

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Streptococcus pneumoniae (S. pneumoniae, Pneumococcus) remains a leading pathogen causing community-acquired pneumonia, bacteremia, sepsis, and acute otitis media (1). The capsule is pneumococcal outmost layer and covalently linked to the peptidoglycan and the target of current pneumococcal vaccines which provide the serotype-specific protection (2). Until now 100 capsule types, which are genetically, structurally, and serologically distinct, have been reported (3). Although the introduction of pneumococcal conjugated vaccines (PCVs) has significantly reduced infections by vaccine serotypes, the emergence of non-vaccine serotypes (NVTs; e.g. 24F, 33F) and new serotypes (e.g. 6C, 6D, 10D) poses a challenge to ongoing multivalent vaccine development (e.g., PCV15, PCV23) (4, 5). Opsonophagocytic killing assays are used to assess vaccine efficacy for various serotypes, but the lack of mouse infection models for some serotypes impedes the development of high multi-valent vaccines containing current NVTs (6).
Various animal models for pneumococcal research are currently in use to investigate pathogenesis mechanisms and to evaluate new drugs and vaccines (7). However, both animal models and pneumococcal strains or serotypes should be carefully considered before approaching the study of pneumococcal disease in vivo. The mouse is the most widely used model for examining pneumococcal colonization and pneumonia (8) and the specific pneumococcal diseases, such as meningitis, endocarditis, and otitis media, are typically studied using rat and chinchilla models rather than mice (9). The pneumococcal strains TIGR4 (ST4), WU2 (ST3), D39 (ST2), and A66.1 (ST6B) are most used in mouse models (10). Additionally, ST3, ST6B, and ST4 typically demonstrate invasiveness in mouse models (11). However, ST14, ST19F, and ST23F are reported to exhibit low or no virulence in mouse models. The reasons for this reduced virulence in certain serotypes remain controversial, potentially involving factors such as host specificity, complement specificity, and lectin binding affinity.
Organoids are in vitro mini-organ models derived from embryonic or adult stem cells or induced pluripotent stem cells. Stem cells are differentiated and proliferated into multiple organ cell type which highly mimic the characteristics of the human organs in vivo (12, 13, 14). Organoid systems have become attractive models for infectious diseases, particularly for non-mice infecting pathogens, during the Zika virus outbreak in 2016 and the SARS-CoV-2 pandemic in 2019 (12). Human lung organoids are widely used to study respiratory infections such as respiratory syncytial virus and influenza virus, as these pathogens primarily target lung epithelial cells and cause severe pneumonia or other diseases (15, 16, 17). These studies have provided a new field of research using the organoid platform for respiratory infectious diseases, which closely mimics the characteristics of human lung alveoli in vivo.
In this study, we demonstrated the potential of using ESC-derived lung alveolar organoids to investigate ST23F, which shows no infectivity in mouse pneumonia models.
METHODS
Ethics Statement
This study was performed in strict accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All animal experiments were approved by the Committee on the Use and Care of Animals at the Korea Atomic Energy Research Institute (KAERI; approval no. KAERI-IACUC-2023-004) and performed according to accepted veterinary standards set by the KAERI animal care center (RI-BIOMICS; Jeongeup, South Korea). Mice were euthanized by CO2 inhalation, as specified by the KAERI Institutional Animal Care and Use Committee guidelines.
Bacteria and Reagents
Most chemical reagents were sourced from Sigma-Aldrich (St. Louis, USA). Human embryonic stem cells (hESCs) and all reagents for the proliferation and culture of organoids were obtained from Organoid Science (Pangyo, South Korea) or STEMCELL Technologies (Vancouver, Canada). The bacterial strains used in this study were generously provided by Prof. Moon H. Nahm (University of Alabama at Birmingham). Pneumococcal strains were cultured in Todd Hewitt Yeast broth or agar at 37°C.
Mouse experiments
Specific pathogen-free female BALB/c mice, aged 6–7 weeks, were obtained from Orient Bio Inc. (Seoul, South Korea). Animal housing conditions, compliant with standards for specific pathogen-free animals, were provided by RI-BIOMICS (Jeongeup, South Korea) and approved by the Committee on the Use and Care of Animals at KAERI (Protocol 2023-003). The mice were housed in individually ventilated cages (Orient Bio Inc.) within a Biosafety Level 2 (BSL2) facility, maintained at 22–23°C with a 12-hour light/dark cycle. For experimental infection, mice (n=5) were challenged intranasally (i.n.) with pneumococci at 1×108 colony forming unit (CFU)/100 μL to compare lung colonization and blood invasion. At 1, 2, and 3 days post-infection, lungs were excised and homogenized in 1.5 mL PBS using a 40-μm mesh strainer. Bacterial load was quantified by culturing serially diluted lung homogenates or blood samples on blood agar plates (BAPs).
