

 PDF
PDF Citation
Citation Print
Print



Introduction
Spinal cord injury (SCI) is a serious neurological disease and constitutes a considerable portion of the global injury burden (1, 2). It has been reported that SCI has a high incidence worldwide; the highest incidence in the United States of America is approximately 906 per million, and the most common cause is traffic accidents (3). Injury to the spinal cord could cause damage to the motor, sensory, and autonomic functions of the spinal cord and physical, social, or psychological damage to patients (4). Moreover, it places a substantial burden on families and healthcare systems (5, 6).
At present, the main measures to improve the lives of patients with SCI mainly include surgery, drug therapy, and rehabilitation (7-10). Unfortunately, an effective treatment to completely repair SCI and improve functional recovery has not been found. Therefore, there is an urgent need to develop a safe and efficacious treatment that can repair SCI.
In recent years, several clinical studies have shown that stem cell therapy can improve motor function and neurological status in patients with SCI (11). In particular, mesenchymal stem cells (MSCs) are the most widely studied type of stem cells, which have a powerful ability in proliferation, differentiation, and tissue regeneration and can be obtained from different adult tissues, such as bone marrow, adipose tissue, muscle, umbilical cord, and placenta (12, 13). However, MSCs therapy also has some side effects in the clinic, including tumorigenicity, profibrogenic potential, and heterogenicity (3, 14). Fortunately, an increasing number of studies have shown that the therapeutic effects of MSCs are mainly related to exosomes secreted by MSCs, which have several advantages, including lower immunogenicity and an improved safety profile (15, 16). Exosomes contain nucleic acids, especially RNA, mainly including mRNA and microRNA (miRNA), the latter being considered bioactive compounds related to exosome function (17). Some studies have found that miRNAs associated with many secondary injuries are differentially expressed after SCI (18). Therefore, miRNAs may be potential targets for SCI therapy (19, 20).
In this review, we briefly describe the pathological process of SCI, provide information about exosomes, and summarize the repair mechanism of mesenchymal stem cells-derived exosomes (MSCs-Exos) in the treatment of SCI. Moreover, we listed the miRNAs found in exosomes and their related mechanisms, analyzed the unique advantages of exosomes as cargo carriers, and finally indicated that MSCs-Exos might become a promising new direction for SCI therapy in the future.
Go to : 
Mechanism of SCI
Although SCI is a complex process involving multiple factors, the related mechanism of SCI has gradually become clear with further research (21). At present, the SCI pathological process includes two aspects: primary injury and secondary injury (22). Primary SCI is mainly due to mechanical trauma resulting in fracture and displaced bone fragments, disc material, and ligaments pressing directly on the spinal cord, which will lead to damaged blood vessels, disrupted axons, and broken neural-cell membranes (22, 23). The characteristics of primary injury are local hemorrhage, edema, and ischemia (24). Secondary SCI produces multiple cascades of chemical events that cause further tissue loss and dysfunction. These events can be divided into three distinct but often continuous sequences: acute (<48 hours), subacute (48 hours∼14 days), and chronic (14 days∼6 months) (25). The acute phase is characterized by vascular injury, blood-spinal cord barrier (BSCB) disruption, inflammation, ionic dysregulation, free radical production, and glutamate excitotoxicity. As the disease progresses, the SCI enters a subacute phase characterized by macrophage infiltration, astrocyte proliferation, glial scar formation, and neuronal apoptosis. Finally, it enters a chronic phase characterized by Wallerian degeneration, glial scar maturation, myelomalacia, and cyst formation. All possible mechanisms are summarized in Fig. 1 (4, 22, 26-28).
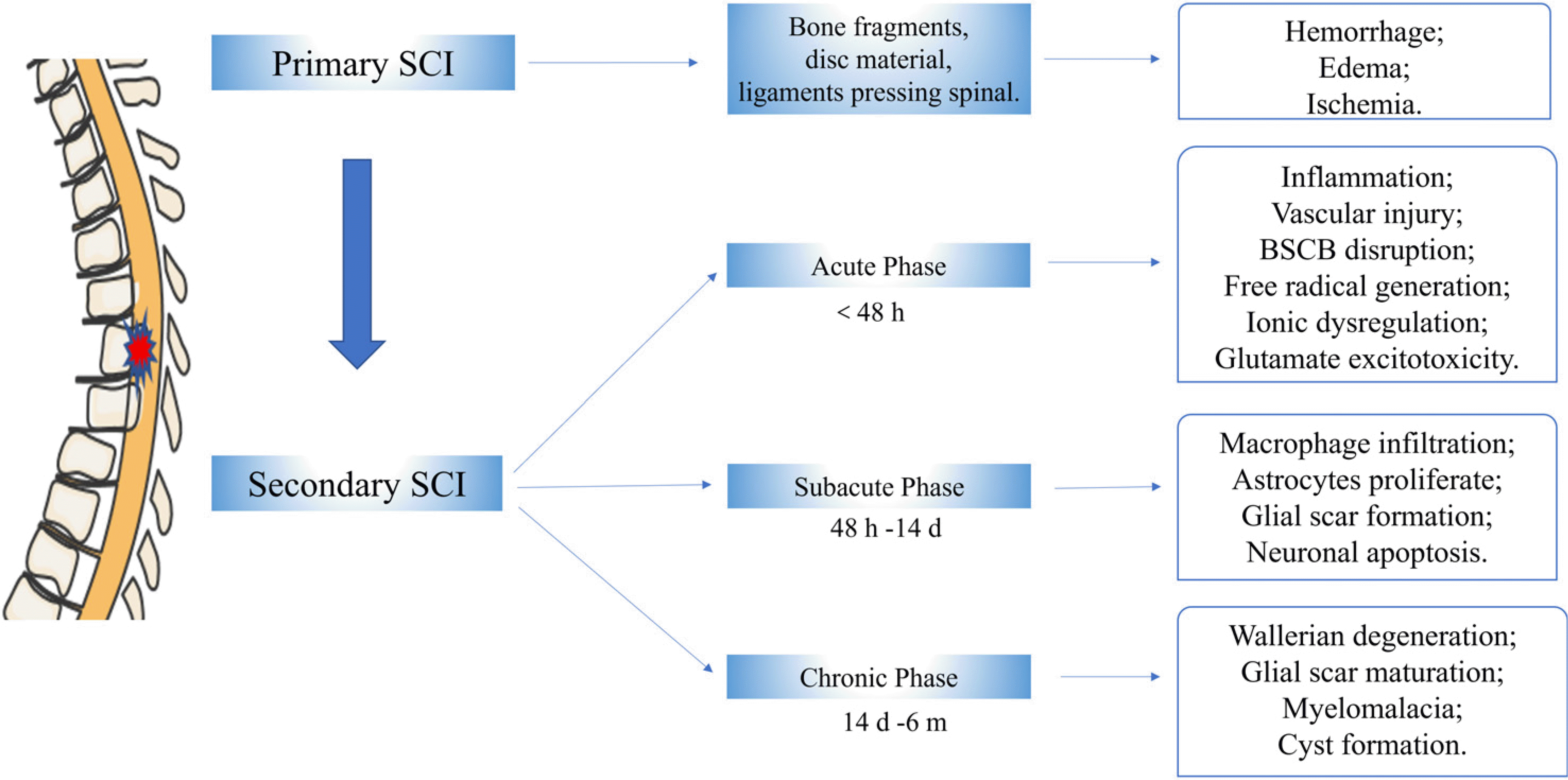
Go to :
Exosomes and MSCs-Exos
Extracellular vesicles (EVs) are lipid-bound vesicles released by all cells, which are divided into three categories depending on their origin and biogenesis: apoptotic bodies, which are produced by the fragmentation of apoptotic cells; ectosomes, which are produced by direct outgrowths from the plasma membrane; and exosomes, which are produced by inward outgrowths from endosomal compartments that subsequently fuse with the plasma membrane (29, 30). Among them, exosomes are vesicles with a diameter of about 40∼160 nm, which contain various components such as specific lipids, proteins, and nucleic acids (31). Because exosomes have the smallest average diameter, complex composition, and diverse functions, they are the most studied EV types at present (30). Recent research has found that exosomes are actively secreted by most cells and widely exist in all body fluids and tissues, playing an important role in intercellular communication (32-35).
