway in molecular cells that are proliferating. Additionally, we assess the chemopreventive, anticancer, and apoptotic mechanism of GTN via P13K/AKT signaling in DEN-induced HCC in rats.
Go to : 
METHODS
Chemicals
DEN and GTN were purchased from Gibco-Thermofischer Scientific. The cytokines (interleukin [IL]-1β, tumor necrosis factor-α [TNF-α], IL-6, and nuclear factor-κB [NF-κB]) levels in serum were investigated by IL-1α ELISA Kit purchased from MyBioSource. The specific antibodies for PCNA, Bcl-2, caspase-3, the primary antibodies of P13K, AKT, β-actin, and secondary antibodies, were purchased from Beyotime Biotechnology.
Animals
Male Wistar albino rats weighing between 150 to 190 gm (18–20 weeks of age) were selected for the study. They were kept under fixed laboratory temperature, humidity, and 12/12 h light-dark cycles. This study was permitted by the Xi'an Hospital of Traditional Chinese Medicine animal ethical committee, approved ethical number No. TC23329981.
Experimental design
The Wistar albino rats were distributed into 4 sets of n = 6. The dose of GTN was determined based on the literature [16]. Group 1 rats were the control, while group 2 rats were given 0.01% of DEN by imbibing water for 16 weeks to induce HCC. Group 3 rats were delivered 0.01% DEN and continued orally administered the GTN (30 mg/kg bw., dissolved in 10% dimethyl sulfoxide [DMSO]) for 16 weeks. Only GTN (30 mg/kg bw., dissolved in 10% DMSO) was administered to group 4. In the 16th week, the animals were euthanized by decapitation after overnight fasting. The blood and liver were preserved in ice-cold dishes for biochemical study.
The volume of the tumor was calculated as follows:
V = (4/3) π [D1/2] [D2/2] [D3/2], where D1, D2, and D3 are the diameters (millimeter; mm3) of lesions in mm. The tumor burden was calculated by multiplying tumor volume and tumor number per rats.
Preparation of tissue homogenate
The liver lobes were washed with saline and homogenized with suitable buffer (W/V), centrifuged for 20 min at 4°C. The supernatant was separated and used for biochemical analysis.
Assessment of incidence of tumor
To assess the incidence of tumor after the experimental period, the liver was detached and cleaned with PBS. Then it was dehydrated using filter paper and counted by macroscopic analysis.
Histopathological analysis of the liver
The liver tissues were fixed with 10% formaldehyde, followed by implanting in paraffin. Afterward, the liver tissues were dissected by a rotary microtome into 3–5 µm sections and successively stained with hematoxylin and eosin. Pathological changes were observed through a light microscope.
Assay of cancer markers
The hepatic enzyme markers aspartate aminotransferase (AST), alanine aminotransferase (ALT) [17], lactate dehydrogenase (LDH), alkaline phosphatase (ALP), and gamma glutamyltranspeptidase (γGT) were analyzed in serum by using the methods of Rosalki and Rau [18]. AST, ALT, LDH, ALP & γGT spectrophotometer Kits purchased from MyBioSource.
Immunohistochemical analysis
Immunohistochemical examinations of PCNA, Bcl-2, and caspase-3 were done in hepatic sections by the Super Sensitive Polymer-HRP Detection System and purchased from GBI Labs, as per the guidelines format.
Determination of cytokines in serum
The cytokines (IL-1β, TNF-α, IL-6, and NF-κB) levels in serum were investigated by an IL-1α ELISA Kit purchased from MyBioSource.
Western blot analysis
The P13K, p-P13K, p-AKT, and AKT expressions were evaluated by Western blot. In brief, the rat liver was crushed with lysis buffer and centrifuged for 5 min at 4°C. Separated from the supernatant and whole protein were estimated by the Bradford technique [24]. 20 µg of protein was electrophoresed by SDS-PAGE and translocated to the nitrocellulose membrane. Thereafter, 3% bovine serum albumin was treated to block the film for 2 h. Added primary antibodies and subsequently secondary antibodies for one hour. For the recognition of bands by smearing the chemiluminescence reagent, and subsequently, the blot densitometry assessment.
Statistical analysis
The Graphpad prism software version 8.0.1 was used to scrutinize the data. The data are expressed as mean ± SD. Statistical comparisons were performed by one-way analysis of variance (ANOVA), followed by Duncan’s multiple range test (DMRT) using SPSS version 17.0 for windows (SPSS Inc.). The data were analyzed using Kruskal–Wallis test. A value of p < 0.05 was considered statistically significant. The results are displayed as the mean ± SEM.
