

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Approximately 9.2% of middle-aged adults of a large cross-sectional sample from UK Biobank suffered from chronic neuropathic pain [1]. Reports from Canada and the USA also have indicated a greater prevalence of chronic peripheral neuropathic pain (17.9% and 15.7%, respectively) [2,3].
Chronic peripheral neuropathic pain is caused by damage to the nerve or its branches, which triggers a sequence of actions leading to the regeneration of new nerves. Inflamed nerve tissue activates the immune system, triggering regeneration and creating a new layer of myelin. This process results in nociceptor/peripheral sensitization, followed by ganglionic and central sensitization [4,5].
The gold standard therapy for chronic peripheral neuropathic pain involves prescribing antidepressants, gabapentinoids, or serotonin norepinephrine reuptake inhibitors [6]. Improvements in pain intensity and frequency, with a minimum improvement of 50%, indicate successful treatment [7]. However, chronic use of antidepressants (> 3 years) can have several side effects. According to Cartwright et al. [8], withdrawal symptoms (73.5%), sexual difficulties (71.8%), and weight gain (65.3%) are the most common side effects. Emotional adverse effects, such as addiction, feelings of emptiness, and suicidal ideation, have also been reported [8]. The current optimal therapy for chronic peripheral neuropathic pain should consider its cellular and molecular pathogenesis.
Methylcobalamin (MeCbl) is the active form of vitamin B12. MeCbl acts as a catalyst for methylation events throughout the body, including the methylation of the basic protein myelin, which forms the myelin sheets. MeCbl also induces Schwann cell differentiation, improves nerve conduction velocity (NCV), and boosts the secretion of brain-derived neurotropic factors (BDNFs), all of which contribute to axon regeneration [9]. MeCbl, apart from its role in nerve regeneration, can alleviate pain in neuropathic pain disorders. It has been approved as a neuropathic pain medication in Hong Kong, Abu Dhabi, and Australia, while not approved yet by US Food and Drug Administration [10]. Several clinical trials have been conducted to test its effectiveness in treating diabetic neuropathic pain [11–13], herpetic neuralgia therapy [14–17], chemotherapy-induced peripheral neuropathy [18], chronic pain post-thoracotomy [19], and glossopharyngeal neuralgia [20]. These trials have shown that treatment with MeCbl, whether used alone or in combination with other drugs or nerve stimulation, can significantly reduce pain intensity [11–17].
However, to the best of the authors’ knowledge, no detailed summary of the evidence regarding the molecular mechanism of action of MeCbl as a chronic peripheral neuropathic pain therapeutic agent is available. Therefore, we aimed to review the evidence regarding the molecular mechanism of MeCbl to identify its underlying molecular pharmacological action in neuropathic pain conditions. The main objective of this review was to reveal the potential of MeCbl as a chronic peripheral neuropathic pain therapeutic agent.
MAIN BODY
1. Pathogenesis of chronic peripheral neuropathic pain
Neuropathic pain arises from nerve damage, initiating a cascade of nerve regeneration processes involving inflammation, demyelination, neuroplasticity, and remyelination. The symptoms of heightened sensitivity linked to nerve injury are closely associated with prolonged neuroinflammation and impairments to remyelination [5,21]. The delay in optimal myelin sheath formation lengthens the internode distance of Ranvier over time. This causes disturbances during remyelination, resulting in increased electrical activity, abnormal sensitivity to various stimulants, and abnormal contact between adjacent axons resulting in ephaptic cross-talk [22].
Peripheral nerve injury triggers an inflammatory response involving various cells, such as Schwann cells, mast cells, macrophages, and T cells. Once the mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK-ERK) signaling pathway in Schwann cells is activated, inflammatory mediators that attract immune cells to the injured nerve are produced. Schwann cells also undergo dedifferentiation to initiate the degradation of the myelin sheath, which is necessary for nerve regeneration. Mast cells release inflammatory mediators that sensitize nociceptors and recruit neutrophils to the injury site. Neutrophils sensitize nociceptors, recruit macrophages and T cells, and are present shortly after the injury. Resident and circulatory macrophages are involved in phagocytosis and can secrete proinflammatory cytokines/chemokines. T-helper cells release proinflammatory and anti-inflammatory cytokines. Sensory nerve endings release neuropeptides that further contribute to the inflammatory response. Changes in the local chemical environment affect not only the injured nerve fibers but also healthy ones, leading to hyperexcitability and neuropathic pain [5,23].