Exosomes are generated in a process that involves double invagination of the plasma membrane and the formation of intracellular multivesicular bodies (MVBs) containing intraluminal vesicles (ILVs), which can be summarized as four steps: budding of the plasma membrane, ILV formation, MVB formation, and exosome secretion (Fig. 2) (31, 36). The formation process and mechanism of exosomes are complex, and most of them act through the endosomal sorting complex required for transport (ESCRT). The ESCRT system contains four protein complexes, ESCRT-0, ESCRT-1, ESCRT-2, and ESCRT-3, together with auxiliary proteins, such as VTA⁃1, ALIX, and VPS4 (36, 37). Notably, some studies have found that some exosomes can also be produced in the absence of ESCRT, and these mechanisms involve lipids, tetraspanins, or heat shock proteins. These approaches are collectively called ESCRT-independent mechanisms (38, 39). The process of exosome secretion into the extracellular matrix is mainly through MVB transport to the plasma membrane and fusion with the plasma membrane, depending on the synergy of several Rab GTPases and SNAREs (soluble N-ethylmaleimide-sensitive fusion attachment protein receptors) complexes (31, 40, 41). Finally, the released exosomes can directly enter cells through three different mechanisms: receptor-mediated entry, direct membrane fusion, and endocytosis, as shown in Fig. 2 (40).
 | Fig. 2Exosome formation, secretion, and action on target cells. Formation of mesenchymal stem cells-derived exosomes (MSCs-Exos): ① Endocytosis: The extracellular component and cell surface protein are engulfed into the cells through the invagination of the plasma membrane to form an early endosome (EE); ② First exchange: The EE could initially exchange goods with the Golgi network and endoplasmic reticulum to form the late endosome (LE); ③ Secondary exchange: LE conducts cargo exchange again through the second plasma membrane invagination and finally forms intraluminal vesicles (ILVs), and the collection of multiple ILVs of different sizes is called a multivesicular body (MVB); ④ Plasma membrane budding: Some MVBs release exosomes extracellularly through plasma membrane budding; ⑤ Recycling: Some MVBs can be degraded by lysosomal fusion, and the degradation products can be recovered by cells. The structure of MSCs-Exos: Exosomes express tetraspanins (CD81, CD63, and CD9), heat shock proteins (HSP60, HSP70, and HSP90), ALG-2 interacting protein X (Alix), TSG101, integrins, flotillin, surface markers (CD44, CD73, and CD90) and adhesion molecules (CD29, CD44, and CD73). In addition, MSCs-Exos carry nucleic acids, proteins, and enzymes. Exosomes enter recipient cells in three ways: ⑥ receptor-mediated entry, ⑦ direct membrane fusion, and ⑧ endocytosis.
|
MSCs-Exos is one of the most studied exosomes. MSCs-Exos contain multiple proteins with multiple functions, including ESCRT complexes, ALG-2 interacting protein X (Alix), tumor susceptibility gene 101 (Tsg101), Rab GTPases, integrins, heat shock protein (HSP60, HSP70, and HSP90), and MHC class I and class II proteins (42, 43). In addition, MSCs also have characteristic tetraspanins (CD81, CD9, and CD63) and some surface markers (CD44, CD73, and CD90) (Fig. 2) (31, 44). MSCs-Exos participate in intercellular communication, which can carry proteins, mRNA, and miRNA into other cells (45). They can participate in the metabolism, energy pathways, cell growth, communication, and transport of target cells and regulate a wide range of normal physiological and pathological processes (46, 47). In particular, nucleic acids are abundant in exosomes, including mRNAs, miRNAs, mitochondrial DNA and piRNAs, ncRNA, ribosomal RNA, and snRNA (48). Among these RNAs, miRNAs have been the most extensively studied. They are endogenous small noncoding RNA molecules 18∼25 nucleotides in length that can control the expression of mRNAs, protein production, and cell function at the posttranscriptional level (49, 50). Primary miRNAs are transcribed, processed, and transported to the cytoplasm and then cleaved to generate double-stranded mature miRNAs, which often interact with target messenger RNAs, resulting in translational repression and degradation of these target mRNAs (49, 51). Biologically, miRNAs have been implicated in development, differentiation, proliferation, apoptosis, and immune responses (52). Currently, increasing evidence shows that MSCs-Exos and their miRNAs have great potential in promoting functional recovery after SCI, which is another new hope after stem cell treatment of SCI.
Go to :
Mechanism of MSCs-Exos treatment of SCI
Compared with MSCs, MSCs-Exos have the advantages of low cytotoxicity, penetrating deep tissues, being able to escape the rapid clearance of the phagocytic system, and penetrating the blood-brain barrier, so there are an increasing number of studies on exosomes for the treatment of SCI (53). At present, a large number of animal experiments and cell experiments have proven that MSCs-Exos contribute to the repair of SCI (Table 1), but there are few related clinical studies. We summarize the mechanisms related to the existing research on MSCs-Exos and their miRNAs in the treatment of SCI.