Go to :
RESULTS
Body weight and liver weight
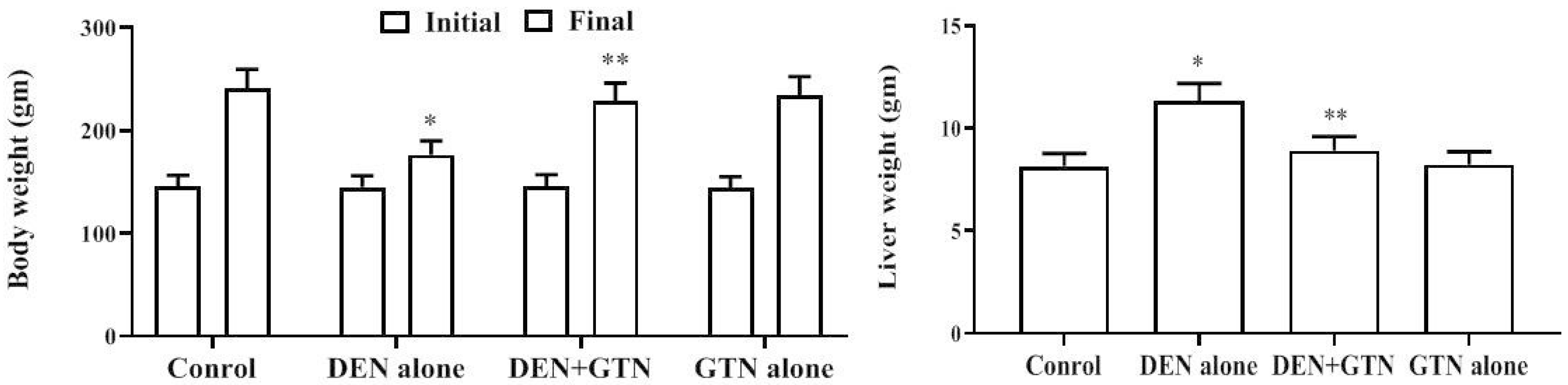
The rats did not show any marked difference in their diet or water intake during the entire experimental period. The body weight was considerably (p < 0.05) decreased in DEN-treated rats versus control rats. The liver weight was exhibited to be elevated remarkably (p < 0.05) in the DEN-induced group. Administration of GTN to the DEN treated rats markedly (p < 0.05) enhanced body weight, and liver weight whereas it reduced liver mass and relative liver mass (Fig. 1). There were no changes observed in the GTN alone treatment.
Gross appearance and occurrence of tumors
The obvious presence of liver tumors seen in rats treated with DEN alone (Fig. 2A). The DEN group displayed a high mean number of tumors per rat, 100% tutor incidence, and a large number of nodules. When GTN was administered, nodules (Fig. 2B), tumor incidence (Fig. 2C), tumor number (Fig. 2D) and the mean number of tumors (Fig. 2E) per rat were all significantly reduced (Table 1).
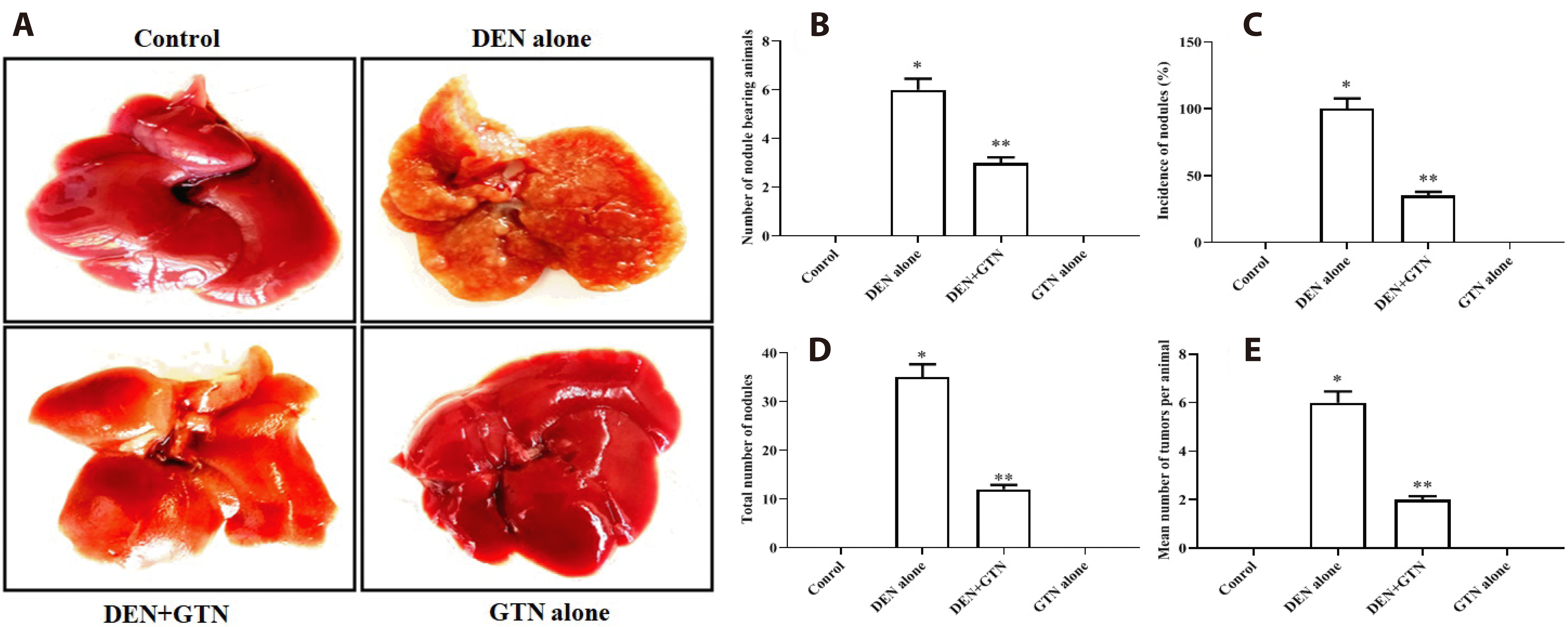 | Fig. 2Gross appearnae and tumour incidence of hepatic cancer in experiemental model.(A) Photomicrograph showing the gross appearance of liver tumor tissue at the 16th week of DEN-induced rats (n = 6). (B) Effect of GTN on the incidence of cancer nodules, number of nodule bearing animals/total number of animals. (C) Incidence of nodules (%) (incidence of nodules and number of nodules bearing rats/total number of rats in each group) × 100. (D) Total number of nodules. (E) Mean number of tumors per animal (total number of tumor/number of tumor bearing rats in each group). Data is represented as mean ± SD. At *,**p < 0.05, values that do not share a common superscript letter symbol differ statistically significant. GTN, goniothalamin; DEN, diethylnitrosamine.
