

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Mosquito-borne viruses can be transmitted to humans and animals by infected mosquitoes and some of those viruses are potential to cause widespread epidemics in humans, domestic and wild animals (1, 2). They include the families of Bunyaviridae, Flaviviridae, Reoviridae, Rhabdoviridae, and Togaviridae (3). Mosquitoes which can transmit several pathogens to human mainly belong to three genera Anopheles, Aedes and Culex of the family Culicidae (4). Mosquito-borne arboviruses are usually restricted to specific habitats of their vectors and are seasonally abundant with transmission corresponding to the abundance of competent vector species (1).
The burden of mosquito-borne diseases is highest in the tropics and subtropics around the world (5). Several mosquito- borne viral diseases have been reported, of which human epidemic mosquito-borne arboviruses have emerged in both hemisphere during recent centuries including yellow fever virus (YFV), four serotypes of dengue viruses (DENV), West Nile virus (WNV), chikungunya virus (CHIKV), Japanese encephalitis virus (JEV) and Zika virus (ZIKV) (1, 6).
Vietnam is located in the tropical zone in Southeast Asia where the climate is characterized by high temperature and humidity all year round, the average temperature varies and gradually increases from the North to the South. Incomes historically dependent on agriculture when agricultural sector makes up one-third of the nation’s economy and livestock represents the second biggest contribution to household incomes. However, beside the perceived food and economic benefits, livestock production let the country faced with challenges such as the emergence and re-emergence of zoonotic diseases involving DENV, JEV, and ZIKV. DENV is the most widespread while all DENV serotypes circulate and regularly transmit in urban and semi-urban areas yearly, epidemics caused by this virus occurred in 1963, 1969, 1987 and the biggest outbreak occurred in 2022 with 367,729 cases (7, 8, 9). JEV has the potential to rapidly emerge in urban communities when pig-keeping is frequently occurring in peri-urban and urban areas of Vietnam. ZIKV seems to have recently re-emerged only in urban areas of the Central, Central Highlands and Southern regions, especially in Ho Chi Minh city.
Understanding information about the circulation of viruses and their vectors is very important. Monitoring the circulation of viruses and their vectors in Vietnam has not been conducted in a large number of studies to date. In 2005, Akabane virus, Oya virus, Getah virus, and Sagiyama virus were first isolated from mosquitoes collected in four northern provinces of Vietnam by Bryant et al (10). In 2005, Nabeshima et al isolated Banna virus from mosquitoes collected in 2002 in Ha Tay and Quang Binh provinces, they are located in the Northern and Central Coast regions of Vietnam (11). While in the southern region, virus isolation from mosquitoes has not been reported. In order to understand more about the circulation of viruses transmitted by mosquitoes in Vietnam, research on virus isolation from mosquitoes collected in An Giang Province was carried out.
MATERIALS AND METHODS
Mosquito collection
In three districts of An Giang province, mosquitoes were collected every month from June to December 2023. In each district, there were two collection sites. During each collection period, mosquitoes were collected within 30 minutes in each site, from 18:30 to 19:00 by using sweep nets near human residential areas and animal shelters (cowsheds and swinery). After each net, the mosquitoes were stored in a net cage, different cages will be allocated for the preservation of mosquitoes at each collection area. Blood-feeding mosquitoes were kept in cages for a few days to let them digest the blood in their guts, then they were identified based on their morphology by using “Illustrated key to the mosquitoes of Vietnam” (12). After identification, mosquitoes were sorted into pools with a maximum of 50 adults per pool according to mosquito species, placed and time, and kept at −80°C until assayed for virus.
Cell culture
Mammalian cell lines, Vero African green monkey kidney cells (Vero ATCC CCL81) and mosquito cell lines, Aedes albopictus C6/36 cells were used for virus isolation. The cells were cultured in Dulbecco’s modified eagle’s medium containing heat-inactivated 10% fetal bovine serum and 1% penicillin and streptomycin. Mosquito cell lines were maintained at 28°C under 5% CO2 while mammalian cell lines were maintained at 37°C in 5% CO2.