Table 1
Current preclinical studies on MSCs-Exos in the treatment of spinal cord injury
| References | Types of exosomes | Research objects | Type of miRNA | Mechanism | Effect |
|---|---|---|---|---|---|
| (57) | BMSC-Exos | Rat and PC12 cell | - | Inhibit the TLR4/MyD88 /NF-κB pathway | Inhibit the inflammatory response and apoptosis |
| (58) | BMSC-Exos | Rat | miR-181c | Inhibit the PTEN and NF-κB signaling pathway | Reduce spinal cord inflammation and apoptosis |
| (59) | BMSC-Exos | Rat and BV2 cells | miR-23b | Inhibit TLR4/NF-κB pathway | Attenuate the inflammatory cytokine response |
| (60) | AD-MSCs-Exos | Rat | - | - | Inhibit the release of proinflammatory cytokines |
| (70) | EF-MSCs-Exos | Rat | - | Inhibit NLRP3 inflammasome activation, decrease the level of pro-apoptotic protein Bax | Reduce the expression of inflammatory cytokines and reduce apoptosis |
| (71) | HWJ-MSCs-Exos | Rat | - | Decrease the expression of caspase 1 | Attenuate neuroinflammation |
| (80) | BMSC-Exos | Rat | miR-125a | Negatively regulate IRF5 expression | Promote M2 macrophage polarization and reduce the inflammatory response |
| (81) | BMSC-Exos | Rat | miR-124-3p | Negatively regulate Ern1 expression | Promote M2 macrophage polarization |
| (82) | MSCs-Exos | Mice | miR-216a-5p | Inhibit TLR4/NF-κB and activate the PI3K/Akt signaling pathway | Promote M2 macrophage polarization |
| (88) | BMSC-Exos | Rat | - | Inhibit nuclear translocation of NF-κB p65 | Reduce A1 astrocytes |
| (89) | BMSC-Exos | Rat | - | - | Suppress activation of A1 neurotoxic reactive |
| (90) | HUC-MSCs-Exos | Rat | miR-146a-5p | Inhibit the Traf6/Irak1/ NFκB pathway | Reduces the toxic effects of neurotoxic astrocytes |
| (96) | HP-MSCs-Exos | Mice | - | - | Promote the new vessel formation |
| (97) | MSCs-Exos | Endothelial cells | - | Rromote VEGF expression through activation of PKA signaling pathway | Enhance angiogenesis |
| (98) | MSCs-Exos | Rat | - | - | Promote the new vessel formation |
| (114) | BMSC-Exos | Rat | - | Activate the Wnt/β-catenin signaling pathway | Inhibit neuronal apoptosis |
| (115) | HUC-MSCs-Exos | Rat | - | Regulate BCL2/Bax and Wnt/β-catenin signaling pathways | Anti-apoptosis and anti-inflammatory |
| (116) | BMSC-Exos | Rat | - | Regulate the expression of pro-apoptoticprotein cleaved caspase-3,anti-apoptotic protein Bcl-2, and autophagy-related proteins LC3IIB and Beclin-1 | Promote autophagy to reduce neuronal apoptosis |
| (117) | MSCs-Exos | Rat and SH-SY5Y cells | miR-21 and miR-19b | Regulate PTEN expression | Regulate the apoptosis and differentiation of neuron cells |
| (118) | MSCs-Exos | Rat | miR-21 | Regulate PDCD4 and PTEN expression | Inhibit the apoptosis of neurons |
| (119) | MSCs-Exos | Rat | miR-21-5p | Downregulate the expression of the pro-apoptotic target gene FasL | Inhibit the apoptosis of neurons |
| (120) | MSCs-Exos | Rat | miR-29b-3p | Activate the Akt/mTOR pathway through the PTEN axis | Inhibit the apoptosis of neurons |
| (121) | MSC-Exos | Rat | miR-381 | Downregulate the BRD4/ WnT5A axis | Reverse neuronal apoptosis |
| (122) | AD-MSCs -Exos | Rat | miR-499-5p | Inhibit JNK3/MAPK10 and negatively regulate the JNK3/c-jun signaling pathway | Regulate neuronal apoptosis |
| (123) | BMSC-Exos | Rat and PC12 cell | miR-9-5p | Regulate the HDAC5/FGF2 axis | Alleviate apoptosis and inflammation |
| (124) | AD-MSCs-Exos | Rat and PC12 cell | miR-511-3p | Regulate TRAF6/S1P axis | Inhibit neuronal apoptosis and inflammatory |
| (129) | BMSC-Exos | Rat | - | Activate the NF-κB signaling pathway | Maintain BSCB integrity |
| (130) | BMSC-Exos | Rat | - | Suppress pericyte pyroptosis | Maintain BSCB integrity |
| (131) | MSCs-Exos | Rat | - | Increase the expression of cell junction proteins via the TIMP2/MMP pathway | Attenuate the destruction of the BSCB |
| (132) | MSCs-Exos | Rat | - | Upregulate TGFβ, TGFβ receptors, and tight junction proteins | Reduce BSCB permeability |
|
(134, 135) |
MSCs-Exos | Cortical neurons | - | - | Promote the growth of axons in neurons |
| (136) | HP-MSCs-Exos | Rat | - | Activate of the MEK/ERK/CREB signaling pathway | Neural progenitor cells activation and neurogenesis |
| (137) | MSCs-Exos | Rat | - | Inhibit the NF-κb/NLRP3 signaling pathway | Promote axon regeneration and myelin regeneration |
| (138) | HUC-MSCs-Exos | Rat and PC12 cell | miR-199a-3p/145-5p | Regulate NGF/TrkA signaling pathway | Promote axonal growth |
| (139) | MSCs-Exos | Rat | - | Upregulate R-Smad 6 expression by carrying TGFβ | Promote neuronal regeneration |
| (140) | MSCs-Exos | Rat | miR-let- 7a-5p | Regulate the HMGA2/ SMAD2 axis | Promote neuronal regeneration |
MSCs-Exos: mesenchymal stem cells-derived exosomes, miRNA: microRNA, BMSC: bone marrow stem cell, PTEN: Phosphatase and Tensin homolog, AD: adipose tissue, EF: epidural fat, HWJ: human Wharton’s jelly, HUC: human umbilical cord, HP: human placenta, VEGF: vascular endothelial growth factor, BSCB: blood-spinal cord barrier.
![]()
Regulation of the neuroinflammatory response
Inhibiting inflammation by exosomes: The inflammatory response induced by SCI is an important factor in secondary injury. Microglia, neutrophils, macrophages, and lymphocytes accumulate and release various cytokines in the microenvironment due to the disruption of capillaries, the BSCB, and chemokines, which unbalance the local microenvironment (54, 55). The microenvironment contains multiple proinflammatory factors (IL-1, IL-6, and TNF-α) and anti-inflammatory factors (IL-4, IL-8, IL-10, and IL-13) that are related to the functional recovery of SCI patients (56). Therefore, modulating local microenviron-mental inflammation may be a therapeutic approach for SCI (10, 56).
Several studies have demonstrated that exosomes can modulate the levels of inflammatory factors (Fig. 3). Fan et al. (57) showed that bone marrow stem cell-derived exosomes (BMSC-Exos) can attenuate the inflammatory response and apoptosis of SCI by inhibiting the TLR4/MyD88/NF-κB signaling pathway. Zhang et al. (58) found that miR-181c-enriched BMSC-Exos could downregulate the levels of TNF-α and IL-1β to reduce spinal cord inflammation and apoptosis, and the study illustrated that the mechanism may be the inhibition of Phosphatase and Tensin homolog (PTEN) and NF-κB signaling. Nie and Jiang (59) also made a similar discovery that miR-23b delivered by BMSC-Exos could decrease the levels of proinflammatory cytokines (IL-6, IL-1β, and TNF-α) and increase the levels of anti-inflammatory cytokines (IL-10). miR-23b also promotes the transformation of microglia to the M2 type. Further studies revealed that it attenuates the inflammatory cytokine response by inhibiting the TLR4/NF-κB pathway. Recently, Sung et al. (60) demonstrated that exosomes from human epidural adipose tissue-derived MSCs (AD-MSCs) attenuated local inflammatory responses by inhibiting the release of proinflammatory cytokines after SCI.