|
Table 1
Effect of goniothalamin on the incidence of cancer nodules, total number and number of tumors of control and experimental rats
| Groups | No. of nodule bearing animals/total no. of animals |
Incidence of nodules (%)a |
Total number of nodules |
Mean no. of tumors per animalb |
|---|---|---|---|---|
| Group I | 0/6 | - | Nil | 0 |
| Group II | 6/6 | 100 | 35 | 6 |
| Group III | 3/6 | 35.11 | 12 | 2 |
| Group IV | 0/6 | - | Nil | 0 |
![]()
Histopathological analysis of liver
Histopathological alterations were detected in the hepatic tissues of the experimental rats (Fig. 3). Control rats exhibited normal liver histology with uniform nuclei. In the liver of the DEN-treated rats abnormal arrangements, irregular nuclei, and infiltration of fats, were observed. DEN together with GTN improved the cytosolic arrangement and could help in maintaining normal histology. GTN alone treated rats displayed similar to normal histology.
 | Fig. 3Histopathological modifications in the hepatic tissue of the control and trial rats.Control group showing normal liver histology. DEN-induced liver tissue showing nodules, vascular channels, and fat deposits. DEN + GTN treated rat liver tissue showing mild alterations. GTN treated rat showing normal histology of hepatic tissue. GTN, goniothalamin; DEN, diethylnitrosamine.
|
Liver marker enzymes in serum
Fig. 4 depict the toxicity enzymes and liver cancer marker in serum. The activity of ALT, ALP, AST, γGT, and LDH were remarkably high (p < 0.05) in DEN-induced rats in contrast to the control group. The hepatic enzyme levels reduced the enzyme levels in GTN-treated along with DEN. GTN alone treated group liver marker enzymes levels showed similar to normal rats.
