

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is a result of an imbalance between energy intake and energy expenditure due to a sedentary lifestyle coupled with overnutrition and is commonly associated with other metabolic disorders such as type 2 diabetes mellitus (T2DM), hypertension, cardiovascular disease, and even cancers [12]. In 2022, NCD Risk Factor Collaboration reported that the combined prevalence of underweight and obesity in adults was over 10% in all countries, with 89% (women) and 73% (men) countries having higher obesity prevalence than underweight [3]. According to the 2016 epidemiology of obesity prevalence by the World Health Organization (WHO), almost 60% of adults and nearly one-third of children in Europe were overweight or suffered from obesity [4]. A study of adult obesity in Shanghai, China from 2016 to 2017 reported that the prevalence of general obesity and central obesity were 13.5% and 28.9%, and anticipates that this figure will continue to grow in the future [5]. Obesity represents a major health challenge as it significantly increases the risk of chronic diseases contributing to a decline in quality of life and posing a serious socio-economic burden [6]. The current treatment for obesity is primarily focused on controlling energy intake and expenditure, such as dietary restrictions, weight loss surgery, medications that reduce food absorption, and increasing physical activity, etc. [7]. However, the efficacy of lifestyle and behavioral interventions is poor; surgical intervention usually expensive and not widely available; weight loss drugs typically have dependency and cardiovascular side effects [8]. These side effects largely limit their therapeutic effect on obesity and its related metabolic diseases [9]. Therefore, there is an urgent need to find new therapies to combat obesity and related metabolic diseases from a new perspective.
The WHO defines obesity as a condition that is characterized by abnormal or excessive fat accumulation and is associated with increased health risks. Individuals with a body mass index (BMI) > 30 kg/m2 are considered obese [10]. The adipose tissue plays a central role in energy storage and has a key regulatory role in body metabolism. Adipose tissue in mammals can be categorized into 2 subtypes: white adipose tissue (WAT) and brown adipose tissue (BAT). WAT mainly stores extra energy and also serves as an endocrine organ [11], while BAT utilizes nutrients for thermogenesis under the action of the uncoupling protein 1 (UCP1) [12]. UCP1, the signature molecule located in the inner mitochondrial membrane of BAT, can catalyze the leak of protons across the mitochondrial inner membrane, dissipating the electrochemical proton gradient generated during electron transport chain (ETC) activity, which would otherwise drive ATP production by ATP synthase. By doing so, UCP1 dissipates the proton gradient, converting energy into heat instead of ATP production [13]. In adults, the amount of BAT is negatively correlated with BMI and age, and positively correlated with the resting metabolic rate [14]. Meanwhile, several basic research studies have confirmed that improving the activity of BAT by various means can increase the body's thermogenic capacity and prevent the progression of metabolic conditions such as high fat-induced obesity, hyperlipidemia, impaired glucose tolerance, and insulin resistance [1516]. The above evidence has led to a renewed understanding of the role of brown fat in adult metabolic diseases, and its study as a new therapeutic target in obesity and associated diseases has become the focus of attention.
Adenosine-5’-monophosphate (AMP)-activated protein kinase (AMPK) is a highly conserved serine/threonine protein kinase that is expressed in almost all eukaryotes [17]. AMPK acts as a sensor for cellular energy metabolism and is activated when intracellular energy levels are low or when the utilization of adenosine triphosphate (ATP) is high to promote ATP production by increasing the activity or expression of proteins involved in catabolism [18]. Increased AMPK activity is also associated with brown fat activity. Studies have found that AMPK knockdown in mice have decreased non-shivering thermogenesis in BAT [19]. This suggests that AMPK may regulate brown adipose activity in humans [20]. Also, AMPK in adipose tissue has a significant role in maintaining browning levels in vivo.
Cryptotanshinone (CT) is a bioactive compound isolated from the root of the herb Salvia miltiorrhiza with anti-inflammatory effects and is used to treat metabolic disorders in Chinese medicine [21]. Studies have shown that CT can regulate lipid metabolism [22]. Terado et al. [23] found that CT suppresses tumorigenesis by inhibiting adipogenesis. Notably, CT has been found to play a significant role in promoting the commitment of mesenchymal stem cells (MSCs) to the brown adipocyte lineage and enhancing mitochondrial biogenesis via AMPK signaling [24]. However, the role of CT in increasing the level of browning in vivo through the activation of AMPK to increase body thermogenesis, promote fat consumption, and reduce body weight remains to be further elaborated.