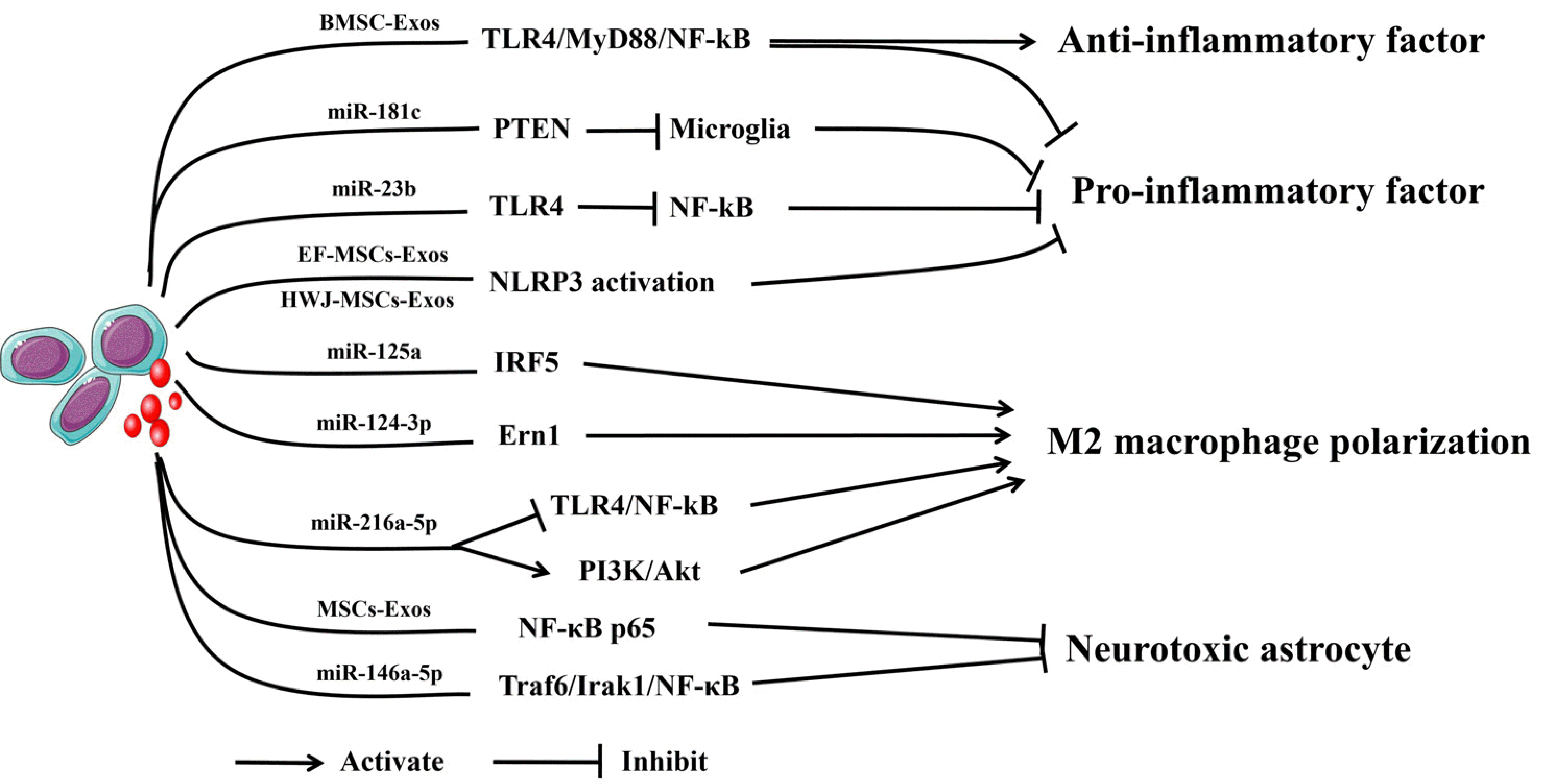 | Fig. 3The mechanism by which mesenchymal stem cells-derived exosomes (MSCs-Exos) regulate the immune response. Bone marrow stem cell (BMSC)-Exos can regulate the TLR4/MyD88/NF-κB signaling pathway to produce the production of anti-inflammatory factors. MSCs-Exos with miR-181c and miR-23b can inhibit the formation of microglia via the Phosphatase and Tensin homolog (PTEN) signaling pathway and inhibit the TLR4/NF-κB pathway to inhibit the production of proinflammatory factors. Epidural fat (EF)-MSCs-Exos and human Wharton’s jelly (HWJ)-MSCs-Exos can inhibit the NLRP3 activation to inhibit the production of proinflammatory factors. MSCs-Exos with miR-125a and miR-124-3p can inhibit M2 macrophage polarization by regulating the IRF5 and Ern1 signaling pathways. MSCs-Exos with miR-216a-5p can inhibit M2 macrophage polarization by inhibiting TLR4/NF-κB and activating the PI3K/Akt signaling pathway. MSCs-Exos can inhibit the NFκB p65 signaling pathway to reduce astrocyte toxicity. MSCs-Exos with miR-146a-5p can inhibit the Traf6/Irak1/NF-κB signaling pathway to reduce astrocyte toxicity.
|
In addition, neuroinflammation, which is characterized by the activation of central immune cells triggered by various causes, also plays a key role in the secondary injury process of SCI, and the most important step is the activation of various inflammasome complexes (61, 62). Inflammasome complexes consist of three components: a sensor, adaptor protein apoptotic speck protein, and effector caspase-1 (63). Currently, the sensors that have been identified include NLR family pyrin domain containing (NLRP)1, NLRP3, and NOD like receptor family CARD domain containing 4, and absent in melanoma 2 (64, 65). Studies have found that the expression of the above four inflammasomes is upregulated after SCI injury (64, 65), and inhibition of inflammasome activation reduces neuronal death and promotes motor recovery (66-68). Recently, several studies have shown that exosomes can inhibit the activation of inflammasomes to attenuate inflammatory responses (69). Huang et al. (70) suggested epidural fat mesenchymal stem cells-derived exosomes (EF-MSCs-Exos) via tail vein injection to improve neurological functional recovery, and the possible mechanism is to inhibit NLRP3 inflammasome activation and reduce the expression of inflammatory cytokines. In addition, Noori et al. (71) indicated that human Wharton’s jelly mesenchymal stem cells-derived exosomes (HWJ-MSCs-Exos) attenuated neuroinflammation after SCI by decreasing the expression of caspase 1, a component of the inflammasome complex, and the inflammatory cytokines IL-1β, IL-18, and TNF-α. Thus, MSCs-Exos and their miRNA can modulate the levels of inflammatory factors to improve the outcome of SCI.
Promotion of macrophage polarization by exosomes: Microglia are resident immune cells of the central nervous system (CNS) and are mainly derived from microglial progenitor cells derived from the embryonic yolk sac (72, 73). Macrophages are derived from blood mononuclear cells, which are derived from myeloid progeny in the bone marrow (74). After SCI, the destruction of the cerebrospinal barrier allows monocytes to infiltrate the spinal cord tissue and transform into macrophages, which together with resident microglia activate and participate in inflammatory responses (75). Notably, macrophages and microglia were morphologically and immunohistologically indistinguishable after activation (76). Thus, activated microglia and blood-derived monocytes are collectively referred to as CNS macrophages (77). Macrophages have two main polarization phenotypes: M1 and M2 (74, 78). M1 macrophages (proinflammatory cells) produce various pro-inflammatory factors (IL-6, IFN-γ, IL-12, IL-23, IL-1β, and TNF-α), reactive oxygen species (ROS), and nitric oxide to promote inflammation and neuronal damage. In contrast, M2 macrophages (anti-inflammatory cells) have anti-inflammatory and tissue repair properties because they express high levels of the anti-inflammatory factors IL-10 or transforming growth factor (TGF)β, and some studies have confirmed that the predominance of M1 macrophages compared with M2 macrophages can aggravate injury after SCI (54, 76, 79). Therefore, the regulation of macrophage polarization may be a treatment method for SCI.
Some studies have reported that MSCs-Exos can promote macrophage polarization (Fig. 3). Chang et al. (80) found that miR-125a from BMSC-Exos could promote M2 macrophage polarization by negatively regulating interferon regulatory factor 5 (IRF5) expression and ultimately reduce the inflammatory response caused by SCI and promote the recovery of motor function. Li et al. (81) showed that exosomal miR-124-3p from BMSC attenuated nerve injury induced by SCI, which may inhibit endoplasmic reticulum to nucleus signaling 1 (Ern1) and promote M2 pola-rization. Liu et al. (82) suggested that exosomal miR-216a-5p from hypoxia-treated MSCs inhibited TLR4/NF-κB signaling pathway and activated the PI3K/Akt signaling pathway to convert microglia from the M1 proinflammatory phenotype to the M2 anti-inflammatory phenotype and increase the treatment potential of SCI. In addition, Liu et al. (83) revealed that dental pulp stem cell-derived exosomes could suppress M1 macrophage polarization by inhibiting the ROS-MAPK-NF-κB P65 signaling pathway after SCI. Thus, MSCs-Exos and their miRNA can promote macrophage polarization to improve the outcome of SCI.
