

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
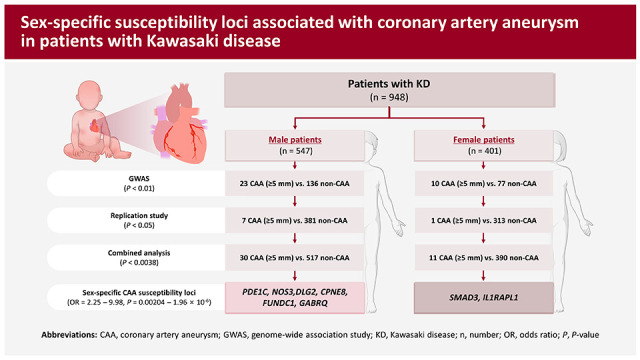
Kawasaki disease (KD) is a systemic vasculitis that predominantly affects children under 5 years of age, as well as being the leading cause of acquired heart disease in developed countries.1) KD is characterized by fever, rash, swollen lymph nodes, and inflammation of the blood vessels, particularly the coronary arteries.1) Coronary artery aneurysms (CAAs) are among the most frequent complications of KD, affecting approximately 15–25% of untreated children and 3–5% of children treated with high-dose intravenous immunoglobulins (IVIGs).2)3) CAAs are classified as small (internal diameter <5 mm), medium (internal diameter 5–8 mm), and giant (internal diameter >8 mm),4) with the infiltration of inflammatory cells playing a crucial role in the pathogenesis of CAAs in patients with KD.5)6) Several clinical factors, including male sex, resistance to IVIG treatment, prolonged duration of fever, and incomplete KD, have been found to influence the outcomes of CAAs in patients with KD.7)8) Specifically, male patients with KD have shown a greater susceptibility to CAA formation than females with KD.9)10) Although several genome-wide association studies (GWASs) using small sized samples have been performed to identify genetic loci associated with susceptibility to CAA formation in KD patients,11)12)13)14) the genetic factors contributing to CAA formation in patients with KD remain incompletely understood. The present study was designed to identify additional susceptibility genes associated with CAA formation in KD patients by performing a sex-stratified GWAS using previous GWAS data,15) comparing KD patients without CAA (controls) and with medium-sized aneurysms (diameter ≥5 mm) (cases). These findings revealed 6 male-specific and 2 female-specific CAA susceptibility genes in patients with KD, shedding light on the genetic factors involved in CAA formation in KD patients.
METHODS
Ethical statement
The study protocol was approved by the Institutional Review Board (IRB) of Asan Medical Center (IRB No. 2014-0823), and written informed consent was obtained from the parents of all the patients with KD.
Study subjects
A total of 1,975 children with KD were recruited from 12 tertiary academic hospitals in Korea (Supplementary Table 1), all of which were members of the Korean Kawasaki Disease Genetics Consortium. For the genetic study, 1,270 patients with KD were included (Table 1). KD was diagnosed by pediatricians based on the criteria of the American Heart Association (AHA).1)4) Each patient initially received a single infusion of high-dose (2 g/kg) IVIGs. In instances where patients did not respond to the initial treatment, additional doses of IVIG were administered. Two-dimensional echocardiography results obtained at worst were interpreted by pediatric cardiologists, with coronary arteries categorized as normal or abnormal based on the presence of coronary artery lesions, such as dilation or aneurysms. Additionally, the CAAs were classified by size into small (internal diameter 3 to 5 mm), medium (5 to 8 mm), and giant (≥8 mm).4)
Table 1
Demographic and clinical characteristics of the children with KD used in genetic analysis, stratified by size of CAA (n=1,270)
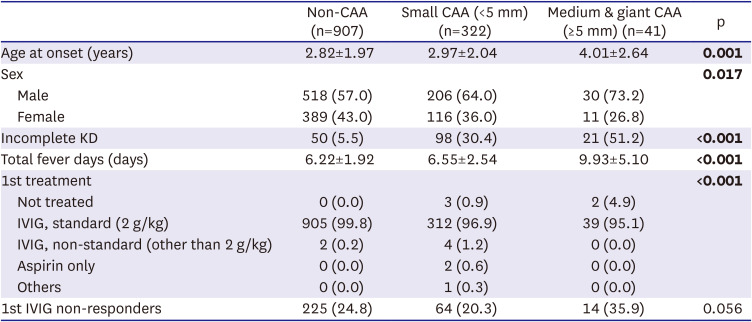
Values are presented as number (%). Quantitative variables are shown as mean±standard deviation. Differences were assessed by analysis of variance test or χ2 test, with significant differences shown in bold.