Therefore, in this study, we aimed to observe the effects of CT on the level of fat browning in mice on a high-fat diet. We further investigated the specific molecular mechanism of CT in increasing brown fat activity and promoting white fat browning through AMPK at the cellular level to offer new ideas for obesity prevention and treatment.
MATERIALS AND METHODS
Procurement of animals
Thirty male C57BL/6J mice (20 ± 2 g, 4 weeks) were purchased from Shanghai SLAC Laboratory Animal Co., Ltd., Animal Production License No.: SCXK (Shanghai, China) 2017-0005. The mice were reared at an air humidity of 60 ± 10% and a temperature of 22 ± 3°C. The experimental procedures on the animals were approved by the Animal Experimentation Ethics Committee of Zhejiang Eyong Pharmaceutical Research and Development Center (approval number: ZJEY-20221013-01).
Establishment and grouping of the animal models
After 2 weeks of adaptive feeding, the mice were randomly divided into 4 groups: normal diet (ND) group, high-fat diet (HF) group, CT-200 group, and CT-400 group, with 6 rats in each group. The ND group was fed a normal diet throughout the study. The mice in the HF, CT-200, and CT-400 groups were provided with a high-fat diet for 8 weeks from the 6th week of age. Subsequently, the 14-week-old mice of the HF, CT-200, and CT-400 groups received saline, 200 mg/kg/d CT (B21586; Yuanye, Shanghai, China) and 400 mg/kg/d CT via gavage for 6 weeks, respectively [2526]. We weighed the mice at 6, 10, 14, and 20 weeks of age.
Intraperitoneal glucose tolerance test (IPGTT)
After 6-week CT intervention, mice undergo a 12-h fasting. Then, we measured their basal blood glucose. At 8:00 a.m., we take blood from the tail tip to check baseline blood glucose level (0 min). Then, the glucose solution was injected intraperitoneally. Blood glucose was measured and recorded at 30, 60, 90, and 120 min, and the change in the blood glucose with time was plotted.
Intraperitoneal insulin tolerance test (IPITT)
One day after IPGTT, the tail tips were weighed and clipped. Basal blood glucose was measured after discarding the first drop of blood. Blood glucose levels were recorded at 0, 30, 60, 90, and 120 min after intraperitoneal injection of recombinant human insulin. The blood glucose curve was plotted and analyzed over time.
Cold stimulation test
The anal temperature of the mice was measured at room temperature. Subsequently, the mice were placed in a cold room at 4°C, and the anal temperature was measured and recorded at 30, 60, 90, and 120 min. During the test period, the mice were fasted by depriving them of food and water. They were then transferred to room temperature as soon as possible after the end of the test. Cold stimulation test starts at 48 h after the end of IPGTT.
Blood and tissue sample collection
After the above experiments, the blood was collected from the eyes of the mice and centrifuged at 3,500 r/min after 1 h. The serum was separated and used for the analysis of the serum lipids including total cholesterol (TC), triglycerides (TG), and low-density lipoprotein cholesterol (LDL-C) using a Hitachi 3100 automatic biochemical identification instrument (Hitachi, Tokyo, Japan). After euthanasia, mice were dissected to obtain subcutaneous adipose tissue from the groin and interscapular BAT. These were weighed and fixed in a formalin solution or stored in liquid nitrogen for subsequent use.
Hematoxylin and eosin (H&E) staining
The subcutaneous adipose tissue and BAT of each group of mice were washed, fixed in 4% paraformaldehyde, dehydrated, and embedded in paraffin. The paraffin sections were treated in xylene and gradient ethanol solution in turn and then placed in hematoxylin (H3136; Sigma-Aldrich, St. Louis, MO, USA) and eosin (E4009; Sigma-Aldrich) solution for 3–5 min. After dehydration and sealing of adipose tissues, the tissue morphology was observed under a light microscope (Nikon Eclipse Ci-L; Nikon, Tokyo, Japan). The ImageJ (NIH, Bethesda, MD, USA) software was used to analyze the diameter of the adipocytes.