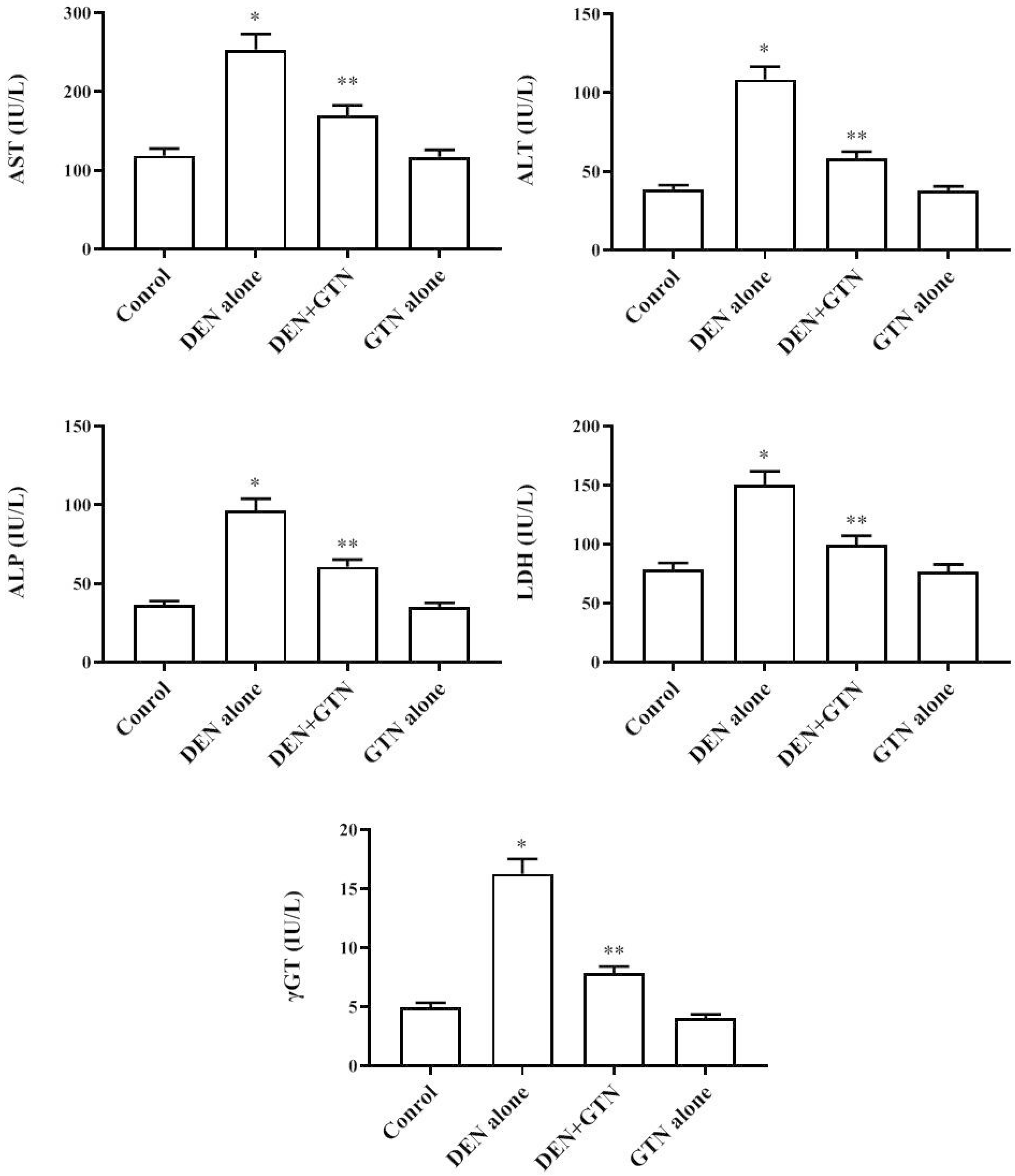 | Fig. 4Influence of GTN on liver cancer enzyme markers in serum on DEN-induced rats (n = 6).Data is represented as mean ± SD. Values not sharing common superscript letter differ significantly at *,**p < 0.05. GTN, goniothalamin; DEN, diethylnitrosamine; AST, aspartate aminotransferase; ALT, alanine aminotransferase; ALP, alkaline phosphatase; LDH, lactate dehydrogenase; γGT, gamma glutamyltranspeptidase.
|
Antioxidant enzyme activity of liver tissues
The antioxidant enzymes namely CAT, SOD, GPx, and GR were diminished substantially (p < 0.05) in DEN-induced HCC rats versus control (Fig. 5). Administration of GTN reversed the enzymatic antioxidant status to near normal. Normal antioxidant status was observed in GTN alone treated rats.