Virus isolation
Each mosquito pool was homogenized in 500 µl of ice-cold DMEM containing 2% heat-inactivated FBS and 1% penicillin and streptomycin. The suspensions were clarified by centrifugation (21,500×g for 5 min at 4°C), and the supernatants were passed through sterile 0.45-µm filters (Corning Inc., Corning, NY, USA). Filtrated supernatants of each pool were inoculated into duplicate wells of C6/36 and Vero cells in 24-well culture plates. These plates were incubated for 2 hours to allow virus adsorption. After adding fresh medium, cell cultures were incubated at cell culture conditions and observed daily for the presence of the cytopathic effect (CPE). Supernatants were collected from cells with CPEs. If no CPE was observed, after the fifth blind passage, culture supernatants were collected even if no CPEs were observed. These culture supernatants were stored at −80°C.
Detection of viral genome
Each culture supernatant from cells with CPEs was extracted RNA by using a QIAamp Viral RNA Mini Kit (QIAGEN, Inc, Valencia, CA, USA) according to the manufacturer’s recommended protocol. By contrast, culture supernatants from cells without any CPEs were pooled for RNA extraction. If RNA from mixed pools yielded was positive RT-PCR results, then RNA was again extracted from the supernatant in each pool and analyzed individually. To detect viral genes, RT-PCR was conducted using the OneStep RT-PCR kit (QIAGEN) with four universal primer sets (Table 1). RT-PCR for each primer set were performed at 50°C for 30 min (reverse transcription), 95°C for 15 min, followed by 35 cycles of 94°C for 30 sec, 55°C for 30 sec, and 72°C for 1 min (amplification), and final extension at 72°C for 10 min. 2% agarose gels were used to separate PCR products by electrophoresis, and intensive bands were purified by using the MinElute Gel Extraction Kit (QIAGEN). Purified PCR products were directly cycle-sequenced.
Table 1.
Primer pairs used in the study
| Name | Polarity | Sequence | Target virus | Target gene | Size (bp) | Reference |
|---|---|---|---|---|---|---|
| MAMD | Forward | 5′AACATGATGGGRAARAGRGARAA-3′ | Flavivirus | NS5 | 260 | (37) |
| cFD2 | Reverse | 5′GTGTCCCAGCCGGCGGTGTCATCAGC-3′ | ||||
| VIR2052F | Forward | 5’TGGCGCTATGATGAAATCTGGAATGTT-3′ | Alphavirus | nsP4 | 144 | (38) |
| VIR2052R | Reverse | 5′TACGATGTTGTCGTCGCCGATGAA-3′ | ||||
| MBPL3100F | Forward | 5′AGTCTCYTCTGCCATYTC-3′ | Phlebovirus | L | 890 | (39) |
| MBPL3287R | Reverse | 5′AGGATCTRGARGGGAACTTRT-3′ | ||||
| RHNB1520F | Forward | 5′ACIAAIAARTWIATGATGATGAA-3′ | Rhabdovirus | N | 188 | (40) |
| RHNB2134R | Reverse | 5′TGIARDATICCYTGCATCAT-3′ |
Sequence analysis
Amplified fragments were purified using a MinElute® PCR Purification Kit (QIAGEN) and directly sequenced using a BigDye Terminator Cycle Sequencing Kit v3.1 (Applied Biosystems, Austin, TX, U.S.A.) according to the manufacturer’s instructions.
Phylogenetic analysis of detected viruses
Phylogenetic relationships of isolated viruses were analyzed based on the available sequences of related virus species or strains obtained from the NCBI database. Phylogenetic analysis was performed by using the MEGA 7 program (13). Sequence data were aligned using the ClustalW method (14). Genetic distances were calculated using the p-distance model, and the phylogenetic tree was constructed using neighbor-joining methods, the reliability of the branching was evaluated by bootstrapping with 100 replications (15, 16, 17).