Immunohistochemical analysis
Adipose tissue paraffin sections were dewaxed, hydrated, and then incubated with 100 μL of 3% H2O2 for 10 min. Sections were placed in an antigen repair solution and incubated with 5% bovine serum albumin (B2064; Sigma-Aldrich) solution for 20 min. Fifty μL of primary and secondary antibodies were added in sequence, respectively, followed by the introduction of diaminobenzidine (P0203, Beyotime, Shanghai, China) for color development after sufficient reaction. The sections were re-stained with hematoxylin, dehydrated, transparentized, and then observed under the Nikon Eclipse Ci-L microscope (Nikon).
Extraction and cell culture of primary MSCs
The ND group mice were euthanized and disinfected by immersion in 75% ethanol for 5 min. The adipose tissue at the bilateral groin and epididymis was removed under aseptic conditions. Vascular and fibrous components and muscle tissues were removed in the culture dish. The adipose tissue was then made into a paste and transferred to a Collagenase Digester (LS004176; Worthington Biochemical, Lakewood, NJ, USA) for 45 min, and low sugar Dulbecco’s Modified Eagle Medium (DMEM) (11054-001; Gibco, Grand Island, NY, USA) with 10% fetal bovine serum was added. After centrifuging at 2,000 rpm for 10 min, cell precipitates were obtained and added to 2 mL of DMEM medium for resuspension. The cells were incubated in a CO2 incubator (BB150; Thermo Scientific, Waltham, MA, USA); the first half-volume fluid change was performed after 48 h, followed by half-volume fluid changes once every 2–3 days. The morphological changes of the primary and passaged cells were observed under an inverted microscope (AE2000; Motic, Xiamen, China).
Oil Red O staining
MSCs were inoculated in 6-well plates with 3 × 105 cells per well. Cells were fixed with 4% paraformaldehyde for 15 min and washed with 60% isopropanol for 3 min. Subsequently, they were washed with phosphate-buffered saline (PBS) thrice and Oil Red O working solution (G1260; Solarbio, Beijing, China) was added. After staining overnight, they were observed under the microscope (AE2000; Motic).
Alizarin red staining
MSCs were inoculated in 6-well plates and osteogenic induction conditioned medium containing 50 μg/mL L-ascorbic acid (ST1434; Beyotime), 10 mM sodium β-glycerophosphate (G9422-10G; Sigma-Aldrich) and 10-8 M dexamethasone (D1756-25MG; Sigma-Aldrich) was added. The medium was changed every 3 days, and the cells were incubated at 37°C in a 5% CO2 incubator. After the cells were fixed, 0.3% TritonX-100 (SH30042.01; Hyclone, Logan, UT, USA) was added to permeabilize the cells. They were stained with 500 μL of alizarin red solution (ST1078; Beyotime) for 20 min and then photographed and recorded.
Flow cytometric analysis
After adipose MSCs were made into a 107 cells/mL single-cell suspension by 100 μL cell buffer. Then 2 μL of primary antibodies were added for detecting the cluster of differentiation (CD) 90 (553016; BD, Franklin Lakes, NJ, USA), CD73 (561544; BD), CD29 (102216; BioLegend, San Diego, CA, USA), CD34 (560238; BD), CD45 (552848; BD), CD11b/c (561688; BD), mixed and incubated for 30 min at 2–8°C. After low-speed centrifugation, the supernatant was discarded and incubated with 100 μL buffer and 2 μL of fluorescein isothiocyanate-labeled fluorescent secondary antibody protected from light. Samples were assayed after buffer resuspension using a BD Accuri C6 flow cytometer (BD).