CAA = coronary artery aneurysm; IVIG = intravenous immunoglobulin; KD = Kawasaki disease; Non-CAA = no coronary artery aneurysm.
Genotyping and quality control
The genotype data utilized for the GWAS of CAA in KD cases were obtained from our previous study.15) Briefly, 296 children with KD were genotyped using the Illumina Human Omni1-Quad Bead Chip (Illumina, San Diego, CA, USA), according to the manufacturer’s instructions. Of these 296 children, 294 had available coronary artery data, including 213 children (136 males and 77 females) with KD but without CAA and 81 children (55 males and 26 females) with KD and CAA. Initially, a sex-stratified GWAS was performed on previous GWAS data to identify candidate susceptibility loci or genes associated with CAA in patients with KD. For GWAS, as single nucleotide polymorphism (SNP) quality control, we excluded 488 SNPs with missing call rates greater than 2%, 364 SNP markers with a Hardy-Weinberg Equilibrium p value <1×10−6 in controls, and 208,625 SNP markers with a minor allele frequency <0.01. After SNP filtering, 721,371 SNPs were included in the GWAS analysis. Thirteen candidate SNPs were selected based on their relevance to cardiovascular functions and/or significant associations with CAAs in either males or females with KD (p<0.01). To confirm these findings through replication, the same set of SNPs was genotyped in DNA samples from an additional 976 patients with KD, including 694 (381 males and 313 females) without CAA and 282 (181 males and 101 females) with CAA. Genotyping in the replication study was performed using either TaqMan genotyping assays (Applied Biosystems, Foster City, CA, USA) or the high-throughput Fluidigm EP1 system (Fluidigm Corp., South San Francisco, CA, USA), as described.16)
Statistical analysis
Statistical analyses were performed using SPSS Statistics, version 24, statistical software (IBM, Armonk, NY, USA). Between group differences in continuous variables were analyzed by Student’s t-test or the Mann-Whitney test. Differences among 3 or more groups were assessed by one-way analysis of variance (ANOVA), followed by Tukey’s test, or by the Kruskal-Wallis test. Categorical variables were analyzed using the χ2 test. Two-tailed p values <0.05 were considered statistically significant. Genetic associations between CAAs and SNPs in patients with KD were determined by comparing allele frequencies in cases and controls by χ2 tests using PLINK software, version 1.07 (https://zzz.bwh.harvard.edu/plink/).17) Loci on the X chromosome were separately analyzed in males and females, with the results merged by meta-analysis using PLINK software. The p values <0.01 (for analyzing the initial GWAS data), <0.05 (for analyzing the replicated data), and <0.0038 (for combined analysis of the 13 identified SNPs with Bonferroni correction, 0.05/13) were considered statistically significant. Haploview 4.2 was used for drawing a Manhattan plot (https://www.broadinstitute.org/haploview/haploview). The GWAS catalog database (https://www.ebi.ac.uk/gwas/) was examined to investigate the potential associations between candidate KD susceptibility genes and other diseases. The GTEx portal (https://gtexportal.org/home/) was also utilized to investigate differential gene expression of the CAA-associated variants in KD.