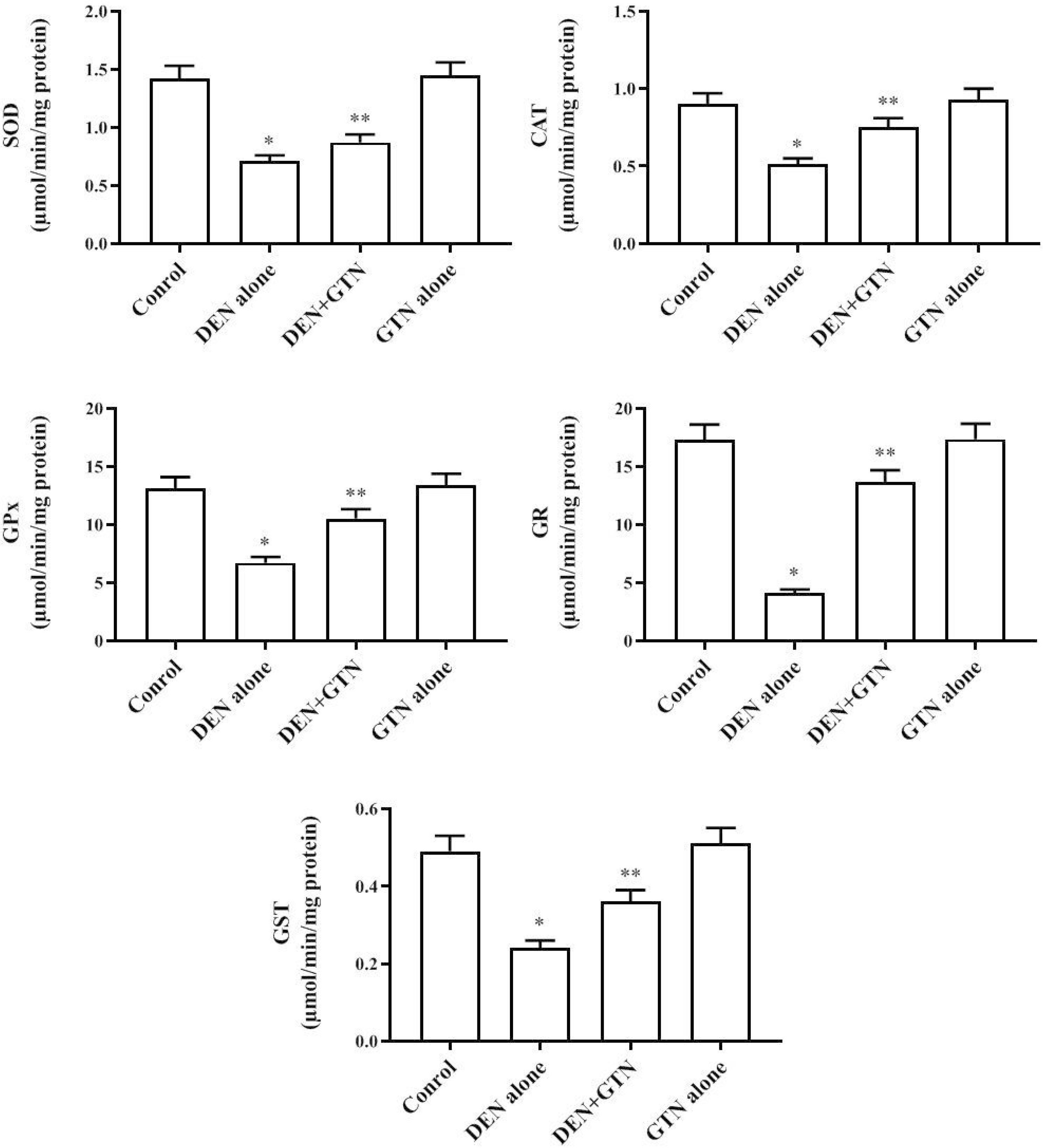 | Fig. 5Influence of GTN on anti-oxidant enzymes in liver tissues on DEN-prompted rats (n = 6).Results were depicted as mean ± SD. At *,**p < 0.05, values that do not share a common superscript letter symbol differ statistically significant. GTN, goniothalamin; DEN, diethylnitrosamine; SOD, superoxide dismutase; CAT, catalase; GPx, glutathione peroxidase; GR, glutathione reductase; GST, glutathione-S-transferase.
|
Influence of GTN on PCNA, Bcl-2 & caspase-3 expression
PCNA is an exceptional marker for cell propagation. In the control and GTN alone treated group was not expressed the PCNA antigen in the liver (Fig. 6A). Conversely, DEN-treated liver showed positive PCNA staining, which points towards vigorous proliferation of liver cells. Minor PCNA expression was observed in GTN together with DEN, revealing the anti-proliferative action of GTN against DEN-stimulated HCC. The protein level of Bcl-2 in the control group showed insignificant staining. Augmented expression of Bcl-2 protein was perceived in the DEN-treated group (Fig. 6B). The GTN-treated group showed a lesser amount of staining in liver cells. Very negligible staining was observed in the GTN-only-treated group. The protein expression of caspase-3 in the control group showed feeble hepatic staining (Fig. 6C). The DEN-persuaded group showed trivial staining. GTN, along with DEN, showed moderate hepatocyte staining. GTN-only-treated group showed weak hepatic staining.
 | Fig. 6Immunohistochemistry (IHC) staining patterns of cell proliferation.(A) PCNA and apoptosis (B) Bcl-2, and (C) caspase-3 protein expression in the control and experimental rats (n = 6). Scale bar = 50 μm. Arrow indigates the over expression of expression markers. GTN, goniothalamin; DEN, diethylnitrosamine.
|
Measurement of pro-inflammatory cytokines
The cytokines involved in inflammation, for instance IL-6, IL-1β, TNF-α, and NF-κB, were elevated in DEN-induced HCC rats (Fig. 7). Pro-inflammatory cytokines are released by DEN and emanate in the serum. These cytokine levels were diminished by GTN in DEN-induced rats. Normal cytokine levels were maintained in the GTN alone treatment group.