Cell grouping and intervention
Acadesine (AICAR, HY-13417) is an AMPK agonist, and dorsomorphin (Compound C, HY-13418A) is an AMPK inhibitor. They were purchased from MedChemExpress (Monmouth Junction, NJ, USA). The adipose MSCs were randomly divided into 4 groups to verify that AMPK is a key target for CT: the MDI group (0.5 mM 1-methyl-3-isobutylxanthine, 1 μM dexamethasone, and 10 μg/mL insulin were used to induce the adipocyte differentiation of MSCs) [24], the CT group (MDI + 8 μmol/L CT) was used to observe the effect of CT on adipocyte differentiation, the AICAR group (MDI + 8 μmol/L CT + AICAR 200 μM) [27], and the Compound C group (MDI + 8 μmol/L CT + Compound C 4 μM). The AMPK agonists group (AICAR) and AMPK inhibitors group (Compound C) were compared with the CT group to demonstrate that CT regulates lipid differentiation by targeting AMPK.
Immunofluorescence staining
Cells were inoculated onto sterile coverslips and treated as described above in groups after cell apposition. Cells were fixed in 4% paraformaldehyde (P0099; Beyotime) and permeabilized by 0.5% Triton X-100. After blocking, a Bodipy™ stain (agci70160; Amgicam, Wuhan, China) was added, and the cells were incubated for 15 min. Excess stain was washed off with PBS, then 1 μg/mL diamidino-2-phenylindole (C1005; Beyotime) was added to color the nuclei, and the fluorescence intensity was observed using Ts2-FC inverted microscopy (Nikon).
Quantitative real-time polymerase chain reaction (qRT-PCR)
We extracted the total mRNA from adipose tissue or cells, and cDNA was generated using a reverse transcription kit (CW2569; CWBIO, Taizhou, China). The reverse transcription reaction conditions were 42°C, 15 min; 85°C, 5 min. Real-time fluorescence quantitative polymerase chain reactions were implemented according to the operating instructions of the qRT-PCR kit (11201ES08; YEASEN, Shanghai, China). The primer sequence information is listed in Table 1.
Table 1
Primer sequence information
PGC-1α, proliferator-activated receptor-gamma coactivator-1α; UCP1, uncoupling protein 1; CIDEA, cell death inducing DFFA like effector A; Prdm16, prdomain-containing 16; Tmem26, transmembrane protein 26; PPARγ, peroxisome proliferator-activated receptor γ; CD, cluster of differentiation; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Western blot
Radio immunoprecipitation assay (P0013B; Beyotime) lysates were used to extract the total proteins from adipose tissue and MSCs. A bicinchoninic acid assay kit (P0012; Beyotime) was used to determine the concentration of each group of proteins. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used to separate the target bands. After transferring and closing the membrane, it was incubated overnight in the primary antibodies of p-protein kinase B (AKT) (AF0016; Affinity, San Francisco, CA, USA), AKT (AF6259; Affinity), p-hormone-sensitive triglyceride lipase (HSL) (AF8026; Affinity), HSL (AF6403; Affinity), UCP1 (DF7720, DF7720), Complex III (DF7720; Abcam, Cambridge, UK), Complex V (ab176569; Abcam), p-AMPK (AF3423; Affinity), AMPK (AF6423; Affinity) with a dilution of 1:1,000. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (1:10,000, AF7021; Affinity) and β-actin (1:10,000, AF7018; Affinity) served as internal references. After the addition of the secondary antibody (7074/7076; CST, Danvers, MA, USA) and adequate antigen-antibody reaction, electrochemiluminescence (ECL) was used for assessing chemiluminescence development.
Statistical analysis
The data analysis was carried out using the SPSS statistical software (version 25.0; IBM, Armonk, NY, USA). The data conformed to a normal distribution and met the χ2 test. One-way analysis of variance (ANOVA) was used for data comparison between multiple groups, followed by Tukey’s test. The GraphPad Prism (version 7.0; GraphPad Software, Boston, MA, USA) software was used for data presentation and analysis of the 2-way ANOVA, followed by Tukey’s test. All data were expressed as mean ± SD. P < 0.05 was considered a statistically significant difference.