RESULTS
Males have a higher incidence of Kawasaki disease and a higher incidence of coronary artery aneurysm accompanying Kawasaki disease
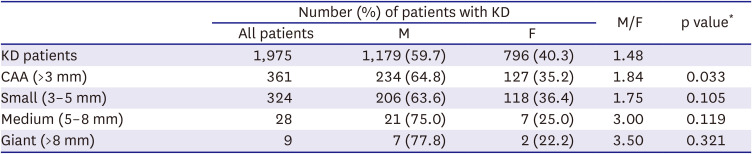
Analysis of clinical data from the Korean Kawasaki Disease Genetics Consortium revealed that, of the 1,975 patients with KD included in this study (Supplementary Table 1), 59.7% were male and 40.3% were female, resulting in a male-to-female ratio of 1.48:1 (Table 2). This result was consistent with several previous reports.18) Moreover, the incidence of CAA in patients with KD was significantly higher in males than in females (p=0.033), with a male-to-female ratio of 1.84 for CAAs with an internal diameter >3 mm. Although not statistically significant, the male-to-female ratio increased with increasing aneurysm size, being 1.75 for small (3–5 mm; p = 0.105), 3.00 for medium (5–8 mm; p = 0.119), and 3.50 for giant (>8 mm; p=0.321) CAAs (Table 2). These results indicate that the incidence of KD and the occurrence of CAA in patients with KD exhibit a sex bias, with the affected populations consisting predominantly of males.
Table 2
The effect of sex on the incidence of CAA in patients with KD (n=1,975)
Sex-stratified genome-wide association study identified 8 sex-specific susceptibility loci for coronary artery aneurysm in Kawasaki disease
To identify sex-specific susceptibility loci associated with CAA in patients with KD, a sex-stratified GWAS was performed using our previous GWAS data, involving 294 KD patients (191 males and 103 females) and an independent set of 976 patients with KD (562 males and 414 females). KD patients without CAA were categorized as controls, while those with aneurysms were considered cases. In particular, using patients with aneurysms having internal diameter ≥5 mm as cases, 13 candidate SNPs were selected in the initial sex-stratified GWAS (23 CAA vs. 136 non-CAA in male patients and 10 CAA vs. 77 non-CAA in female patients; Supplementary Figure 1) and tested in the replication study (7 CAA vs. 381 non-CAA in male patients and 1 CAA vs. 313 non-CAA in female patients; Supplementary Table 2). The combined analysis of both sets of samples resulted in the identification of 6 male-specific CAA susceptibility loci, involving the genes PDE1C (odds ratio [OR], 2.26; p=0.00184), NOS3 (OR, 2.25; p=0.00204), DLG2 (OR, 2.74; p=0.00012), CPNE8 (OR, 3.15; p=0.0.00186), FUNDC1 (OR, 3.96; p=0.00058), and GABRQ (OR, 9.98; p=1.96×10−6) (Table 3). Additionally, 2 female-specific CAA susceptibility loci were identified, involving the genes SMAD3 (OR, 4.59; p=0.00016) and IL1RAPL1 (OR, 4.35; p=0.00026) (Table 3).
Table 3
Sex-stratified genome-wide association results for CAA (internal diameter ≥5 mm) among patients with Kawasaki disease
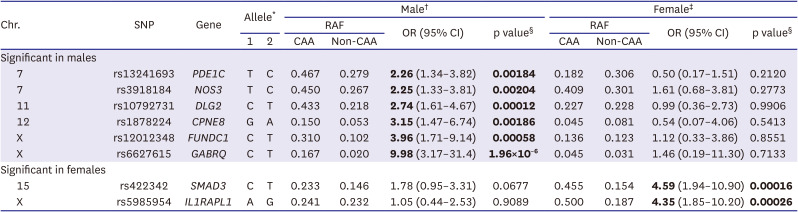
CAA (case group) = Kawasaki disease patients with coronary artery aneurysm (≥5 mm); CI = confidence interval; Non-CAA (control group) = Kawasaki disease patients without coronary artery aneurysm (<3 mm); OR = odds ratio; RAF = risk allele frequency; SNP = single nucleotide polymorphism.
*Allele 1 refers to risk allele.
†Male samples: 30 CAA (≥5 mm) samples and 517 non-CAA samples.
‡Female samples: 11 CAA (≥5 mm) samples and 390 non-CAA samples.