Lung organoid generation
To differentiate into definitive endoderm cells, the hESCs were seeded on a Matrigel-coated 6-well plate at 8 × 105 cells/2 mL/well and cultured with RPMI1640 medium containing B27 supplement (2%), human activin A (100 ng/mL), CHIR99021 (1 μM), and sodium butyrate (0.125 mM) for 6 days. For anteriorization, the cells were differentiated using a DMEM/F12 plus Glutamax medium containing B27 supplement (2%), L-ascorbic acid (0.05 mg/mL), monothioglycerol (0.4 mM), human noggin (100 ng/mL), and SB431542 (10 μM) for 4 days. Then, the cells were treated with DMEM/F12 plus Glutamax medium containing B27 supplement (2%), L-ascorbic acid (0.05 mg/mL), monothioglycerol (0.4 mM), human BMP4 (20 ng/mL), ATRA (0.05 μM), and CHIR99021 (3 μM) for additional 4 days to differentiated into ventralized anterior foregut endoderms (VAFEs). VAFEs were stimulated with human FGF10 (10 ng/mL), human KGF (10 ng/mL), CHIR99021 (3 μM), and DAPT (20 μM) for 7 days. On differentiation day 21, the cells were detached with Accutase and seeded onto ultra-low attachment 96-well U-bottom plate at 2 × 105 cells/250 μL/well to generate fibroblast-free spheroids. After 2 days, spheroids were gently collected and resuspened in 20 μL of human lung normal organoid medium (hLuN medium; Organoid Sciences) mixed with precooled Matrigel at a ratio of 1:1. The drop was embedded onto a 48-well plate to form a three-dimension culture environment and cultured with 400 μL of hLuN medium in the presence of Y27632 (10 μM). The hLuN medium was changed every 2 days. Embedded spheroids mature into human lung organoids (hLOs) by passaging every 5 - 7 days. Final organoids were sub-cultured more than three-times to remove cell debris and undifferentiated cells.
Real-time polymerase chain reaction (RT-PCR)
Organoids were collected and total RNA was isolated using the TRIzol Reagent (Thermo Fisher Scientific) according to the manufacturer’s protocol. Reverse transcription on 3 µg of total RNA was performed using random primers, dNTP mixture, and MMLV reverse transcriptase (Promega, Madison, WI, USA). Quantitative real-time PCR was performed using StepOne Real-Time PCR (Applied Biosystems, Foster City CA, USA) with the SYBR Green reagent (Takara, Tokyo, Japan). Primers were designed using Primer-BLAST, and the sequences are presented in Supplemental Table 1. The comparative Ct method was used, and relative mRNA expression was calculated based on the normalization to GAPDH expression. All experiments were repeated thrice.
Immunofluorescence Staining
hLOs were fixed with either 10% neutral formalin or Carnoy’s solution for 2 days and embedded in paraffin. Histology sections (5 μm) were collected. Formalin-fixed lung organoid sections were deparaffinized prior to performing antigen retrieval in sodium citrate buffer (10mM Sodium citrate, 0.05% Tween 20, pH 6.0). Sections were permeabilized with 0.2% Triton X-100 in PBS for 30 min, then incubated in a blocking buffer (5% BSA in PBS) for 1 h. Clara cell, AEC-I, AEC-II, and goblet cells were stained using anti-uteroglobin (CC10) rabbit monoclonal antibody (Santa Cruz Biotechnology, Dallas, TX, USA), anti-sodium potassium ATPase mouse monoclonal antibody (Abcam, Cambridge, UK), anti-surfactant protein B (SP-B) mouse monoclonal antibody (Santa Cruz Biotechnology), anti-mucin 5Ac (MUC5Ac) rabbit monoclonal antibody (Abcam) in a blocking buffer overnight at 4°C. Goat anti-mouse or rabbit secondary antibody conjugated to Alexa-488 or Alexa-568 were used according to manufacturer’s instructions (Thermo Fisher) for 1 h RT in blocking buffer. DAPI was used to stain DNA. Sections were mounted using coverslips and Prolong Glass Antifade Mountant (Thermo Fisher). Fluorescence images were taken using a Zeiss LSM 700 confocal fluorescence microscope (Carl Zeiss; Oberkochen, Germany). In a separate experiment, S. pneumoniae serotype ST23F or ST4 (4 × 108 CFU/mL) were stained by CellTrace CFSE (Thermo Fisher) and injected in hLOs using a FemtoJet 4i microinjector (Eppendorf, Hamburg, Germany) at a pressure of 200 hPA for 4 seconds. At 2 hours post-infection, the hLOs were fixed and treated with DAPI to stain nuclei. Fluorescence images were taken using a Zeiss LSM 700 confocal fluorescence microscope (Carl Zeiss).
Statistics
All experiments were repeated twice unless indicated otherwise. Data are presented as the mean ± SD of a representative experiment. Statistical significance was determined using one-way or two-way analysis of variance with Tukey’s multiple comparison post-hoc test. Statistical analyses were performed using GraphPad Prism 8.0 (GraphPad Prism Software, San Diego, CA, USA).