RESULTS
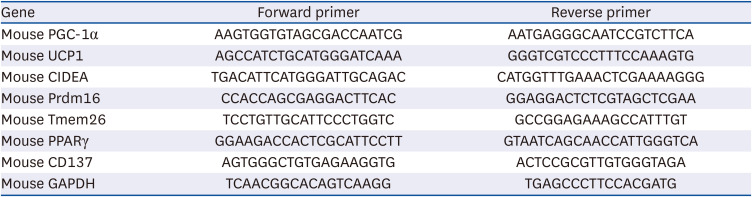
CT reduces body weight and improves glucose intolerance and insulin resistance in mice
We weighed the mice at 6th, 10th, 14th, and 20th weeks of age. An increase in body weight was observed in all the mice on a high-fat diet, other than the ND group which was fed a normal diet at the 10th and 14th week of age (P < 0.05; Fig. 1A). After 6 weeks of saline intervention, the body weights of mice increased in the HF group (P < 0.01; Fig. 1A) with a significant increase in food intake (P < 0.01; Fig. 1B). The 3 groups on a high-fat diet have similar food intake (P > 0.05; Fig. 1B). Compared with the HF group, mice with the CT-200 and CT-400 interventions for 6 weeks showed significant weight loss (P < 0.01; Fig. 1A). In the IPGTT and IPITT tests, blood glucose fluctuations in the HF group are both significantly higher than those in the ND group with notably increased IPGTT and IPITT area under the curve (AUC) (P < 0.01; Fig. 1C-F). Compared with the HF group, blood glucose fluctuations of mice in the CT-200 and CT-400 groups are significantly lower with decreased IPGTT and IPITT AUC (P < 0.01; Fig. 1C-F).
Fig. 1
CT reduces body weight and improves glucose intolerance and insulin resistance in mice. (A) Mice body weight and (B) food intake were examined (n = 6). (C) IPGTT was used to examine the effect of CT on glucose tolerance in mice on HF group. (D) AUC was calculated for IPGTT (n = 6). (E) IPITT was used to evaluate the effect of CT on insulin tolerance in mice on HF group (n = 6). (F) AUC was calculated for IPITT (n = 6). Data from A, B, D and F were analyzed by 1-way ANOVA. Data from C and E were analyzed by 2-way ANOVA.
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; IPGTT, intraperitoneal glucose tolerance test; AUC, area under the curve; IPITT, intraperitoneal insulin tolerance test; ANOVA, analysis of variance.
*P < 0.05, **P < 0.01 vs. ND; ##P < 0.01 vs. HF.
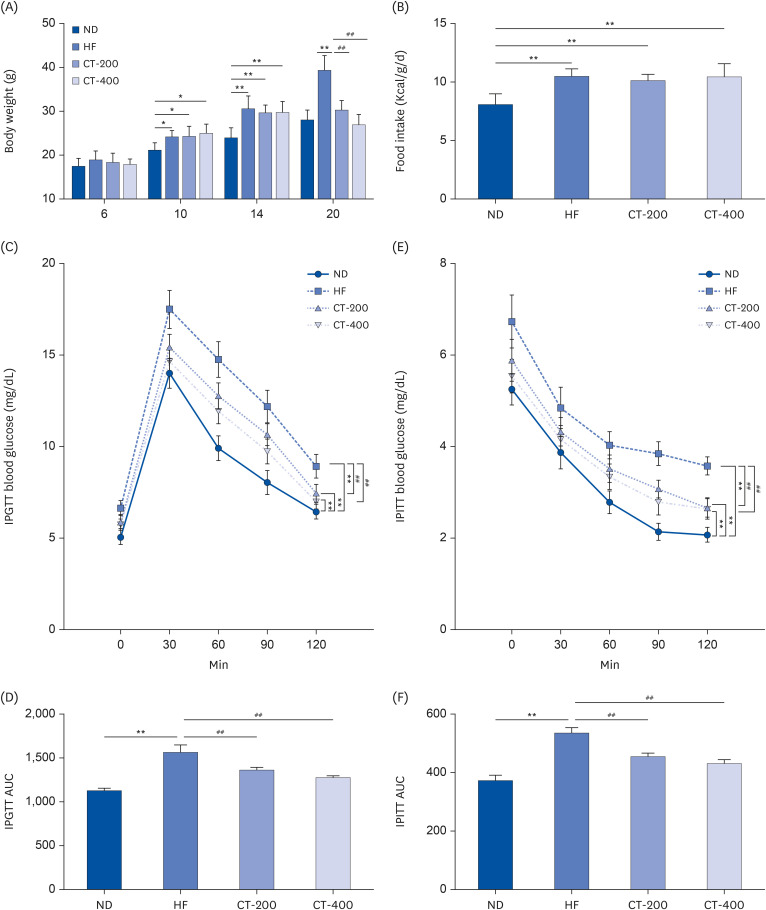
CT increases adaptive thermogenesis and suppresses dyslipidemia
The cold stimulation test showed that the anal temperature in the HF group increased at 30, 60, 90, and 120 min compared with the ND group (P < 0.01; Fig. 2A). Compared with the HF group, the anal temperature of mice in the CT-200 group increased at 120 min, and that of mice in the CT-400 group increased at 30, 60, 90 and 120 min (P < 0.01; Fig. 2A). The levels of TC, TG, and LDL-C in the serum of mice in the HF group increased compared with those in the ND group (P < 0.01; Fig. 2B-D). The levels of TC, TG, and LDL-C in the serum of mice in the CT-200 and CT-400 groups decreased compared with the HF group (P < 0.01; Fig. 2B-D).