§Significant p values (p<0.0038 after Bonferroni correction with p value <0.05 of 13 SNPs) are shown in bold.
The additive effects of risk alleles for CAA formation were evaluated by calculating the numbers of risk alleles or weighted risk alleles, with the latter determined by multiplying the OR by the number of the 6 male-specific risk alleles present in individual male patients or by the number of the 2 female-specific risk alleles present in individual female patients. The number of risk alleles correlated significantly with aneurysm size in males, with the mean±standard deviation numbers of CAA risk alleles in patients without CAA and with small, medium, and giant CAAs being 1.88±1.36, 1.90±1.37, 3.86±2.01, and 4.25±1.28 (p=3.39×10−13), respectively. Similarly, the number of risk alleles showed an additive effect for CAA formation in females with KD (Table 4). These findings suggest that risk alleles have an additive effect in the development of CAAs, particularly for aneurysms having internal diameter ≥5 mm.
Table 4
Additive effects of risk alleles for CAA formation in patients with Kawasaki disease
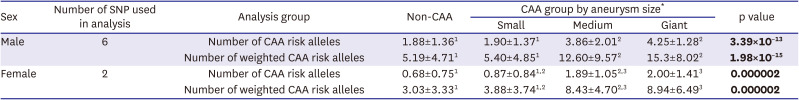
Data are presented as mean±standard deviation. Statistical analyses by analysis of variance and Tukey’s test. After Tukey’s test, values in the same column with different superscripts indicate significant differences between groups (p<0.05). Significant p values are shown in bold.
CAA = Kawasaki disease patients with coronary artery aneurysm (internal diameter ≥3 mm); Non-CAA = Kawasaki disease patients without coronary artery aneurysm; SNP = single nucleotide polymorphism.
*Aneurysm size was classified as small (3–5 mm), medium (5–8 mm), or giant (≥8 mm).
DISCUSSION
CAA is a serious complication of KD that can lead to significant morbidity and mortality, including increased risks of myocardial infarction, ischemic heart disease, sudden cardiac death, and the need for long-term monitoring and management.1)4)19) The incidence of CAA in KD patients varies based on sex, being much higher in males than in females.9)10) The sex-stratified GWAS performed in this study identified 6 male-specific (PDE1C, NOS3, DLG2, CPNE8, FUNDC1, and GABRQ) and 2 female-specific (SMAD3 and IL1RAPL1) CAA susceptibility loci.
Of these 8 CAA susceptibility genes, 3 (SMAD3, NOS3, and PDE1C) were previously found to be associated with arterial aneurysms. Mutations in SMAD3 have been linked to familial thoracic aortic aneurysms and dissections with intracranial and other arterial aneurysms.20)21) Knockout of the NOS3 gene in mice predisposed them to the formation of intracranial aneurysms.22) The eNOS G894T polymorphism (rs1799983) has been identified as a mild predisposing factor for abdominal aortic aneurysms.23) Additionally, the PDE1C gene was shown to play a critical role in regulating vascular structure remodeling and function,24) and to be expressed at high levels in human cardiac myocytes.25)
PDE1C has also been implicated in the development of abdominal aortic aneurysms.26) Interestingly, these 3 genes have also shown sex-specific differential expression and/or functions. For example, PDE1C gene expression levels in arterioles were higher in adult male than in adult female rats,27) with the T risk allele rs13241693 in the PDE1C gene being associated with the higher expression of PDE1C (Supplementary Table 3). These findings suggest that higher PDE1C gene expression and risk alleles for CAA in males may be involved in triggering CAA formation in patients with KD. Expression of the eNOS gene was shown to be higher in endothelial cells of human females than males, accompanied by differences in activity and function in vitro and ex vivo.28) In mouse models, differential sex-specific Smad3 gene expression in the liver and sex-dependent functions of Smad3 have also been observed.29)30) By contrast, the T risk allele rs3918184 in the NOS3 gene was found to be associated with higher expression of the neighboring KCNH2 gene, located 12.7 kb upstream of NOS3, in artery (aorta) tissue (Supplementary Table 3). Furthermore, the T risk allele rs4725982 in the KCNH2 gene was found to be significantly associated with CAA formation in patients with KD (OR, 2.77; p=0.0042; Supplementary Table 2), although the level of significance did not reach the Bonferroni corrected significance threshold (p<0.0038 in the combined analysis). Therefore, the role of NOS3 gene in CAA formation in patients with KD may be attributed to the direct effect of the NOS3 gene itself and/or its synergy with the neighboring KCNH2 gene that is involved in cardiovascular phenotypes, such as the QT interval, blood pressure, and electrocardiographic morphology (Supplementary Table 4).