Regulation of astrocytes by exosomes: Astrocytes are involved in CNS repair and play a very important role in the process of SCI (84-87). There are two types of reactive astrocytes, A1 and A2 astrocytes, which are induced by neuroinflammation (induced by IL-1α, TNF-α, and C1q) and ischemia. A1 astrocytes have neurotoxic effects on myelin sheaths, synapses, and neurons; in contrast, A2 astrocytes exert neuroprotective effects by upregulating the expression of certain neurotrophic factors (86, 87). There-fore, reducing the proportion of A1 neurotoxic astrocytes may be a potential treatment strategy for SCI.
There are few reports that MSCs-Exos reduce A1 neurotoxic astrocytes (Fig. 3). Wang et al. (88) found that intravenous administration of MSCs-Exos exerts anti-inflammatory and neuroprotective effects in a rat model of SCI, possibly by inhibiting nuclear translocation of NF-κB p65 to reduce SCI-induced A1 astrocytes. Liu et al. (89) identified that BMSC-Exos can inhibit the activation of A1 neurotoxicity-responsive astrocytes and promote functional behavioral recovery after acute SCI. Lai et al. (90) showed that exosomal miR-146a-5p from human umbilical cord mesenchymal stem cells (HUC-MSCs) significantly reduces the toxic effects of neurotoxic astrocytes and promotes the recovery of neurological function in injured rats, and its possible mechanism is to inhibit the activation of the NF-κB pathway by suppressing the expression of TNF receptor associated factor 6 (TRAF6) and interleukin 1 receptor associated kinase 1 (IRAK1). Thus, MSCs-Exos and their miRNA can reduce A1 neurotoxic astrocytes to improve the outcome of SCI.
Promotion of angiogenesis by exosomes
Blood vessels can transport multiple ingredients (oxygen and nutrients), remove metabolic waste, and facilitate cell circulation (91, 92). After SCI, vascular injury leads to ischemia, hemorrhage, and immune cell infiltration, accelerating the onset of secondary injury and neurological deficits (93, 94). Studies have proven that angiogenesis is one of the most important conditions for ensuring tissue survival and functional regeneration (95). Therefore, promoting angiogenesis is another research direction for SCI.
Several studies have shown that exosomes can promote angiogenesis after SCI (Fig. 4). Zhang et al. (96) found that human placenta mesenchymal stem cells-derived exosomes (HP-MSCs-Exos) could promote tube formation by endothelial cells in vitro and new vessel formation in vivo and improve neurologic function in mice post-SCI. Xue et al. (97) revealed that MSCs-Exos induced by hypoxia have been found to accelerate angiogenesis by promoting vascular endothelial growth factor (VEGF) expression through activation of the PKA signaling pathway. Recent research also revealed that hypoxia-stimulated exosomes derived from MSCs could be transplanted into the injured spinal cord through an adhesive hydrogel, which could promote angiogenesis and functional recovery after injury in vitro and in vivo (98). Furthermore, besides exosomes from MSCs, exosomes derived from neural stem cells (99), M2 macrophages (100), microglia (101), and Schwann cells (102) have also been found to promote angiogenesis to facilitate functional recovery after SCI. However, the mechanism by which exosomes promote angiogenesis still needs further research. Thus, MSCs-Exos can promote angiogenesis to improve the outcome of SCI.
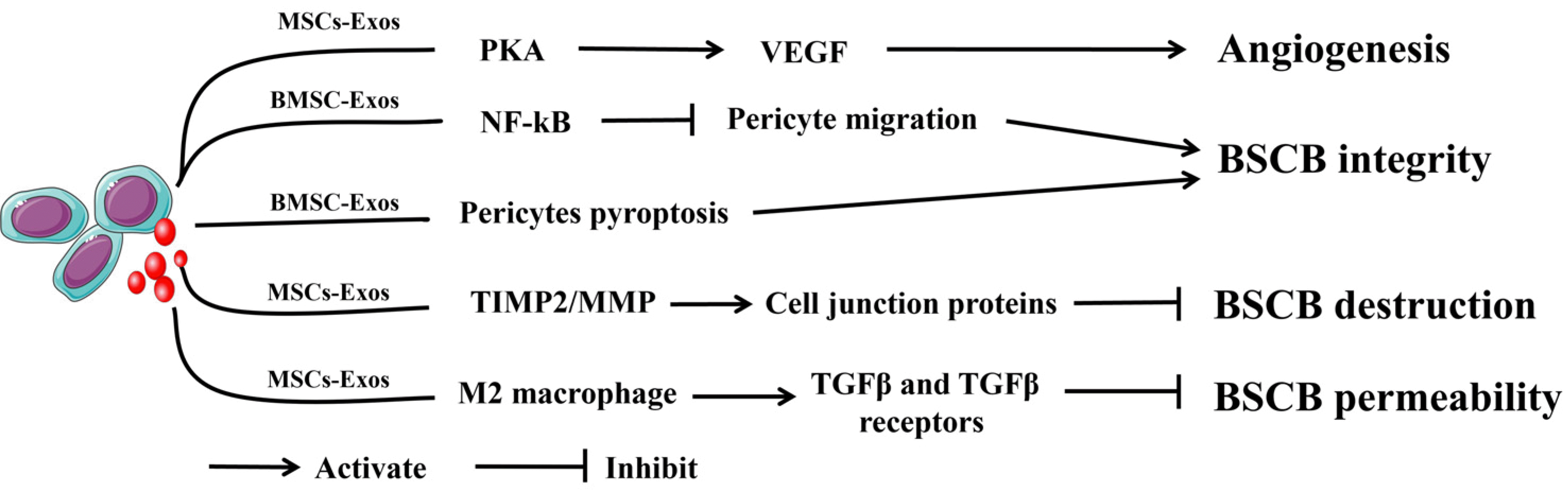 | Fig. 4The mechanism by which mesenchymal stem cells-derived exosomes (MSCs-Exos) promotes angiogenesis and regulates blood-spinal cord barrier (BSCB) integrity. MSCs-Exos can promote angiogenesis by promoting the expression of vascular endothelial growth factor (VEGF) via the PKA signaling pathway. BMSC-Exos can suppress pericyte pyroptosis and inhibit pericyte migration by activating the NF-κB signaling pathway to maintain BSCB integrity. MSCs-Exos could increase the expression of cell junction proteins via the TIMP2/MMP pathway to attenuate the destruction of the BSCB. MSCs-Exos could reduce BSCB permeability by targeting M2 macrophages to upregulate TGFβ and TGFβ receptor expression.
|
Activation of autophagy and inhibition of apoptosis by exosomes
Autophagy is an important defense and protection mechanism for an organism. Autophagy mainly participates in cell recycling by degrading damaged and denatured proteins, organelles, and other substances, thus enabling neurons to survive in an environment deficient in nutritional factors (103-105). Proper activation of autophagy is beneficial to eliminate toxic proteins and damaged mitochondria for promoting neuroprotection in traumatic SCI (106). Previous studies have found that the levels of autophagy biomarkers (Beclin-1, LC3B II, and P63) are elevated during the acute phase of secondary injury in SCI, indicating that autophagy was significantly activated (107, 108). In addition, apoptosis, also known as programmed cell death, plays a crucial role in SCI-induced cell death (109, 110). Inhibition of apoptosis signaling pathways could impede neuronal cell death (111). Some studies have shown that activation of autophagy and inhibition of neuronal apoptosis can promote the recovery of neural function after SCI (112, 113). Therefore, modulation of autophagy and apoptosis is a promising therapeutic strategy for SCI.