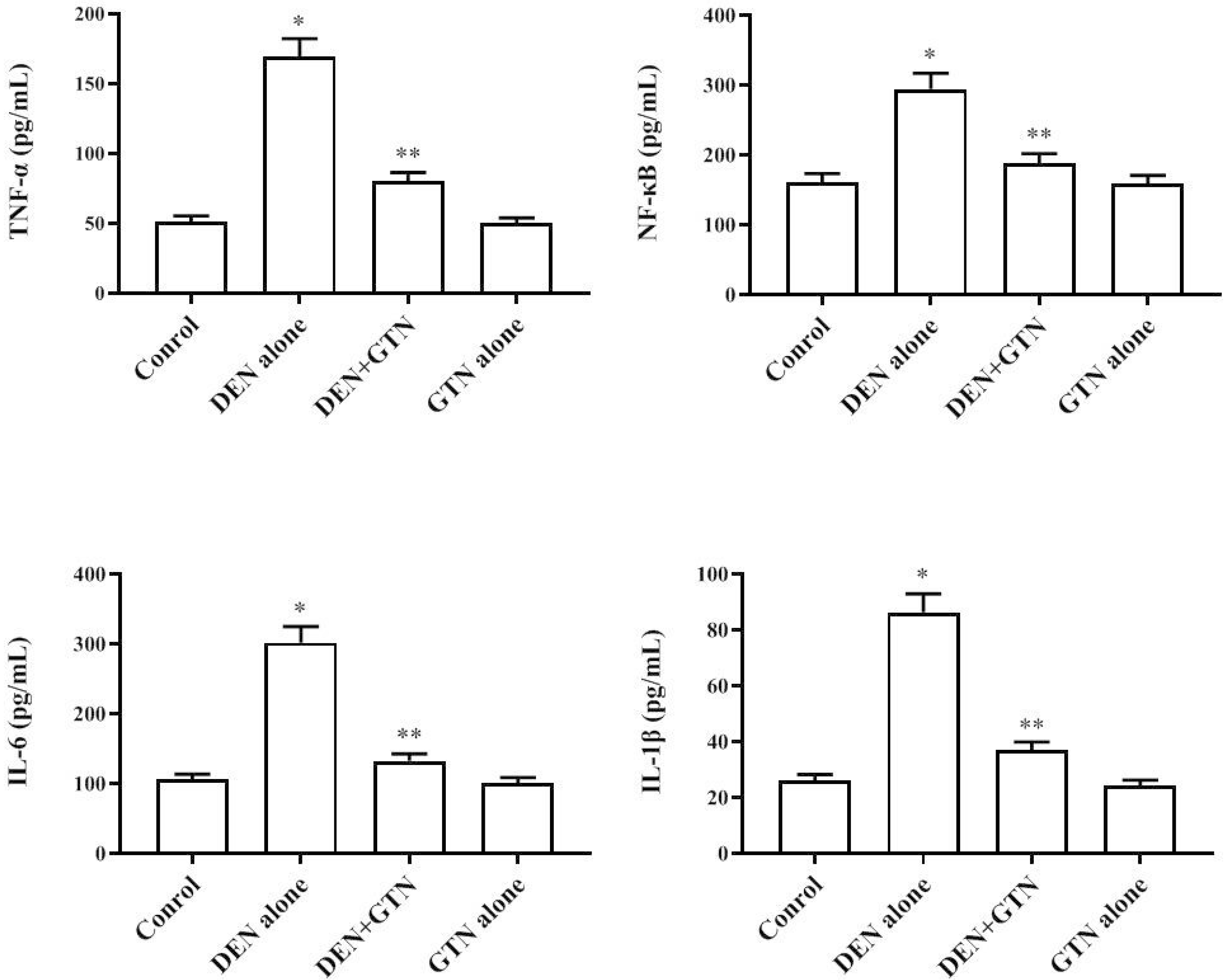 | Fig. 7Levels of cytokines in the serum control and experimental rats (n = 6).Results are displayed as mean ± SD. At *,**p < 0.05, values that do not share a common superscript letter symbol differ statistically significant. GTN, goniothalamin; DEN, diethylnitrosamine; IL, interleukin; TNF-α, tumor necrosis factor-α; NF-κB, nuclear factor-κB.
|
Effect of GTN on P13K/Akt pathway
p-PI3K, PI3K, p-Akt, and AKT levels in the apoptotic pathways have a very noticeable effect on the initiation of liver cancer (Fig. 8). DEN-treated rats revealed enhanced P13K and Akt levels compared to control group. Supplementation of GTN in the HCC rats dropped the P13K/Akt levels. The protein levels are similar in control and GTN-only-treated rats.