This sex-stratified genetic association study resulted in the identification of 3 candidate susceptibility loci (FUNDC1, GABRQ, and ILRAPL1) on the X chromosome that were associated with CAA in patients with KD. Variants on sex chromosomes are usually excluded from GWAS due to differences in gene dosage when compared with autosomal chromosomes. Because KD itself and CAA events in patients with KD were found to exhibit a male-dominant pattern, this study hypothesized that identifying CAA-associated variants and loci on sex chromosomes could provide valuable insights into the sex-specific occurrence of CAA in patients with KD. Nevertheless, the specific role of these 3 genes in CAA formation remains unknown. Similarly, although the present study found that the DLG2 and CPNE8 genes were associated with CAA formation in patients with KD and that the G risk allele of rs1878224 in the CPNE8 gene increased CPNE8 gene expression, the roles of these genes have not been determined. Additional studies are needed to evaluate the involvement of these genes in CAA.
The present study focused on identifying CAA-associated variants with a strong effect size, a selected subset of KD patients having CAA of significant size (≥5 mm; only 1.87% of patients) was included in the genetic association study. This approach resulted in larger effect sizes and more significant p values than assessments of KD patients with CAAs >3 mm (data not shown), likely due to the enrichment of CAA-associated variants within the group containing larger CAAs. However, this approach also resulted to lower statistical power due to the small sample size.
This sex-stratified association analysis resulted in the identification of 8 sex-specific susceptibility loci associated with the formation of CAAs in patients with KD. Interestingly, the sex-stratified association study, both for males and females, exhibited greater significance and larger effect sizes than the combined analysis of both sexes. Furthermore, the analysis of variants revealed sex-specific CAA susceptibility loci on the X chromosome. These findings indicate that sex-stratified association analysis and examination of variants located on the sex chromosomes are valuable methods for identifying susceptibility loci associated with sex-skewed phenotypes.
The present study had several limitations, primarily due to the small sample size resulting from the selection of KD patients with CAA, the division of these patients by sex, and the choice of a CAA size threshold having internal diameter ≥5 mm. Additionally, we also have potential for misclassification of CAAs, attributable to variations in the diagnostic processes during multi-center data collection. Furthermore, we did not use the Z-score system to classify the CAA. The Z-score system to classify the CAA in KD were used in 2017 for the first time by AHA and then subsequently the system was partly adapted by the Japanese Ministry of Health in 2020. The data collection form of our consortium (the Korean Kawasaki Disease Genetics Consortium) was made in 2008 and used it for 15 years to collect clinical samples. At that time the Z-score system was not introduced yet. That is the reason why we cannot use the Z-score system.
Although this approach allowed the identification of several sex-specific loci associated with susceptibility for CAA formation in patients with KD, the limited number of samples led to relatively lower statistical significance. Additional studies are required to validate these findings in independent sample sets or other cohorts of patients with KD.
In summary, this sex-stratified GWAS identified 6 male-specific (PDE1C, NOS3, DLG2, CPNE8, FUNDC1, and GABRQ) and 2 female-specific (SMAD3 and IL1RAPL1) CAA susceptibility loci in patients with KD, with all having large effect sizes. These findings highlight the effectiveness of sex-stratified association studies in identifying susceptibility genes associated with sex-skewed phenotypes, particularly in the context of male-dominant events associated with CAA formation in patients with KD.
 XML Download
XML Download