Recent studies have demonstrated that autophagy and apoptosis are closely related to exosomes (Fig. 5). BMSC-Exos can inhibit neuronal apoptosis by activating the Wnt/β-catenin signaling pathway (114). EF-MSCs-Exos could decrease the expression level of the proapoptotic protein Bax and upregulate antiapoptotic proteins to reduce apoptosis after SCI (70). HUC-MSCs-Exos improved rat motor function after SCI through anti-apoptosis and anti-inflammatory effects, possibly by regulating the B-cell lymphoma 2 (Bcl-2)/Bcl-2-associated X protein (Bax) axis and Wnt/β-catenin signaling pathways (115). Another study found that BMSC-Exos not only enhanced the expression of the autophagy-related proteins LC3IIB and Beclin-1 and induced autophagosomes to form but also reduced the expression level of the proapoptotic protein cleaved caspase-3 and upregulated the expression level of the antiapoptotic protein Bcl-2. These effects ultimately attenuate neuronal apoptosis by promoting autophagy and functional behavior recovery in SCI rats (116).
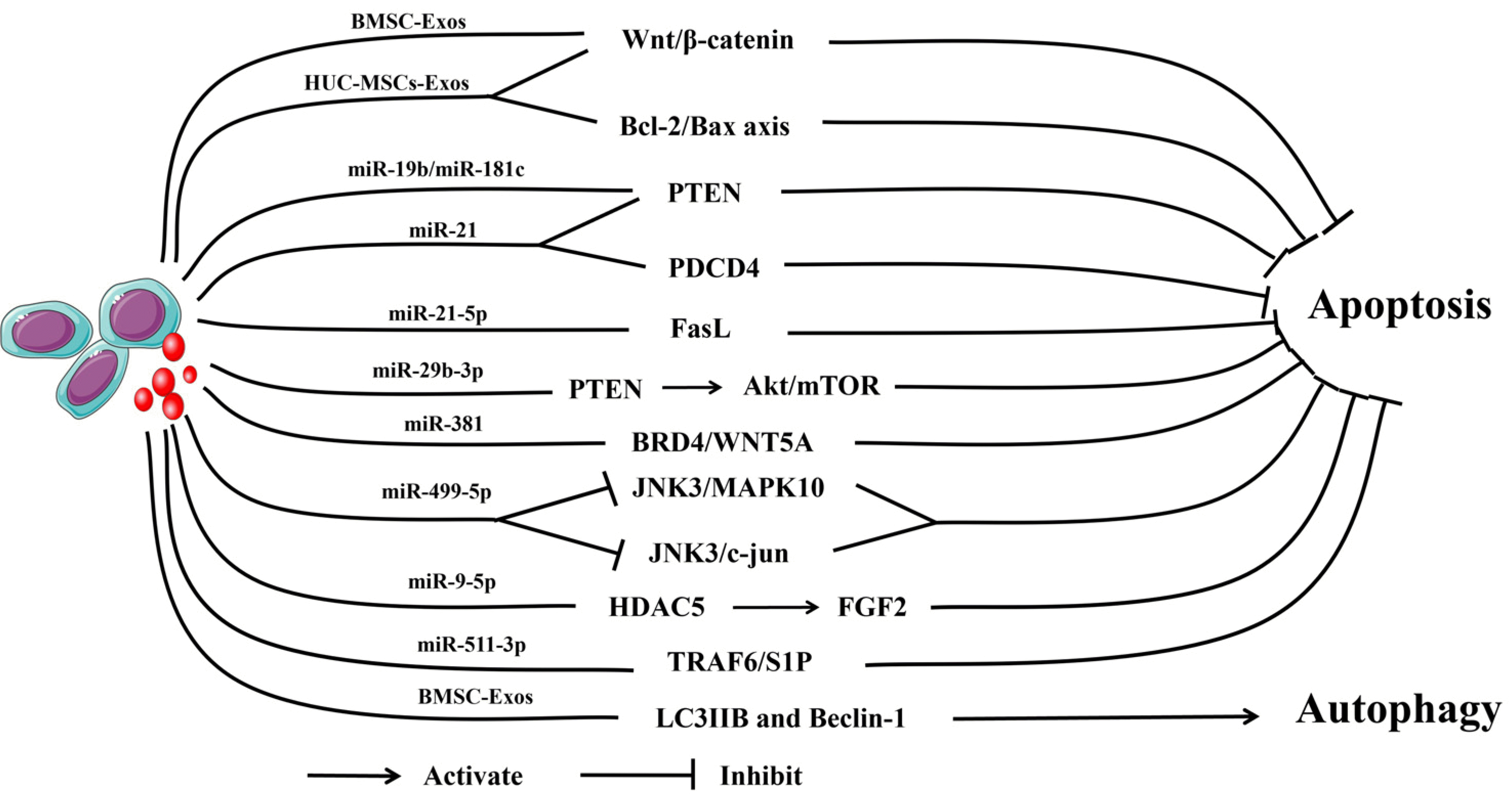 | Fig. 5The mechanism by which mesenchymal stem cells-derived exosomes (MSCs-Exos) regulates apoptosis and autophagy. Bone marrow stem cell (BMSC)-Exos can inhibit apoptosis by regulating the Wnt/β-catenin signaling pathway. human umbilical cord (HUC)-MSCs-Exos can regulate the Wnt/β-catenin signaling pathway and the Bcl-2/Bax axis to inhibit apoptosis. MSCs-Exos with miR-19b and miR-181c can inhibit apoptosis via the Phosphatase and Tensin homolog (PTEN) signaling pathways. MSCs-Exos with miR-21 can inhibit apoptosis via the PTEN and PDCD4 signaling pathways. MSCs-Exos with miR-21-5p and miR-29b-3p can inhibit apoptosis by regulating the FasL signaling pathway and activating the Akt/mTOR signaling pathway. MSCs-Exos with miR-381 can inhibit apoptosis via BRD4/WNT5A signaling pathways. MSCs-Exos with miR-499-5p inhibits the JNK3/MAPK10 and JNK3/c-jun signaling pathways to inhibit apoptosis. MSCs-Exos with miR-9-5p and miR-511-3P could regulate the HDAC5/FGF2 and TRAF6/S1P signaling pathways to inhibit apoptosis. MSCs-Exos could regulate the autophagy-related proteins LC3IIB and Beclin-1 to promote autophagy.
|
In addition, some miRNAs from exosomes have similar findings. Xu et al. (117) found that miR-21 and miR-19b derived from MSCs-Exos could reduce the expression of PTEN in a rat model of SCI, and further studies confirmed that they could regulate the apoptosis and differentiation of neuronal cells by regulating PTEN expression. Kang et al. (118) similarly found that miR-21 derived from MSCs-Exos inhibited the apoptosis of neurons in SCI rats by regulating programmed cell death 4 protein (PDCD4) and PTEN expression. Zhou et al. (119) suggested that miR-21-5p was one of the most highly expre-ssed miRNAs in MSCs-Exos, which could downregulate the expression of the proapoptotic target gene FasL and attenuate apoptosis in the injured spinal cord. Xiao et al. (120) reported that the expression of miR-29b-3p was upregulated in the spinal cord tissue of SCI rats treated with MSCs-Exos. Treatment of SCI rats with MSCs-Exos transfected with miR-29b-3p inhibitor significantly increased apoptosis in the spinal cord tissue. Further research found that its possible mechanism is to activate the Akt/mTOR pathway through the miR-29b-3p/PTEN axis to promote neurological function recovery in rats. Jia et al. (121) showed that miR-381 derived from MSCs-Exos reverses neuronal apoptosis by downregulating the BRD4/WnT5A axis and can promote recovery from SCI in a rat model. A recent study found that miR-499-5p derived from hypoxia-induced adipose mesenchymal stem cells-derived exosomes (AD-MSCs-Exos) may promote SCI repair by inhi-biting JNK3/MAPK10 and negatively regulating the JNK3/c-jun signaling pathway to regulate neuronal apoptosis (122). Furthermore, He et al. (123) explored exosomal miR-9-5p on SCI progression in an in vitro or in vivo model and discovered that exosomal miR-9-5p derived from BMSC inhibited the expression of histone deacetylase 5 (HDAC5), deacetylated the FGF2 gene and decreased its expression, thereby inhibiting FGF2-mediated apoptosis, inflammation, and endoplasmic reticulum stress to alleviate SCI in rats. Similarly, Huang et al. (124) revealed through in vitro and in vivo experiments that miR-511-3p from hypoxia-induced AD-MSCs-Exos can inhibit neuronal apoptosis and inflammatory reactions by regulating the TRAF6/S1P axis and weakening SCI progression. Thus, MSCs-Exos and their miRNA can regulate autophagy and apoptosis to improve the outcome of SCI.