RESULTS
Pneumococcal ST23F is non-virulent serotype in mouse pneumonia model
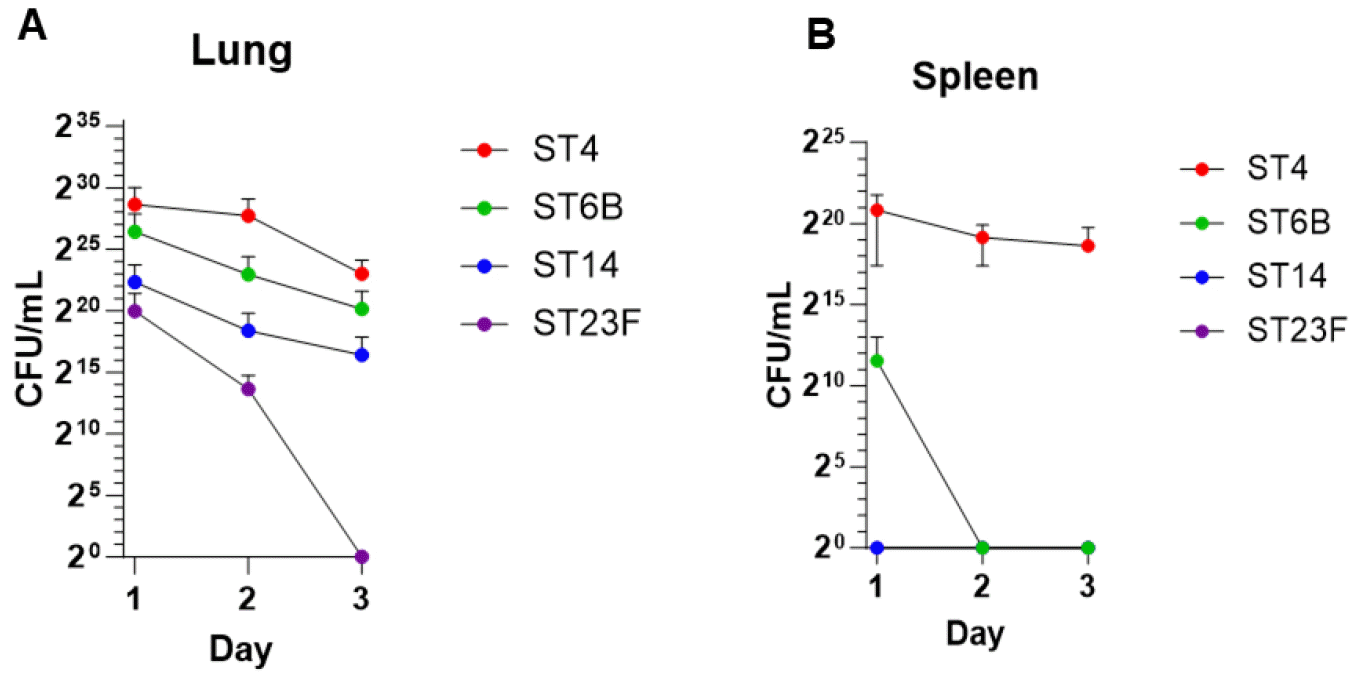
Four pneumococcal serotypes (ST4, ST6B, ST14, and ST23F) were selected to assess their virulence using a mouse pneumonia model. As reported previously, ST4 is highly virulent, ST6B shows moderate virulence, while ST14 and ST23F are low virulence in mice (11). Mice were intranasally inoculated with same numbers of pneumococci, and the bacterial load in the lungs and blood was counted from Day 1 to Day 3. Fig. 1A shows that all serotypes were present in the lungs on Day 1, with their CFU declining daily. In note, ST23F was rapidly cleared from the lungs, and disappearing by Day 3. Although there were slight differences, the colonization and clearance patterns of the other serotypes in the lungs were similar. To assess pneumococcal invasion from the lungs, bacterial numbers in the spleen were measured by plating on BAPs. As shown in Fig. 1B, no invasive pneumococci were detected in the spleen of mice infected with ST14 and ST23F. ST6B was detected on Day 1 but cleared by Day 2. In contrast, ST4 showed a high bacterial load from Day 1 through Day 3. In conclusion, ST4 exhibited high pathogenicity and ST6B showed moderate pathogenicity, indicating that both serotypes can effectively colonize the lungs and invade the bloodstream. ST14 demonstrated similar lung colonization patterns to ST6B but lacked the ability to invade the blood. Conversely, ST23F showed neither lung colonization nor blood invasion. Thus, ST14 is of low pathogenicity, and ST23F is non-pathogenic in mice.
Fig. 1
Lung colonization and blood invasion of Pneumococcus. Mice were intranasally infected with S. pneumoniae serotypes ST4, ST6B, ST14, or ST23F at a dose of 1×108 CFU/100 μL/mice (n=5). The bacterial count (CFU/g) in lung tissues (A) and blood (B) was determined by serial plating on BAP at days 1, 2, and 3. Values are shown as mean ± SD. Statistical significance was assessed using two-way ANOVA followed by Tukey’s test. *p < 0.05, **p < 0.01, ***p < 0.001. ns: not significant (p > 0.05).