Fig. 2
CT increases adaptive thermogenesis and suppresses dyslipidemia. (A) The cold stimulation test was used to examine the regulation of body temperature in mice by CT and the data were analyzed by 2-way ANOVA (n = 6). (B-D) Serum TC, TG, and LDL-C levels were measured in each group of mice, and the data were analyzed by 1-way ANOVA (n = 6).
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; ANOVA, analysis of variance; TC, total cholesterol; TG, triglycerides; LDL-C, low-density lipoprotein cholesterol; ANOVA, analysis of variance.
**P < 0.01 vs. ND; ##P < 0.01 vs. HF.
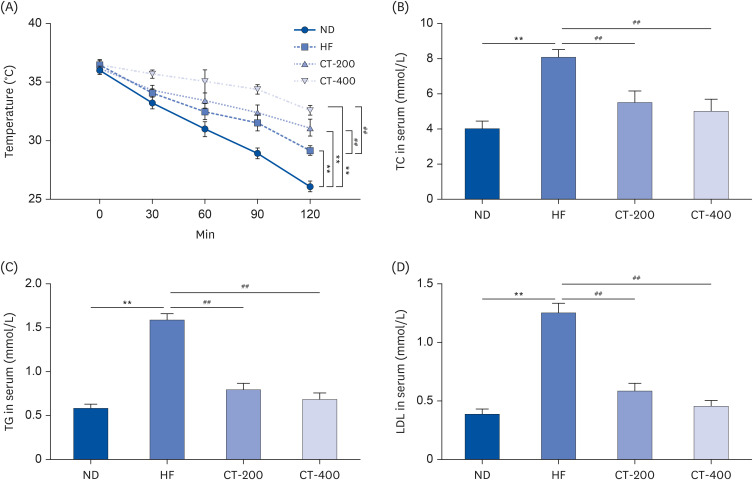
CT improved adipose tissue morphology and promoted UCP1 levels
The H&E staining results showed that WAT and BAT in the ND group were normal with tightly arranged cells and no obvious damage (Fig. 3A). Compared to the ND group, the HF group exhibited enlarged adipocytes with loosely arranged cells and severe damage in both WAT and BAT. Conversely, the severity of damage was milder and adipocyte size was smaller in the CT-200 and CT-400 groups compared to the HF group (P < 0.01; Fig. 3A and B). Immunohistochemistry revealed that the expression of UCP1 in the WAT and BAT of the HF group was lower than that of the ND group (P < 0.01) and that the CT intervention increased the levels of UCP1 (P < 0.05; Fig. 3B and C).
Fig. 3
CT improved adipose tissue morphology and promoted UCP1 levels. (A) Morphological effects of CT on WAT and BAT in mice as observed by H&E staining (magnification, 200×). (B) Average adipocyte diameter for quantitative analysis. (C, D) Immunohistochemistry was used to detect the effect of CT on UCP1 expression levels in WAT and BAT (magnification, 200×), and the data were analyzed by 1-way ANOVA (n = 3).