Regulation of BSCB permeability by exosomes
The BSCB is the most important interface for the exchange of molecules between the blood and spinal cord parenchyma; it is mainly responsible for maintaining the normal function of the nervous system and is regulated by neurovascular unit cells (125). The BSCB is composed of the basement membrane, pericytes, capillary endothelial cells, and astrocyte foot processes (126). In particular, pericytes, as a part of the neurovascular unit, are very important for maintaining the integrity and function of blood vessels and the BSCB (127). Current studies have shown that pericytes can maintain the stability of microvessels, possibly through the following three mechanisms: promoting the expression of endothelial tight junction proteins, regulating vesicle transport and body flow across cells, and moderating the tightness connection arrangement (128). Following SCI, the structural disruption and increased permeability of the BSCB can lead to secondary injuries such as spinal cord edema, hemorrhage, oxidative stress, and excessive inflammation (127). Therefore, maintaining the integrity of the BSCB after SCI may be a potential therapeutic approach.
Some studies have identified that exosomes can improve BSCB integrity (Fig. 4). Lu et al. (129) provided the first evidence that BMSC-Exos could inhibit pericyte migration via suppression of the activation of the NF-κB signaling pathway, thereby maintaining the integrity of the BSCB and promoting functional recovery after SCI. Zhou et al. (130) showed that BMSC-Exos effectively suppresses pericyte pyroptosis and maintain BSCB integrity. Recently, another study found that MSCs-Exos increases the expression of cell junction proteins via the TIMP2/MMP pathway, which ultimately attenuates the destruction of the BSCB and improves functional recovery after SCI (131). In addition, Nakazaki et al. (132) suggested that MSCs-Exos were able to target M2 macrophages to upregulate TGFβ, TGFβ receptors, and tight junction proteins, thereby reducing BSCB permeability. Thus, MSCs-Exos can improve BSCB integrity to improve the outcome of SCI.
Promotion of neuronal regeneration and axonal growth by exosomes
Axonal damage and neuronal apoptosis usually occur during the secondary injury period of SCI (10). Due to the weak ability of neurons and axons to regenerate, they are almost unable to regenerate after injury, which ultimately leads to a large degree of limited functional recovery of the spinal cord (133). Therefore, promoting axonal growth and neuronal regeneration is one of the directions of SCI treatment.
Some studies have verified that exosomes can promote axonal growth and neuronal regeneration (Fig. 6). Early researchers found that MSCs-Exos could promote the growth of axons in neurons, but the mechanism was unclear (134, 135). Recently. Zhou et al. (136) reported that HP-MSCs-Exos can promote motor function recovery after SCI through endogenous neural progenitor cell activation and neurogenesis, a process that may involve activation of the MEK/ERK/CREB signaling pathway. Li et al. (137) provided that Exosomes derived from lipopolysaccharide-precondi-tioned MSCs can promote M2 macrophage polarization by inhibiting the NF-κb/NLRP3 signaling pathway, which ultimately promotes axon regeneration and myelin regene-ration. Wang et al. (138) found that miR-199a-3p/145-5p derived from HUC-MSCs-Exos target Cblb and Cbl mRNAs, inhibit the ubiquitination-mediated degradation of TrkA, and ultimately mediate the NGF/TrkA signaling pathway to promote neurite outgrowth and functional recovery in rats. Han et al. (139) revealed that MSCs-Exos upregulate receptor-regulated Smad 6 expression and promote neuronal regeneration in SCI rats by carrying TGFβ. Wang et al. (140) showed that MSCs-Exos containing miR-let-7a-5p can downregulate SMAD2 expression by inhibiting high-mobility Group A2 (HMGA2) and ultimately promote neuronal regeneration and improve neurological recovery through the HMGA2/SMAD2 axis in SCI rats. Thus, MSCs-Exos and their miRNA can promote axonal growth and neuronal regene-ration to improve the outcome of SCI.
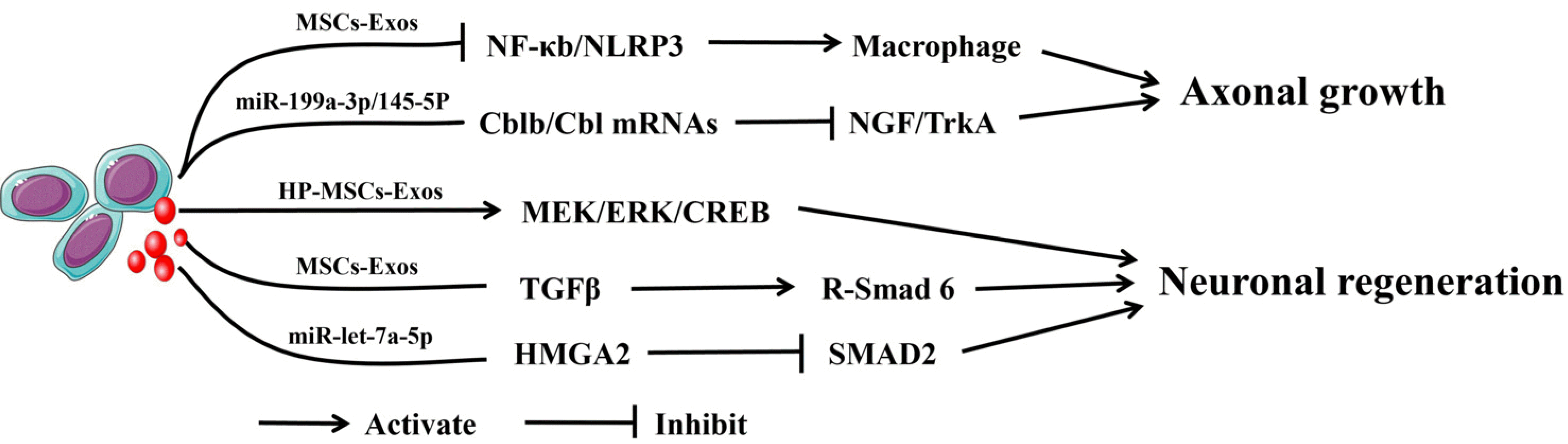 | Fig. 6The mechanism by which mesenchymal stem cells-derived exosomes (MSCs-Exos) promotes axonal growth and neuronal regeneration. MSCs-Exos can promote axonal growth by promoting M2 macrophage polarization by inhibiting the NF-κb/NLRP3 signaling pathway. MSCs-Exos with miR-199a-3p/145-5p can target Cblb/Cbl mRNAs to inhibit the NGF/TrkA pathway to promote axonal growth. Human placenta (HP)-MSCs-Exos can promote neuronal regeneration by activating the MEK/ERK/CREB signaling pathway. MSCs-Exos could attenuate R-Smad 6 expression to promote neuronal regeneration by carrying TGFβ. MSCs-Exos with miR-let-7a-5p downregulated SMAD2 expression by inhibiting HMGA2 to promote neuronal regeneration.
|
Exosomes as delivery vehicles for the transport of cargo
Recent studies have shown that exosomes not only have innate therapeutic potential but also serve as drug-delivery vehicles (141). The commonly used drug delivery carriers are multifunctional nanomaterials, including polymer nanoparticles, micelles, liposomes, mesoporous silica nanoparticles, microneedles, and polymer vesicles. Not only are nanomaterials potentially systemically toxic and immunogenic, but there are still barriers to accurate delivery to in vivo target sites (142). Compared with other carriers, exosomes have the following advantages: low cytotoxicity, deep tissue penetration, rapid clearance from the phagocytic system, and penetration of the blood-brain barrier (53). Various methods (electroporation, sonication, coincubation with modified cargo) have been developed to load proteins, siRNA, miRNA, and small molecule compounds into exosomes, which are subsequently delivered to the animal via intravenous, intraperitoneal, subcutaneous, and topical injections to produce therapeutic effects (143). Studies have found that exosomes carrying miRNAs can repair SCI; thus, exosomes can be used as cargo transporters, which is another new method of SCI treatment.