Peripheral sensitization occurs alongside inflammation after nerve injury. When cells are injured, they release molecular signals that cause upregulation of certain proteins such as interleukin (IL)-1β, tumor necrosis factor (TNF)-α, and IL-6. Inflammatory mediators such as prostaglandin E2, bradykinin, serotonin, and H+ ions then bind to receptors on nerve cell membranes, leading to the activation of protein kinases A and C. These proteins phosphorylate ion channels involved in nociceptors, including the transient receptor potential cation channel subfamily V members 1 and 2 (TRPV1 and TRPV2, respectively), and voltage-gated sodium channel (VGSC). Consequently, the threshold for activating nociceptors decreases, and the excitability of the peripheral nerve terminal cell membrane increases. These findings suggest that peripheral sensitization is closely linked to the inflammatory response and involves the modulation of ion channels in nerve cells. Understanding these processes can lead to the development of treatment strategies for pain [21].
After nerve injury, axon remyelination begins with the proliferation and migration of Schwann cells. These cells guide the direction of axon growth to form the band of Büngner. The migration of Schwann cells is facilitated by the interaction between laminin and neuregulin-1 (NRG-1). Within the band of Büngner, Schwann cells secrete various growth factors and adhesion molecules that aid in remyelination. These include neurotrophins, BDNFs, neurotrophins 3, 4/5, and 6 (NT-3, NT-4/5, and NT-6), insulin-like growth factor (IGF-1), cilliary neurotrophic factor, glial-derived neurotrophic factor, neural cell adhesion molecule, N-cadherin, and integrins. The repair of axon and Schwann cell interactions is crucial for successful remyelination. NRG-1 and 3, IGF-1, and matrix metalloproteinase-9 influence the growth of myelin thickness and the distance between Ranvier’s nodes. If remyelination is disrupted, symptoms of neuropathic pain occur [24].
2. Chemical and physical structure of MeCbl
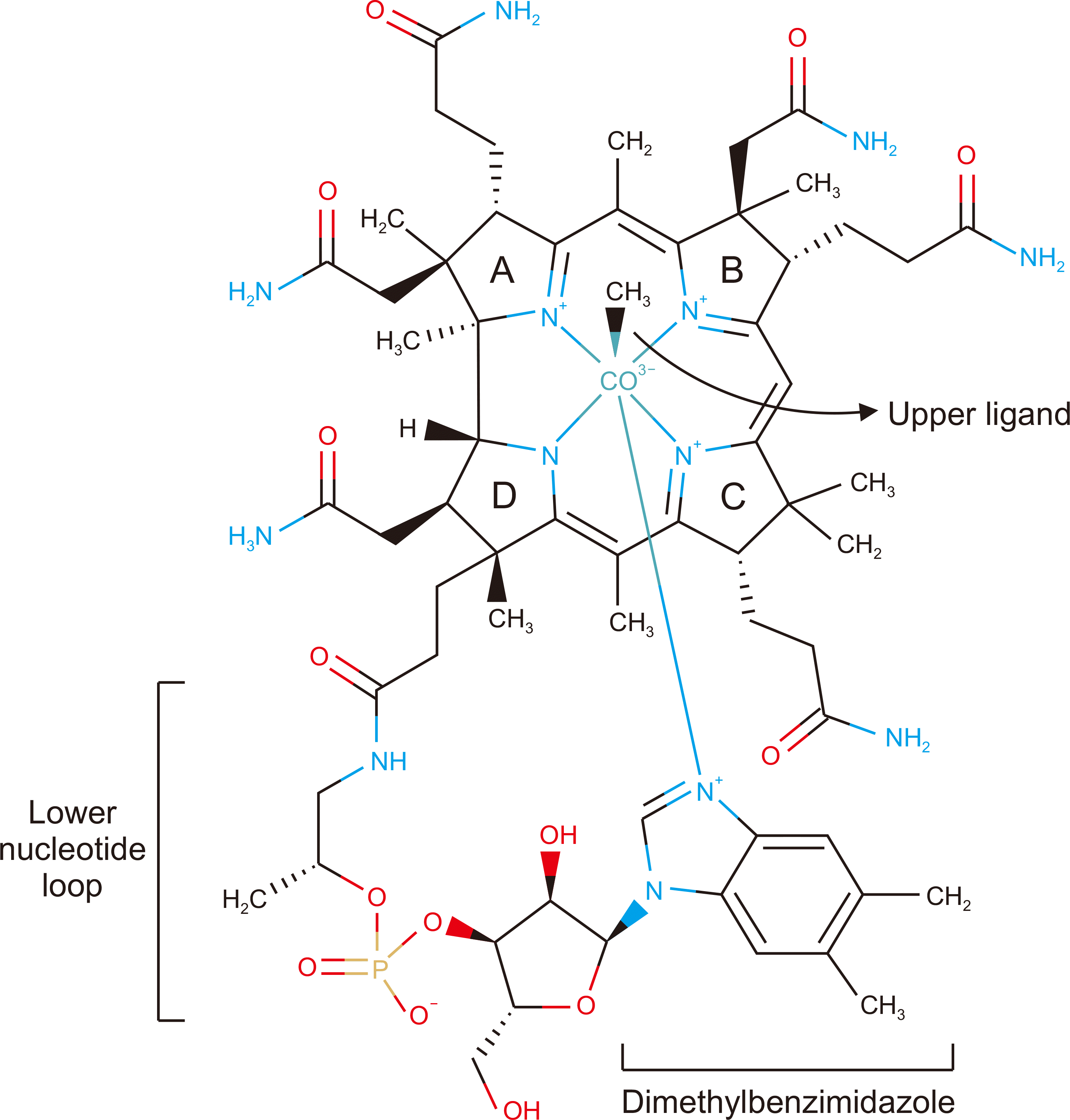
Vitamin B12, or cobalamin, is a water-soluble vitamin that cannot be synthesized independently by the human body. The chemical structure of vitamin B12 comprises a cobalt ion as the center of the structure, which binds to four nitrogen ions to form a corrin ring (Fig. 1) [25]. The lower ligand of the cobalt ion (beta) binds to the nitrogen ion of the dimethylbenzimidazole molecule. The upper ligand (alpha) of the cobalt ion binds to different groups resulting in four analogs of cobalamin, namely, cyanocobalamin, MeCbl, adenosylcobalamin (AdoCbl), and hydroxycobalamin (OHCbl) [26,27].
3. Pharmacokinetics and pharmacodynamics of MeCbl
1) Absorption of ingested MeCbl