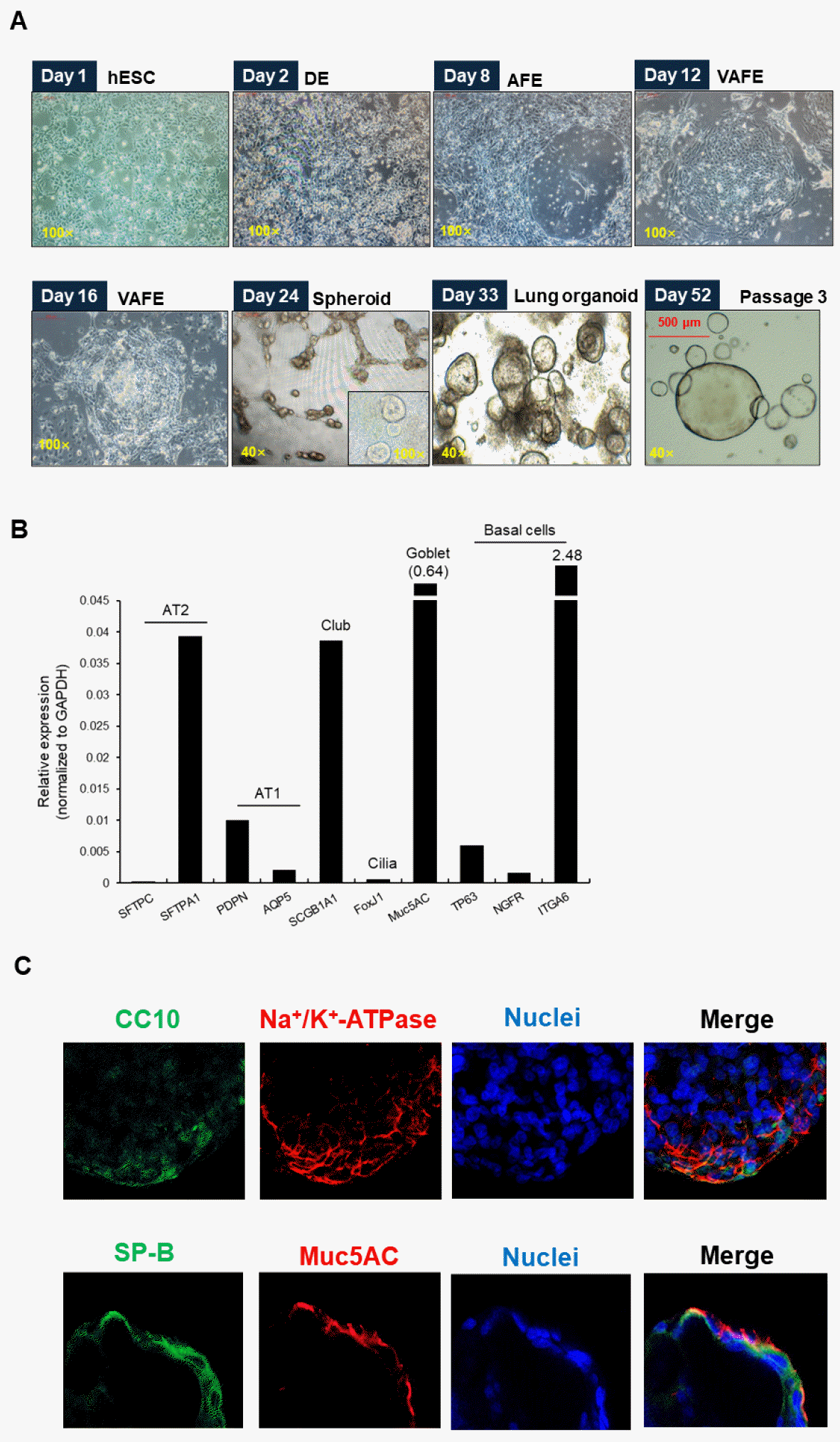
Generation and characterization of hESC-derived lung organoids
hESC-derived lung alveolar organoid was generated by following previous report (18). hESCs were sequentially differentiated by adding appropriate growth factors and differentiation factors to progress through the definitive endoderm (DE) stage (Day 2), anterior foregut endoderm (AFE) stage (Day 8), ventral anterior foregut endoderm (VAFE) stage (Day 12), lung progenitor stage (Day 18), and finally into lung organoids (Day 33) (Fig. 2A). hLOs were developed by cultured in the expansion media and monitored for growth characteristics by brightfield microscopy. They were cultured with consistent phenotypes until passage 10 and beyond, and passage 3–8 hLOs were used for experiments. To confirm the presence of major constituent cells in hLOs, we analyzed cell-specific markers using quantitative RT-PCR (qRT-PCR) and immunofluorescence. mRNA expression of various cell markers of constituent cells in hLOs was significantly higher in the organoids than other cell type (Fig. 2B), and immunostaining verified the presence of AEC-I markers (podoplanin; PDPN), AEC-II markers (surfactant protein A1; SFTPA1), goblet cell markers (MUC5AC), and club cell markers (uteroglobin; CC10) (Fig. 2C). These data suggested that ESCs were stably differentiated into cells that make up the lung through several stages by various growth factors.