At present, some experiments have proven that exosomes carrying miRNA have great potential in the treatment of SCI. Li et al. (144) discovered that miR-544-modified MSCs-Exos improve functional recovery and neuronal survival by reducing the levels of proinflammatory cytokines after SCI. Li et al. (145) identified that loaded with miR-133b, MSCs-Exos preserved neuronal cells and enhanced the regeneration of axons, which was attributed to the activation of ERK1/2, STAT3, and CREB, as well as to the inhibition of RhoA expression. MSCs-Exos containing miR-126 not only inhibits the expression of SCI-related EVH1 domain protein 1 and phosphoinositide 3 kinase regulatory subunit 2 to promote angiogenesis after SCI but also has apoptotic effects (146). Ren et al. (147) discovered that miR-133b-modified AD-MSCs-Exos can affect the signaling pathway related to axon regeneration, NF, GAP43, GFAP, and MBP and promote the recovery of neurological function in SCI animals. Similarly, miRNA-29b loaded in BMSC-Exos repaired SCI in rats by regulating proteins involved in neuronal regeneration, such as NF200, GAP-43, and GFAP (148). Overexpression of miR-338-5p in BMSC-Exos provided neuroprotection and inhibited apoptosis after SCI in vivo and in vitro. Further studies indicated that the possible mechanism is the activation of the PI3K/Akt pathway through cAMP-mediated Rap1 activation (149). Chen et al. (150) and Liu et al. (151) found that loaded with miR-455-5p, BMSC-Exos could alleviate SCI, increase autophagy to inhibit neuronal apoptosis and promote recovery of locomotor function after SCI by downregulating Nogo-A. Chen et al. (152) verified that loaded with miR-26a, BMSC-Exos could activate the PTEN-AKT-mTOR pathway to promote axonal regeneration and neurogenesis and attenuate glial scarring in SCI. Sheng et al. (153) reported that MSCs-Exos loaded with miRNA-22 can inhibit the pyroptosis of microglia and the neuroinflammatory response after SCI and ultimately improve the neurological function of rats. Huang et al. (154) showed that MSCs-Exos loaded with miR-494 can suppress inflammatory responses and resist apoptosis by promoting the polarity of macrophages and increasing the levels of the anti-apoptotic protein Bcl-2. Last, Jiang and Zhang (155) suggested that MSCs-Exos loaded with miR-145-5p reduces inflammation in SCI by regulating the TLR4/NF-κB signaling pathway. The above large number of animal experiments has confirmed the great potential of exosome-carrying miRNAs in the treatment of SCI. Thus, MSCs-Exos can carry external miRNA to improve the outcome of SCI.
Go to :
Discussion
Acute SCI is a severe CNS injury that has high morbidity and mortality. Due to the complex structure of the CNS and the poor regeneration ability of neurons, the treatment of SCI is still a worldwide problem. Previous studies have shown that MSCs may be a new research direction for the treatment of SCI, and a large number of animal studies have shown that stem cells can promote SCI repair through tissue repair and replacement, angiogenesis, and neurotrophic, anti-apoptotic, and anti-inflammatory mechanisms, but MSCs treatment also has many safety problems, such as thrombosis and embolism, infection, immune rejection, and tumorigenicity (11). As research progressed, it was found that the efficacy of MSCs appeared to be related to the exosomes they secreted. Exosomes have the advantages of small size, good biocompatibility, low toxicity, and low immunogenicity, which suggests that exosome therapy is superior to stem cell therapy (156). More importantly, exosomes can transport genetic material, protect it from extra-cellular degradation, penetrate the blood-brain barrier, and act on receptor cells with high selectivity, making them an ideal delivery system for regenerative medicine (157). In recent years, MSCs-Exos have achieved satisfactory results in clinical studies in the treatment of ischemic stroke, macular degeneration, diabetes, and kidney disease (43). Although there are currently no clinical studies on exosomes and SCI, a large number of animal studies in this review demonstrate that exosomes have neuroprotective properties as a treatment for SCI, paving the way for future clinical studies on exosomes. In addition, after SCI, miRNAs related to oxidative stress, inflammation, apoptosis, and other secondary injuries are significantly differentially expressed, resulting in differential expression of target genes and changes in cell function, which makes us pay more attention to the role of miRNAs in SCI. The possible therapeutic mechanisms and effects of some miRNAs in SCI have been discovered thus far, but the direct mechanisms of more miRNAs are still unclear, so further exploration is needed, and miRNAs are expected to become a key component of SCI repair. Moreover, MSCs-Exos are promising cell-free drug delivery systems for delivering therapeutic agents, such as miRNAs, to target tissues in animal models, enabling individualized therapy through proteins in miRNA-regulated pathways.
Despite the progress of exosomes in the treatment of SCI, there are still many challenges to be faced. On the one hand, the separation, purification, and amplification of exosomes need to be solved comprehensively. There is an urgent need for rapid, inexpensive, simple, and standardized isolation and purification procedures to produce high-purity, high-yield exosomes with intact biological acti-vity. On the other hand, current SCI models are mainly rats or mice. Due to species and anatomical differences, the human nervous system is more complex than rodents, and recovery after SCI is more difficult. Therefore, it is important to clarify the experimental efficacy of exosomes in primates, which have a more complex and advanced nervous system, to provide a further theoretical basis for the next clinical study. In addition, exosomes are generally not targeted, and it remains challenging to efficiently translocate them to the site of the lesion and to function in vivo for a long period. Recent studies have identified tissue-engineered scaffolds and bioengineering approaches to assist exosome delivery, which not only improve exosome targeting but also facilitate long-term sustainment and slow release of exosomes in vivo (158). Therefore, further search for efficient tissue engineering and bioengineering techniques can help exosomes to function better.
Go to :
Summary
In conclusion, MSCs-Exos and their miRNAs, as new therapeutic methods for SCI, showed special advantages. Current animal research has clarified that MSCs-Exos and their miRNAs in the treatment of SCI, including regulating immune responses, promoting angiogenesis, activating autophagy and inhibiting apoptosis, regulating the permeability of BSCB, promoting neuronal regeneration and axonal growth, and acting as delivery carriers. However, there are still many challenges, including exploring more efficient methods for isolation, purification, and amplification of exosomes, clarifying the role of exosomes in different animal models of SCI, and finding efficient tissue engi-neering and bioengineering techniques to improve the efficacy of exosomes. Ultimately, we believe that MSCs-Exos offers new possibilities for the treatment of SCI, and we expect that relevant clinical trials will be able to be conducted in the future, ultimately benefiting all of humanity.
Go to :
 XML Download
XML Download