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; UCP1, uncoupling protein 1; WAT, white adipose tissue; BAT, brown adipose tissue; H&E, hematoxylin and eosin; ANOVA, analysis of variance.
**P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
CT promotes brown gene expression and improves insulin sensitivity and lipolysis in adipose tissue
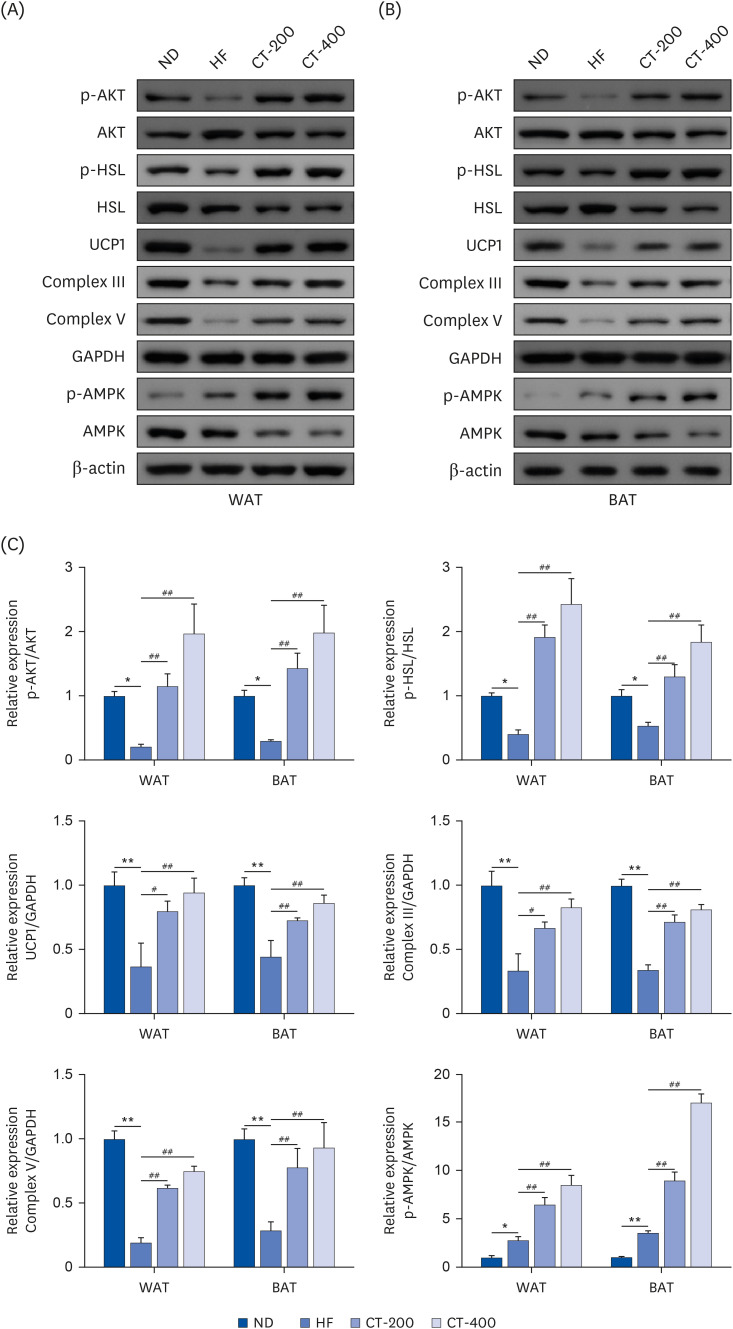
A qRT-PCR was used to examine the effect of CT on brown gene expression in WAT and BAT. Compared with the ND group, the expressions of peroxisome proliferator-activated receptor-gamma coactivator-1α (PGC-1α), UCP1, cell death inducing DFFA like effector A (CIDEA), prdomain-containing 16 (Prdm16), transmembrane protein 26 (Tmem26), peroxisome proliferator-activated receptor γ (PPARγ), and CD137 mRNA were reduced in the adipose tissue of the HF group (P < 0.05), while CT-200 and CT-400 reversed the above changes (P < 0.05; Fig. 4). The western blot showed that the protein expressions of p-AKT/AKT, p-HSL/HSL, UCP1 Complex III, and Complex V were lower in the HF group than in the ND group (P < 0.05), while p-AMPK/AMPK was increased (P < 0.05; Fig. 5). The levels of all the proteins increased after the CT intervention (P < 0.05; Fig. 5).