Fig. 2
Generation of human embryonic stem cells (hESCs) derived lung organoids (hLOs). Cultured hESCs were transferred to Matrigel media for differentiation into lung organoids. (A) The progression from hESCs to hLOs was monitored daily, and images were captured at each differentiation stage using an inverted microscope, including the stages of definitive endoderm (DE), anterior foregut endoderm (AFE), ventralized anterior foregut endoderm (VAFE), lung progenitor, and mature lung organoids. (B) The mRNA expression of lung epithelial cell markers in hLOs was analyzed. Three wells of hLOs in a 48-well plate were collected on a differentiation day 52. The expression of specific cell type markers [SFTPC and SFTPA1 (AT2 markers); PDPN and AQP5 (AT1 markers); SCGB1A1 (club cell marker); FoxJ1 (cilia marker); Muc5AC (Goblet cell marker); TP63, NGFR and ITGA6 (basal cell markers)] was quantified using Real-time RT-PCR. Bar graph showed the relative abundance of various cell type markers normalized to GAPDH mRNA expression. (C) Human LOs were fixed and incubated with antibody specific to uteroglobin (CC10), sodium potassium ATPase, surfactant protein B (SP-B), or mucin 5Ac (MUC5Ac). The fixed hLOs were stained with Alexa-488 or Alexa-568-conjugated secondary antibody and the nuclei were counterstained with DAPI. The stained hLOs were examined by a confocal laser scanning microscope.
Interaction and invasion of ST23F in hLOs
Banked hLOs were sub-cultured in expansion media for 7-10 days. Once the hLOs had formed, more than 100 were microinjected with CFSE-stained ST23F or ST4 in PBS (4×106 CFU per hLO) (Fig. 3A and 3B). As shown in Fig. 3C, ST23F effectively invaded the epithelial cells of the hLOs which was relatively similar with hLOs infected with ST4. Interestingly, both ST23F and ST4 seems to be localized in specific epithelial cells (Fig. 3C), suggesting that they may primarily interact with AEC-II cells indicated as previous reports (19). To assess whether ST23F interaction with epithelial cells of hLOs leads to pro-inflammatory cytokine production, we performed qRT-PCR (Fig. 4). While no pro-inflammatory cytokines (TNF-α, IL-1β) were detected in the lung BALF of mice infected with ST23F (Fig. 5), hLOs infected with ST23F showed significant upregulation of all tested pro-inflammatory cytokines (TNF-α, IL-6, IFN-β, IFN-γ) comparable to the response observed in ST4-infected hLOs. CXCL10 and CCL2 chemokines were also upregulated in hLOs infected with either ST23F or ST3. But both serotypes were not induced the expression of IL-8 and TGF- β. These findings indicate that ST23F, a serotype non-virulent in mice, exhibits similar hLO infectivity to ST4, a highly virulent serotype in mice. This highlights the hLO model’s potential as a valuable tool for studying pneumococcal pathogenicity and evaluating pneumococcal vaccines.
Fig. 3
Infection of human lung organoids (hLOs) with pneumococcal ST4 or ST23F. CFSE-labeled Streptococcus pneumoniae serotypes ST23F or ST4 were microinjected into hLOs (passages 3-8). Bacterial suspensions, prepared at a concentration of 4 × 108 CFU/ml, were injected in hLOs for 4 seconds at a pressure of 200 hPA. (A) Schematic representation of the experimental design. (B) Image showing the injection site of ST23F using a microneedle. (C) At 2 hours post-infection (hpi), the hLOs were fixed and the nuclei were counterstained with DAPI. The fixed hLOs were examined by a confocal laser scanning microscope.