RESULTS
Mosquito collection
A total of 5,662 collected mosquitoes comprising at least 15 species of 5 genera from three districts of An Giang province (Table 2). The major collected mosquito species were Culex which consisted of 5,500 mosquitoes. Of which 4,255 mosquitoes were identified as Cx. tritaeniorhynchus (75.2%), 238 as Cx. vishnui (4.2%); Cx. quinquefasciatus, Cx. gelidus, Cx. theileri, Cx. tarsalis, Cx. Pseudovishnui with lower percentage (3.9, 2.8, 2.4, 2.3, and 2.2, respectively), and the lowest was Culex pipiens (0.1%). Small number of mosquitoes from other genera as Aedes, Anopheles, Armigeres, and Mansonia were also collected. The percentage of male mosquitoes was approximate 78 times lower than female mosquitoes, they were only collected from Ae.aegypti and Culex spp.
Table 2.
Number of collected mosquitoes and isolated viruses
Virus isolation
The collected mosquitoes were divided into 148 pools and used for virus isolation. Filter sterilized aliquots of homogenate’s mosquitoes were used to inoculate monolayers of C6/36 and Vero cells and were then observed for cytopathic effect (CPE). Eleven pools showed weak CPEs in C6/36 cells, CPE can be observed as cell growth retardation, shrinking cells, formation of a syncytium, or increasing gaps of cells. No CPE was observed in Vero cell cultures after five blind passages.
Detection of viral genomes
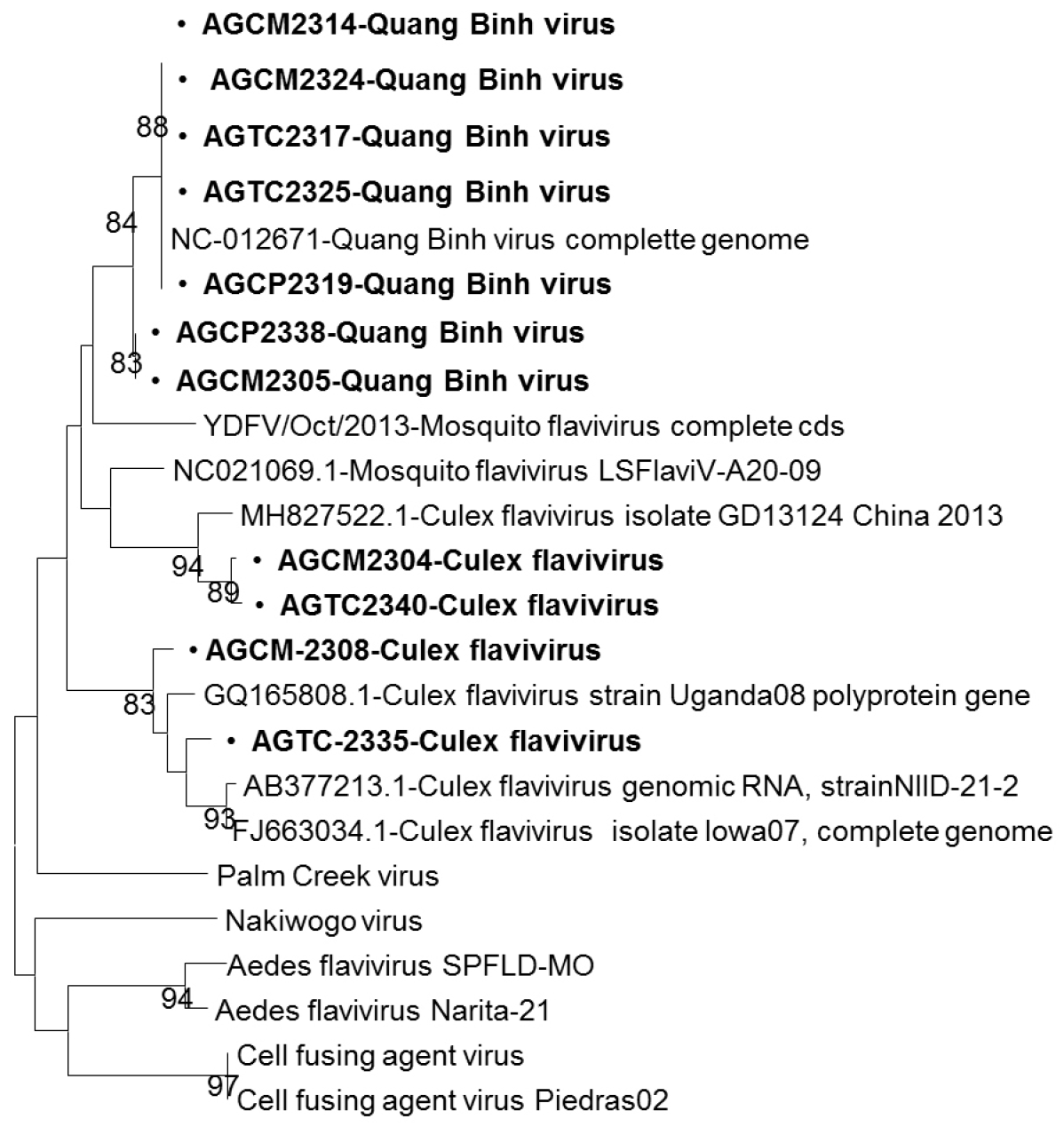
Eleven culture supernatants that showed CPEs in C6/36 cells were individually extracted RNA. These RNAs were analyzed by RT-PCR using four universal primer sets to detect viruses. These samples showed the positive results for the flavivirus primer set but negative for other primer pairs. Sequencing of these eleven RT-PCR products revealed that they showed high homology with sequences of insect-specific flaviviruses in the NCBI database. Among the eleven flavivirus-positive samples, six sample pools were obtained from Cx. tritaeniorhynchus, two from Cx. quinquefasciatus and Cx. fuscocephala, and one from Cx. gelidus (Table 2). Six pools of Cx. tritaeniorhynchus and one pool of Cx. gelidus showed 89.9-96.9% nucleotide identity to Quang Binh virus, complete genome with the GenBank accession no NC_012671.1. One Cx. fuscocephala and one Cx. quinquefasciatus pools demonstrated 95.2-98.8% nucleotide identity to that of Culex flavivirus isolated in China 2013 (GenBank accession no. MH827522.1). The remaining two mosquito pools of Cx. quinquefasciatus and Cx. fuscocephala had 89.9-90.6% nucleotide identity to that of Culex flavivirus genomic RNA, strainNIID-21-2 (Fig. 1).
Fig. 1
Phylogenetic analysis of isolated viruses. Phylogenetic analysis showing the genetic relationships of genotypes based on the partial NS5 gene by the maximum likelihood algorithm. Each branch was assessed by the bootstrap method with 100 replicates under the generalized time reversible model, sequence data for reference isolates were obtained from the NCBI database. Bold letters show isolates detected in this study.