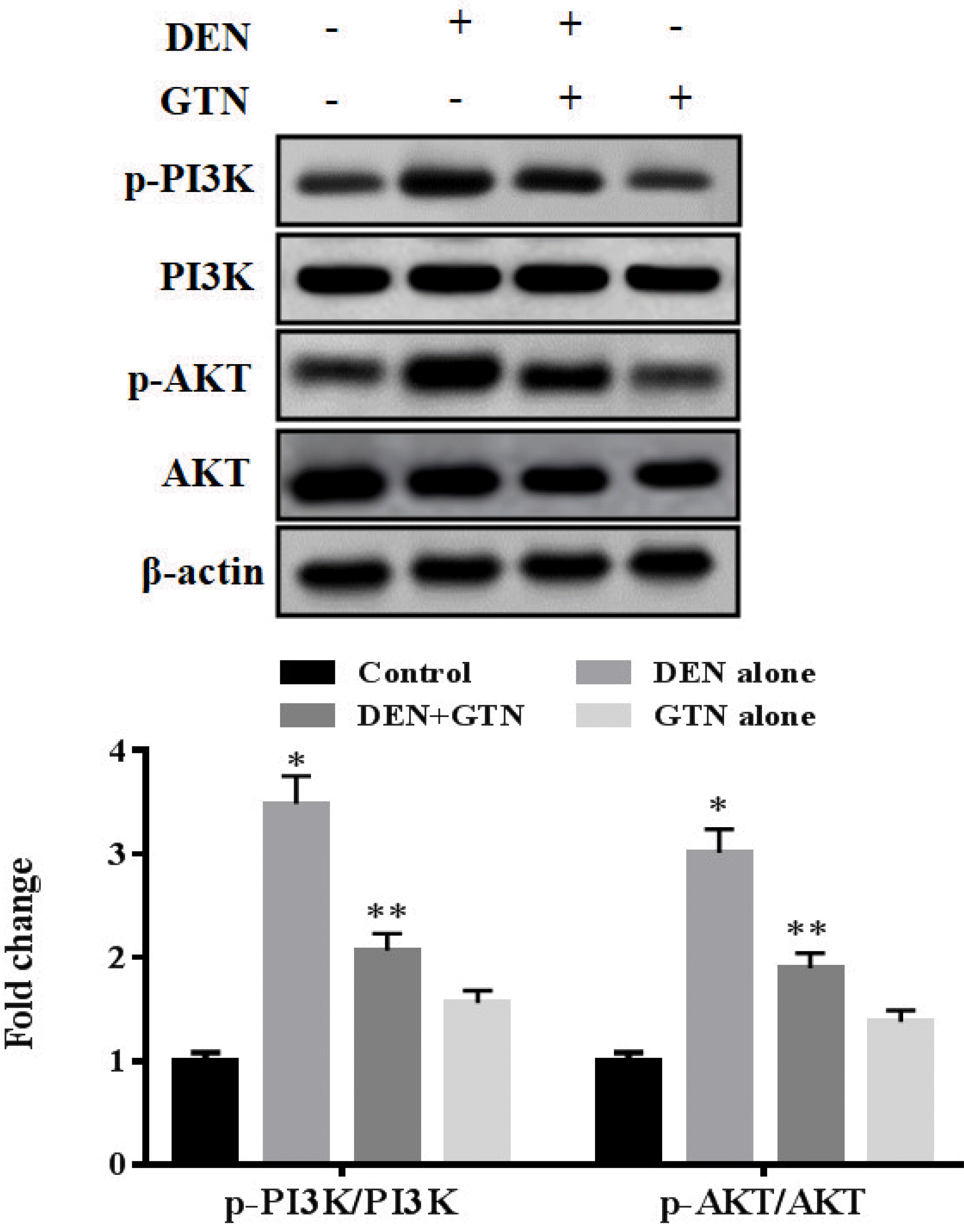 | Fig. 8Expression of PI3K, p-PI3K, AKT and p-AKT pathways in DEN-prompted HCC rats (n = 6).The concentrations of band were measured by densitometry and normalized to the β-actin internal control. Bars are expressed as mean ± SD. At *,**p < 0.05, values that do not share a common superscript letter symbol differ statistically significant. GTN, goniothalamin; DEN, diethylnitrosamine; HCC, hepatocellular carcinoma.
|
Go to :
DISCUSSION
Liver cancer is the primary cause of death around the world which is mostly associated with environmental factors [25]. DEN-induced HCC is an extensively recognized model for evaluating cancer initiation, replication, and necrosis [26]. The liver is known as a vital organ and is absorbed by xenobiotics and drugs due to its unique vascular system. Detoxifying enzymes present in the liver metabolize the xenobiotics, but may cause toxicity to other organs [27]. Chemopreventive efficacy of bioactive agents involved in the subdual of cell proliferation, genetic mutation, detoxification of carcinogens, immune system regulation, and orientation of apoptosis [28]. Natural products from medicinal plants are effectively used in hepatic disease. In the current study, we designed to evaluate the chemopreventive, anti-cancer, and apoptotic activity of GTN in HCC induced by DEN.