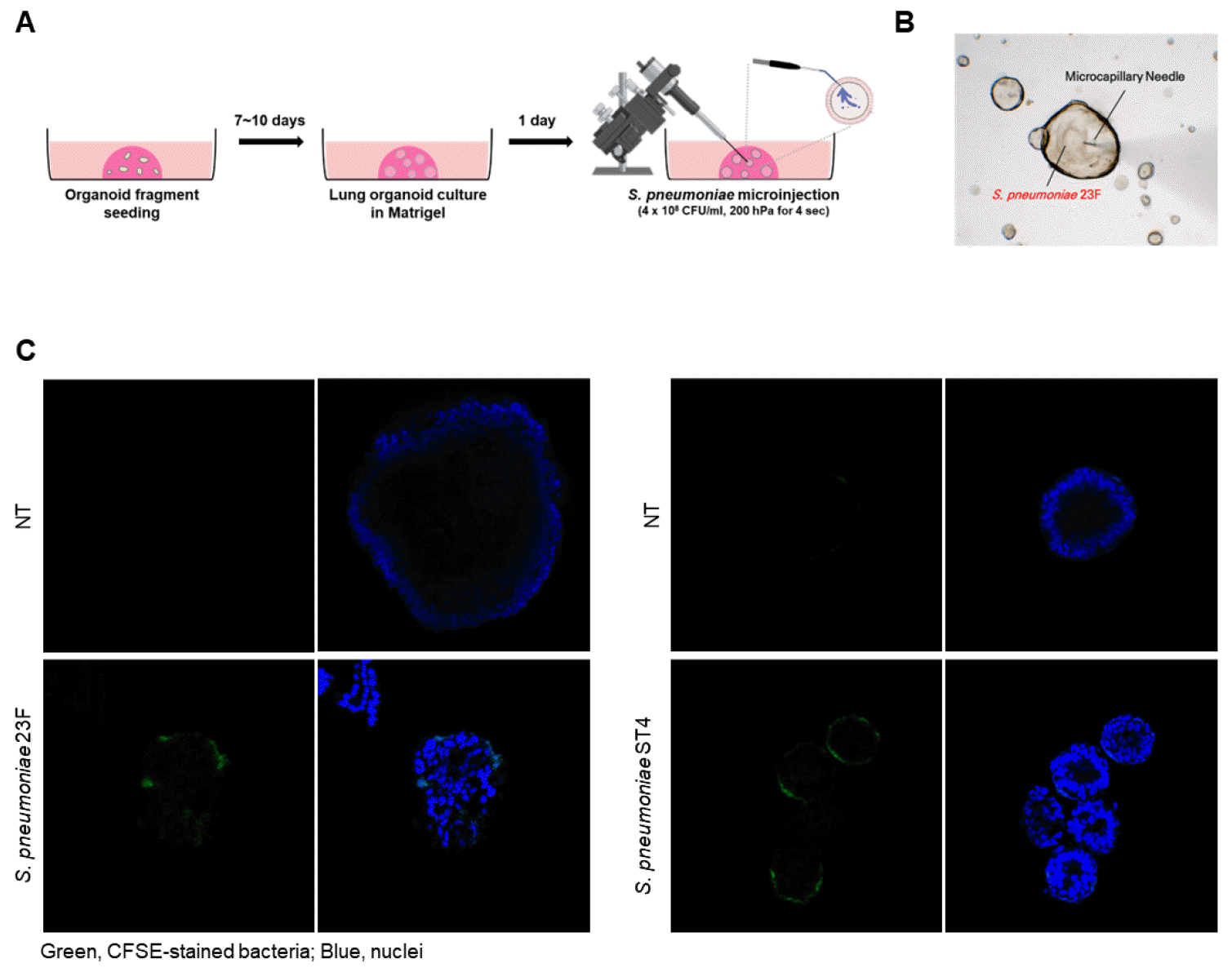
Fig. 4
Cytokine mRNA Expression in hLOs Infected with Pneumococcal Serotypes ST4 or ST23F. hLOs (n=100) were infected with Streptococcus pneumoniae serotypes ST4 or ST23F using a microinjector, and hLOs were collected at 12 hours post-infection. The mRNA levels of cytokines TNF-α (A), IL-6 (B), IFN-β (C), IFN-γ (D), CXCL10 (E), CCL2 (F), IL-8 (G), and TGF-β (H) were quantified by Real-time RT-PCR. Gene expression levels are shown as fold changes relative to uninfected hLOs after normalization to GAPDH mRNA. Data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s test. *p < 0.05.