Vitamin B12, whether ingested or taken orally, passes through numerous phases before reaching the target cells. Various binding proteins, including haptocorrin (HC/transcobalamin I [TC-I]), and intrinsic factor (IF) are involved in the initial binding and absorption of various forms of vitamin B12, including MeCbl. HC molecules are present in the saliva, and upper gastrointestinal system, where they initially bind to MeCbl. The cobalamin–HC complexes are degraded in the ileum, and the resulting free cobalamin molecules are bound by IF protein molecules released by gastric parietal cells. The IF–cobalamin complexes bind to the apical region of intestinal epithelial cells via the cubam receptor, allowing them to be endocytosed into the cells’ lysosomes for degradation. The liberated free cobalamin molecules enter the cytosol and blood circulation via multidrug resistance protein 1 or passive transport [27,28]. Because of its hydrophobic characteristics, MeCbl can enter the cell via passive diffusion, enhancing its conversion in the cell [29]. Hanawa et al. [30] demonstrated that because of its high molecular weight (1,344.4 g/mol), MeCbl cannot penetrate the choclea barrier but can be detected in the gracii muscle.
2) MeCbl in blood circulation
TC-I, transcobalamin II (TC-II), and transcobalamin III (TC-III) are three proteins that play a role in the distribution of cobalamin in the bloodstream. Once it enters the blood circulation, approximately 75%–90% of circulating MeCbl binds to non-specific TC-I, whereas the remainder binds to TC-II with a high affinity to enter the cells [27,28]. TC-III tends to remove cobalamin analogs from the tissues and bloodstream, convey them to the liver for elimination in bile [31].
3) MeCbl enters the cell
The TC-I–MeCbl complexes bind to nonspecific, multifunctional receptors on the cell surface, such as asialoglycoprotein receptors and hepatic-specific proteins, to enter liver cells for storage. On the contrary, only TC-II–MeCbl complexes can enter the peripheral cells via the specific TC-II receptor (CD 320) or passive diffusion because of their hydrophobic nature [29]. MeCbl uptake in neuronal cells is higher than in other cells because megalin receptors are present on the surface of neurons. These receptors are susceptible to TC-II. Megalin receptor activation is controlled by NRG-1 expression via the PI3-Kinase pathway [32].
4) MeCbl inside the targeted cell
Once inside the cell, the TC–MeCbl complex undergoes degradation in lysosomes, releasing free MeCbl into the cytosol using membranes called ABCD4 (cblf) and LMBD1 (cblF). MeCbl reacts with MMACHC (CblC protein), resulting in the release of methyl groups through dealkylation. Subsequently, the free cobalamin is converted into intracellular MeCbl to perform its functions [27,28]. The transformation of cobalamin to intracellular MeCbl in the cytosol requires a methyl donor from the reaction of tetrahydrofolate to 5-methyltetrahydrofolate and is affected by methionine synthase. In neurons, enhanced intracellular MeCbl conversion occurs because of increased glutathione formation due to elevated cysteine uptake triggered by excitatory amino acid transporters activation due to NRG-1 expression [33].