HCC is associated with weight reduction and liver cell damage. In this study, a significant diminution of body mass and an enlargement of liver mass were found during the trial period. These alterations were prevented by the GTN supplementation. We observed 100% tumor formation, total numbers of nodules, and tumor incidence were high in DEN-prompted rats. Treatment with GTN inhibited the formation, incidence, and volume of the tumor. Histopathological studies also confirmed the chemopreventive and anti-cancer effects of GTN. DEN treatment showed loss of normal liver histology, hyperchromatic, irregular nuclei, fat deposits, and neoplastic changes. GTN could reduce these histological changes and restorethe architecture of the liver.
Elevated activity of hepatic toxic enzyme markers such as AST, ALT, γ-GT, ALP, and LDH is reflective of hepatic failure [29]. Our results, too, were high-level toxicity and HCC marker enzymes observed in DEN-treated rats. These hepatic enzyme levels were decreased by the administration of GTN. Enhanced transaminase levels in serum indicate hepatic impairment and rupture of plasma membrane, which might dischargethe enzymes from the cytoplasm into the blood. It has been reported that LDH is a highly sensitive marker for solid tumors and is elevated in liver cancer [30]. Liver mutilation is reflective of the γ-GT level in serum [31]. Treatment for GTN lower the serum levels of these enzymes, reducing liver damage.
Oxidative stress aroused by ROS has been implicated with DEN-induced HCC [32]. Antioxidant enzymes (CAT, SOD, GPx, and GR) play a central role in stabilizing ROS. Our results revealed that DEN decreased the enzymatic antioxidant activity, whereas GTN could reverse it to near normal. This is in line with the previous report that enzymatic antioxidant activities are reduced in HCC [33]. GTN reversed the harmful effects of DEN via the perfection of antioxidant status and diminished oxidative stress.
Pro-inflammatory cytokines are pleiotropic hormones involved in the immune system that are related to tumor propagation and development [34]. Liver cancer is associated with IL-1β, IL6, TNF-α, and NF-κB. The existing study, IL-1β, IL6, TNF-α, and NF-κB levels were augmented by DEN, whereas they were reduced following GTN supplementation. Cell proliferation has a profound effect on cellular components linked with carcinogenesis [35]. Hence, the invention of bioactive agents that can alter the formation of neoplasm is essential for chemoprevention of HCC. An auxiliary protein, PCNA, is precisely involved in the replication of DNA [36]. The results showed that DEN treatment enhanced PCNA expression in the liver, denoting the high proliferation of cancer cells. PCNA expression is a known marker for liver cancer cell proliferation [37]. Attenuation of PCNA expression was perceived in the GTN treatment, revealing its protective action.
Apoptosis characterizes the controlled activation of programmed cell death, and the tumor cells’ apoptosis is a significant diagnostic tool [38,39]. Bcl-2 is known as an anti-apoptotic protein that inhibits cell death. Increased expression of Bcl-2 and diminished caspase-3 were found to be caused by DEN administration. GTN treatment attenuated Bcl-2 and elevated caspase-3 expression. Caspases are cysteine proteases that are formed in pro-active forms and stimulated in a protease cascade. Caspase-3 is the most crucial member among caspases and is responsible for the execution of apoptosis [40,41]. Administration of GTN suppressed P13K/AKT pathway in DEN-treated rats. DEN upregulated the protein levels of P13K and AKT in contrast to control. GTN down-regulated the protein levels of PI3K and p-AKT and thereby suppressed the P13K/AKT pathway. P13K/AKT signalling pathways act as a prominent actionin the progression and development of liver cancer [42]. In our study, we found GTN-mediated apoptosis via the inhibition of the P13K/AKT pathway.
GTN demonstrated significant protective effects in rats against DEN-induced HCC. GTN on apoptosis, inflammation, hepatoprotection, antioxidants, and cancer in rats with HCC. In light of our research, we draw the conclusion that GTN may reduce cell growth and promote apoptosis in DEN-induced hepatocellular cancer. These findings demonstrate GTN's chemopreventive properties in HCC.
Altogether, the results proved that GTN-administered DEN-treated rats experienced reduction in body weight, which was proved by histological improvements. Even the GTN reduced the hepatic toxicity and liver cancer markers and increased antioxidant levels in liver. Furthermore, in DEN-induced rats, GTN decreased the levels of PCNA and PI3K/AKT proliferation markers as well as the production of Bcl-2 and caspase-3 by pro-inflammatory cytokines IL-6, IL-1β, TNF-α, and NF-κB. Hence, we recommended GTN as a chemopreventive drug based on the overall findings.
Go to :
 XML Download
XML Download