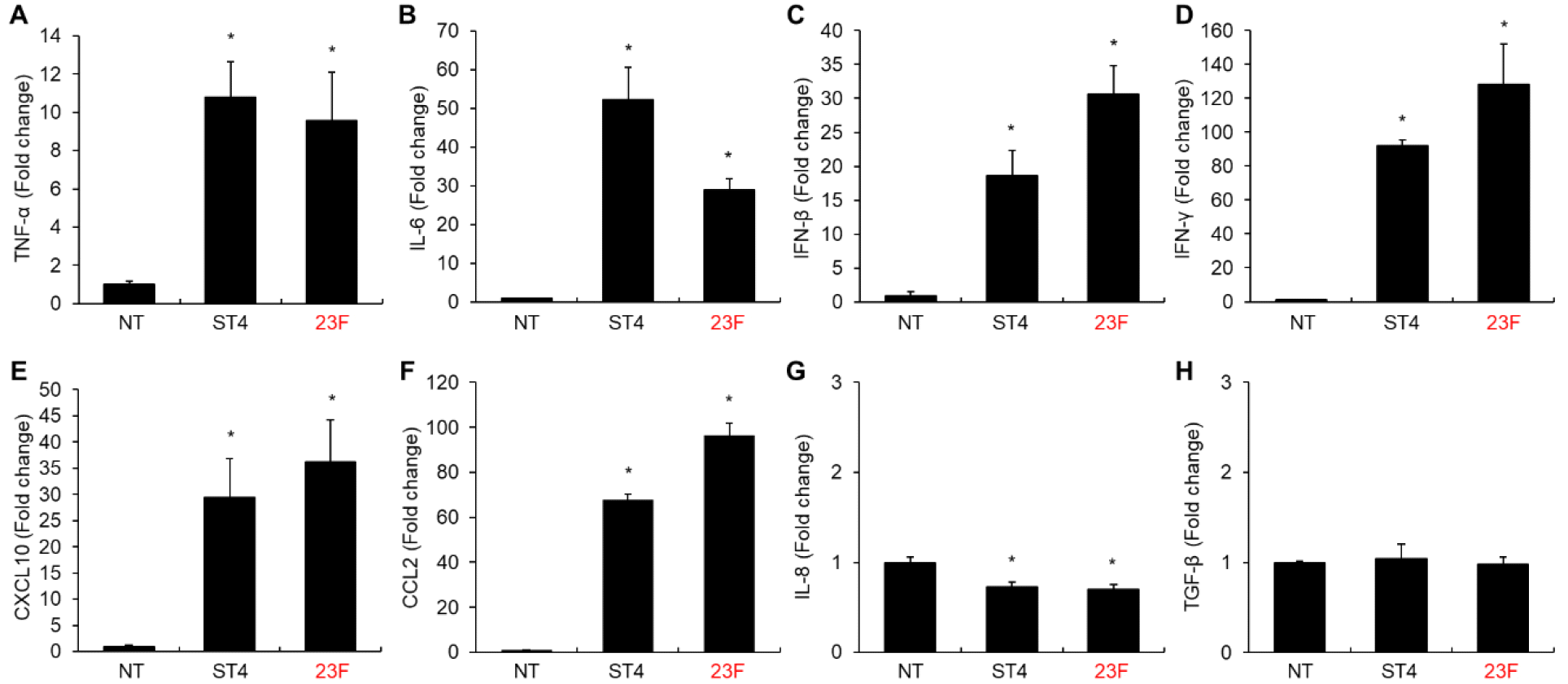
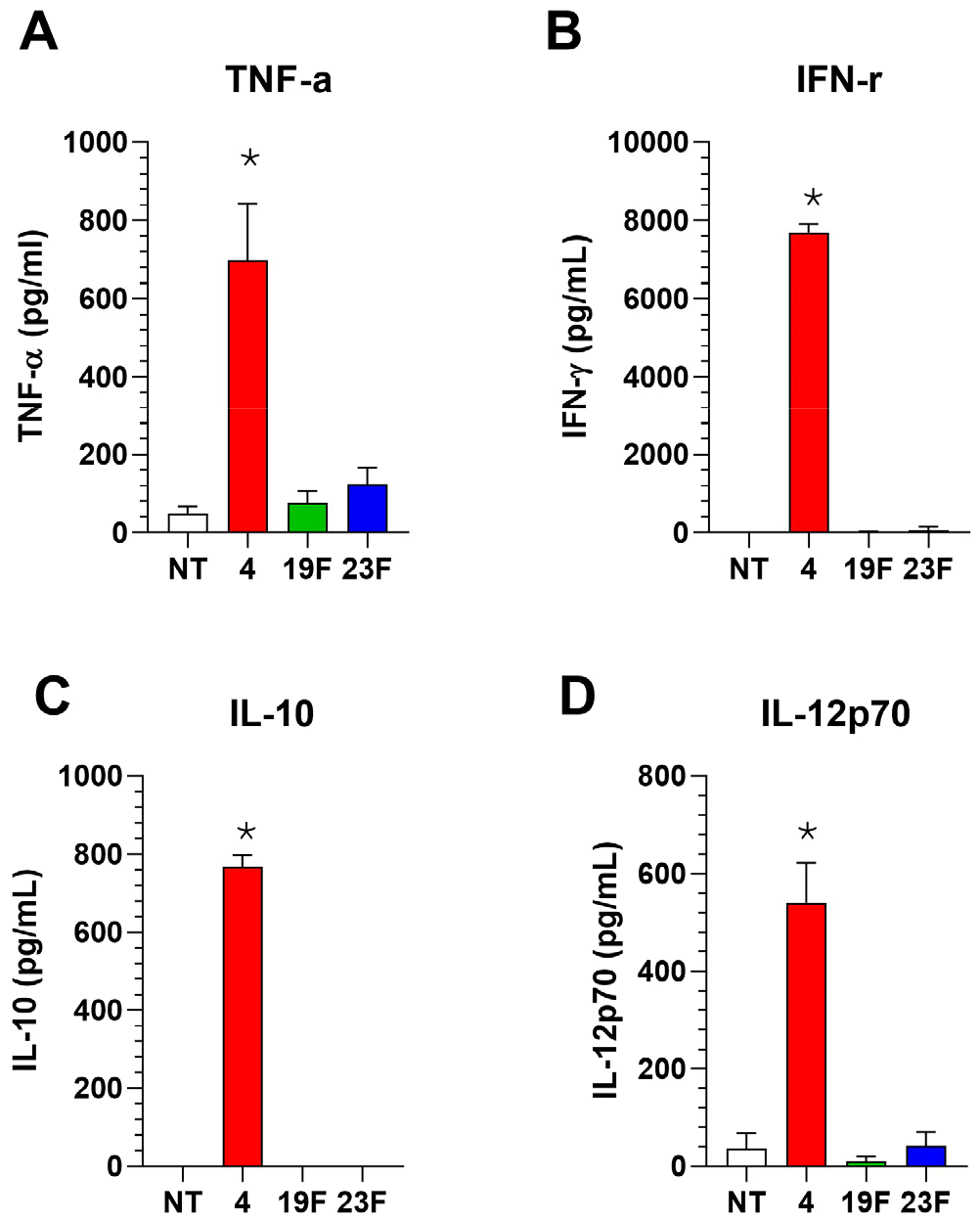
Fig. 5
Cytokine secretion in the BALFs of Mouse Infected with Pneumococcal Serotypes ST4 or ST23F. Mouse (n=5) were infected with Streptococcus pneumoniae serotypes ST4, ST19F, or ST23F intranasally, and mouse BALFs were collected at 24 hours post-infection. TNF-α (A), IFN-γ(B), IL-10(C), and IL-12p70(D) were quantified by ELISA. Data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s test. *p < 0.05.