5) Excretion of MeCbl
From the ingested MeCbl, the liver stores 0.5–5.0 µg of the vitamin daily. This stored vitamin is then secreted in the bile and reabsorbed by the body, primarily in ileal enterocytes. Any unabsorbed MeCbl from food or bile is primarily excreted in feces, leading to an estimated daily loss of 0.1% of the body’s reserves. Excess MeCbl in the blood, such as after an injection, is eliminated in the urine [34].
Genetic abnormalities affect the activity of enzymes involved in absorption and the binding capacity of vitamin B12 to binding proteins during distribution and/or proteins involved in intercellular metabolism [35]. Mutations of aspartate 757 or serine 810 decrease the reactivity of the MeCbl cofactor to the binding site of methionine synthase [36].
4. Cellular and molecular action of MeCbl
Cobalamin present in the cell is converted into two coenzymes, namely, MeCbl and AdoCbl. MeCbl acts as a catalyst in methylation reactions throughout the body. These conversions ensure that cobalamin can perform its vital roles as a coenzyme in various enzymatic reactions [37,38]. MeCbl acts as a methyl donor for methionine synthase, which is important for the conversion of homocysteine to methionine. Methionine is then converted to S-adenosyl methionine, a key methyl donor in various methylation reactions in the body [9].
Cobalamin and its various analog forms help regulate acute and chronic inflammatory conditions by directly controlling the secretion of inflammatory cytokines, such as TNF-α, IL-6, epidermal growth factor, and neuronal growth factor. Cobalamin can also modulate immune reactions through its effects on T lymphocyte and natural killer cell activity [39,40].
Table 1 summarizes research conducted on MeCbl’s cellular and molecular function in various types of cells. A study on Plasmodium falciparum revealed that MeCbl exerts an antimalarial effect. Chemaly et al. [41] found that the 5,6-dimethylbenzimidazole side chain inhibited the production of β-haematin. Weinberg et al. [42] demonstrated that MeCbl exerts a mild inhibitory activity on nitric oxide synthases (NOS); however, light activation converts MeCbl into OHCbl, which has the strongest NOS inhibition action. MeCbl is beneficial in treating neurodegenerative illnesses involving tau aggregation, such as Alzheimer’s disease. MeCbl binds and caps the cysteine residues of tau protein, thereby preventing fibrillation and aggregation [43].
Combining MeCbl and AdoCbl significantly increased trophoblast cell proliferation and viability by upregulating EGFr mRNA expression, downregulating TNF-α mRNA expression, and decreasing homocysteine levels and oxidative stress at normal and supraphysiological concentrations of folic acid [44]. In silico experiments revealed that exogenous MeCbl may decrease viral replication of SARS-Cov-2 by inhibiting the RNA-dependent-RNA polymerase activity of SCV2-nsp12 enzyme [45].
5. Molecular pharmacological action of MeCbl as a chronic peripheral neuropathic pain therapy
Table 2 presents the molecular pharmacological efficacy and pain recovery after MeCbl treatment in diverse chronic peripheral neuropathic pain models by modifying inflammatory conditions, peripheral and central sensitization, and nerve fiber remyelination. A single MeCbl injection before and during the generation of neuropathic pain owing to cancer therapy with vincristine sulfate decreased thermal hyperalgesia and nerve damage. The reduction in nerve damage is attributed to decreased loss of intra-epidermal nerve fibers and the reduction of atypical mitochondria. Observations on the spinal dorsal horn revealed suppression of the NF-κB pathway, decreased activity of nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (NOXs), decreased TNF-α, and increased IL-10. The study suggested that the antinociceptive effect of MeCbl is mediated by a decrease in NOXs activity; however, how MeCbl can reduce NOXs activity is not known yet [46].
MeCbl can alleviate low back pain and lumbar stenosis syndrome caused by prolonged compression of the dorsal root ganglion (DRG). MeCbl administered intraperitoneally at 2.5 mg/kg daily for 28 days alleviated mechanical allodynia symptoms by lowering spontaneous activity in type A DRG neurons and hyperpolarization-induced ion channel activity in medium-sized DRG neurons [47].
MeCbl (100 µg) injected twice a week for 14 weeks in a rat model of neuropathy reduced VGSC expression and appearance, resulting in decreased mechanical allodynia and threshold value for mechanical stimulation [48,49].