DISCUSSION
There were five genera of collected mosquitoes in this study, of which Culex was predominant species. The most dominant mosquito species in this study was Cx. tritaeniorhynchus, accounting for 75.2% of the collected mosquiotes, Cx. vishnui (4.2%), Cx. quinquefasciatus (3.9%), Cx. sitiens, Cx. pipiens, Aedes, Armigeres, and Anopheles comprised a much smaller percentage. No Anopheles mosquitoes were captured in Cho Moi district. In this study, mosquitoes were captured for a mere half hour between 18:30 and 19:00 at each collection sites. Anopheles mosquiotes were much less common than Culex mosquitoes, which were captured between 18:00 and 19:00 (18, 19). The abundance of Cx. tritaeniorhynchus might be the tropics and subtropics are known as the highest suitable areas of Cx. tritaeniorhynchus, they breed mainly in the stagnant water like paddy fields or drainage ditches, therefore its habitat is mainly distributed along the river systems and water plays an important role in larval development. Cx. tritaeniorhynchus is also influenced the climate change, and also intimatively connected with pigs and cattle production (20). In addition, the activity of Cx. tritaeniorhynchus is mainly at night, with peak activity occurring one hour after dark (21). Cx. vishnui is primarily exophilic and zoophilic and their preferred breeding sites are rice fields and ponds, their dynamics are closely related to the phases of rice growth (22). In Vietnam, densities of Cx. vishnui is positively correlated with cattle density (23). Cx. vishnui has also been reported to bite pigs, goats, birds and also humans, especially when populations are high (22). Cx. quinquefasciatus is well adapted to the human environment, and prefers to suck human and cattle blood than pig blood. It breeds mainly in water polluted by organic substances such as sewer systems, toilets, stagnant ponds, and canals polluted by organic waste (23). The abundance of Cx. quinquefasciatus has been reported by the availability of the sites with the changing rainfall patterns. An Giang is a watershed province of Mekong Delta, and has a dense network of rivers that provide access to water year-round. Along with the abundant fresh water resources, the tropical climate and the rainfall make favorable for agricultural development with nearly 640,000 hectares and most areas grow three crops annually. Rice fields are often located at the intersection of human dwellings and natural areas, therefore rice-growing practices play an important role in the ecology of mosquitos in the province. The most important breeding habitat for the Culex mosquitoes is the rice crop ecosystem, rice paddy cover was positively associated with the spatial distribution of mosquito abundance (24). In addition, the mosquito habitat is mainly distributed along the river systems when water plays an important role in larval development, wind helps mosquitoes spread and migrate (25). Along with the development of crops, livestock which mainly cattle, pig, chicken and duck are also being promoted for development. Cattle, pigs, chicken often kept close to humans while duck mostly kept in the rice fields. In this study, all most mosquitoes were collected in the animal shelters while a small number mosquitoes was collected inside people’s houses.