DISCUSSION
This study is the first to demonstrate that hLOs provide as a viable alternative to traditional mouse infection models for assessing the virulence of pneumococcal serotypes that exhibit low or no infectivity in murine systems. Additionally, while significant differences in pathogenesis were observed among various serotypes in the mouse model, our findings reveal that both highly virulent and less virulent serotypes elicit comparable immunological responses in hLOs. This suggests that hLOs can effectively model human-specific pathogen-host interactions, providing valuable insights into pneumococcal virulence and immune response across serotypes.
The impact of capsular polysaccharide on the virulence of S. pneumoniae continues to be a debated issue in the field of microbial pathogenesis. After the introduction of the PCV vaccines, the prevalence of major vaccine serotypes, known for possession of high virulence, significantly decreased. However, NVTs have emerged as the predominant cause of invasive pneumococcal disease (20). Numerous studies have indicated that the capsule serves as a critical antigen with potent immune evasion properties, because the alteration of virulence and complement deposition observed when the capsular polysaccharide synthesis genes is exchanged in strains with identical genomic backgrounds (21, 22). In addition, ST23F was colonized more dominantly than ST4 in the mice pulp, but their colonizing ability was reduced and ST4 was completely dominated after 5 weeks old mice (23). Thus, pneumococcal colonization and invasion are influenced by a combination of host and bacterial factors, including the capsule serotype, the species of the host animal, the host immunity, and the host age. Therefore, findings from mouse models using specific serotypes might not fully replicate the human infection dynamics.
Airway organoids effectively replicate the respiratory tract, which serves as the primary interaction site for respiratory pathogens (24, 25). These organoids comprise various epithelial cell types, including basal, goblet, ciliated, club, and type I/II alveolar epithelial cells. Our study revealed that hESC-derived airway organoids express mRNA and protein markers for all these lung epithelial cell types. Recent advances in lung organoid research have been significantly influenced by studies on SARS-CoV-2, which showed no infectivity in mouse models (17). Moreover, lung organoids have been used to study viral replication, pathogenesis, and vaccine evaluation for several respiratory viruses, such as influenza virus and respiratory syncytial virus. However, using lung organoids to investigate bacterial infections and their pathogenesis is still relatively unexplored, because bacteria can trigger programmed cell death or toxin-mediated cell death. In fact, we observed that pneumococci can disrupt or destroy lung organoids within 12 hours of infections. Unlike lung alveoli, lung organoids lack immune cells in the alveolar space, which means they cannot provide a protective response against pneumococcal infection. Therefore, it is crucial to develop systems that better replicate the interactions between bacteria, epithelial cells, and immune cells in the lung organoid.
In this study, a notable finding was that lung organoids did not induce IL-8, a key chemokine in lung alveoli. Previous 2D cell culture studies have shown that pneumococci can stimulate IL-8 expression in lung epithelial cells (A549 and BEAS-2B) through toll-like receptors (26, 27). Interestingly, we observed that influenza virus A could induce IL-8 expression in the same lung organoids (unpublished data). This suggests two possibilities: (1) IL-8 induced by pneumococci might not be secreted by lung epithelial cells but instead by alveolar macrophages or other immune cells, or (2) lung organoids may not fully replicate the human lung system.
In summary, the lung organoid model offers a robust platform for nonclinical testing, enabling the assessment of pathogen virulence and the efficacy of novel therapeutics and vaccines, which are not easily evaluated in murine models. This is particularly critical for pneumococcal vaccine development, as various serotypes have recently emerged in regions implementing PCV vaccination programs, many of which are difficult to evaluate in mouse models. Therefore, lung organoids present significant potential for advancing the development of new pneumococcal vaccine.
 XML Download
XML Download