Oral treatment with MeCbl at a dose of 15 μg/kg twice a day for 21 days alleviated neuropathic pain symptoms, such as heat, cold, and mechanical hyperalgesia. In addition, the levels of several nociceptor i.e.: transient receptor potential ankyrin 1 (TRPA1), transient receptor potential cation channel subfamily M member 8 (TRPM8), and phosphorylated p38MAPK were decreased in the DRG, whereas the levels of TRPV1 and TRPV4 were not significantly altered compared with those in normal conditions. Lower serum levels of IL-1β, IL-6, and TNF-α were associated with neuropathic pain symptoms [50].
MeCbl also affects neurite outgrowth and neuronal cell survival. It is thought to cause methylation of Ras proteins (H-Ras, K-Ras4A and 4B, and N-Ras), which activate ERK and PI3K/Akt proteins. Phosphorylated ERK and Akt can activate mTOR proteins, resulting in neurite outgrowth and neuronal survival [51].
MeCbl promotes Schwann cell development by blocking the Erk1/2 pathway, which increases myelin basic protein (MBP) and lipid synthesis. Lipid and MBP syntheses are needed to increase the thickness and density of the myelin sheath during remyelination [52].
In an end-to-side neurorrhaphy model of the damaged musculocutaneous nerve, MeCbl accelerates the development of the restored motor units of the injured nerve, resulting in greater conduction velocity and better synchronization when activated. MeCbl increases neuron-specific cytoskeleton molecule βIII tubulin expression, promoting axonal sprout growth in the receiving nerve. MeCbl also increases the activity of Schwann cells, which are responsible for most axonal degeneration-associated debris removal [53]. A study on the molecular action of MeCbl and chronic peripheral neuropathic pain showed that MeCbl has various abilities within the targeted cell to control the neuroinflammatory condition that affects sensitization after nerve injury (Fig. 2).
6. Clinical application of MeCbl in peripheral neuropathy
1) Methycobalamin for diabetic peripheral neuropathy (DPN)
Clinical trials have been conducted to study the use of MeCbl in treating DPN, either on its own or in combination with other treatments [11,12]. A meta-analysis of clinical trials recommended the oral dose of MeCbl is 500 µg three times per day, or 500–1,000 µg via intramuscular injection three times per week. The usage of MeCbl for 1 to 24 weeks is safe [54]. Sil et al. [55] concluded that an intramuscular injection of 500 µg MeCbl three times per week is more effective in elevating the serum cobalamin levels than a single intramuscular injection of an ultra-high 1,500 µg MeCbl dose. While the meta-analysis found no reduction in NCV, MeCbl effectively decreased DPN pain scores, including the neuropathic disability score and the neuropathic total symptom score. MeCbl, a water-soluble vitamin, is relatively safe [54].
2) MeCbl for herpetic neuralgia
MeCbl has been clinically tested as a treatment for acute or subacute herpetic neuralgia and ophthalmic herpetic neuralgia [14–17]. In subacute herpetic neuralgia, local subcutaneous administration of MeCbl (1,000 µg 5–6 times per week for 6 days to 4 weeks) has shown promising results in reducing pain and improving daily activities and quality of life [15]. Local injection of 1% lidocaine combined with 1,000 µg MeCbl effectively alleviated the ophthalmic and acute herpetic neuralgia symptoms [14,16,17]. MeCbl administration within 4–7 days after onset reduced the incidence of post-herpetic neuralgia [14]. The adverse effects encountered during clinical trials were relatively mild to moderate in the injection area, such as redness, bleeding (stopping within 1 minute), and bruises (disappearing after 7 days) [14–17].
CONCLUSIONS
Chronic peripheral neuropathic pain is closely associated with chronic neuroinflammation caused by nerve injury and impediments to remyelination. MeCbl is a potential therapeutic agent that targets this fundamental aspect of pathophysiology, offering improved treatment options with enhanced therapeutic outcomes while minimizing adverse effects. MeCbl reduces inflammation in the damaged nerve region by regulating NFκB activity in immune cells and neurons, which results in the reduction of TNF-α, IL-1β, and IL-6 levels and an increase in IL-10 levels. MeCbl also controls peripheral and ganglionic sensitization, which affects nerve impulse transmission by inhibiting the ion channel activation in neurons. In addition, it modulates remyelination, including axonal development and myelin sheath thickness. It regulates neurite outgrowth and neuronal cell survival by activating mTOR proteins. Furthermore, MeCbl increases the thickness and density of the myelin sheath by blocking the Erk1/2 pathway in Schwann cells, which results in enhanced lipid and MBP syntheses.
 XML Download
XML Download