The isolated viruses in the present study were Quang Binh virus and Culex flavivirus, both of them are insect-specific flaviviruses (ISFVs). Quang Binh virus (QBV), belongs to the genus Flavivirus, was initially isolated from Cx. tritaeniorhynchus mosquitoes in Quang Binh province, Vietnam, in 2002 (26). In this study, QBV was isolated from Cx. tritaeniorhynchus collected in animal shelters in Cho Moi, Tan Chau and Chau Phu districts, and from Cx. gelidus collected in animal shelters in Chau Phu district. This virus has also been isolated from other mosquito species as Anopheles sinensis, Aedes aegypti and Culex pipiens (27). Until now, QBV has been reported in Vietnam, and several provinces in China as Yunnan, Guizhou, Liaoning and Shaanxi province and Chongming district (20, 27, 28, 29). Culex flavivirus (CxFV) was first isolated from Cx. pipiens and Cx. tritaeniorhynchus collected in Japan, and Cx. quinquefasciatus collected in Indonesia during 2003-2004 (30). This virus was subsequently detected in Cx. restuans, Cx. tarsalis, Cx. interrogator, Cx. quinquefasciatus, and Anopheles (31, 32, 33, 34). In this study, CxFVs were isolated from Cx. quinquefasciatus and Cx. fuscocephala mosquitoes that collected in Cho Moi and Tan Chau districts. CxFV has been reported in many regions of Asia, South America, Central America, North America, and Africa (35). These results suggested that CxFV is insect-specific flavivirus specifically associated with Culex and Anopheles mosquitoes, and specialized to adapt to mosquito hosts at different regions. These isolated viruses in the study are belonged to insect-specific flaviviruses (ISFVs), these viruses naturally infect mosquitoes but do not infect humans or other vertebrates; they can replicate in mosquito cells in vitro, but can not replicate in vertebrate cells (36).
CONCLUSIONS
The findings of this study showed Culex tritaeniorhynchus was the most common mosquito species in the province and described the natural occurrence of Quang Binh virus and Culex flavivirus in the mosquito population of the rural areas in An Giang province. Though these detected viruses were insect-specific flaviviruses, further exploration and analysis of the transmission modes and life cycles of these isolated viruses should be investigated to help explain the host specificities of them, assess the possibility of the co-circulation, or horizontal transmission between different host species or genera in sympatric mosquito populations.
 XML Download
XML Download