Fig. 4
CT promotes brown gene expression in adipose tissue. qRT-PCR was used to examine the effect of CT on the brown genes (A) PGC-1α, (B) UCP1, (C) CIDEA, (D) Prdm16, (E) Tmem26, (F) PPARγ, and (G) CD137 mRNA levels in the WAT and BAT tissues. Data were analyzed by one-way ANOVA (n = 3).
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; qRT-PCR, quantitative real-time polymerase chain reaction; PGC-1α, proliferator-activated receptor-gamma coactivator-1α; UCP1, uncoupling protein 1; CIDEA, cell death inducing DFFA like effector A; Prdm16, prdomain-containing 16; Tmem26, transmembrane protein 26; PPARγ, peroxisome proliferator-activated receptor γ; CD, cluster of differentiation; WAT, white adipose tissue; BAT, brown adipose tissue; ANOVA, analysis of variance; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
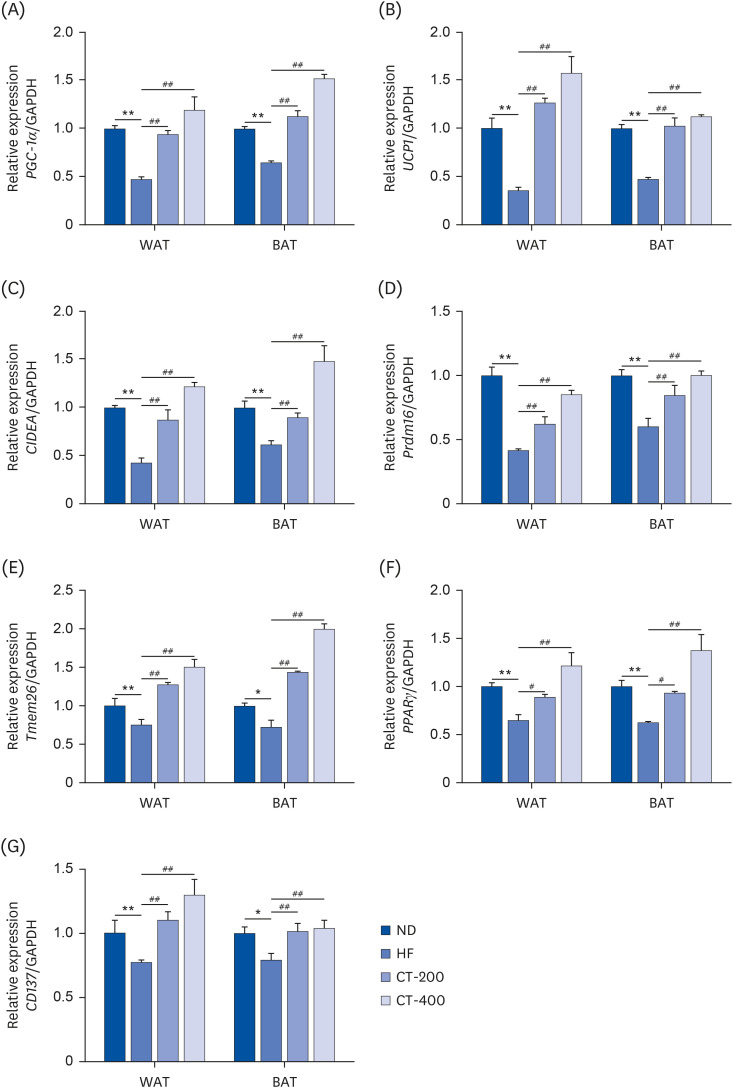
Fig. 5
CT promotes brown gene protein expression and improves insulin sensitivity and lipolysis in adipose tissue. Western blot was used to detect the effect of CT on protein expression levels of p-AKT/AKT, p-HSL/HSL, UCP1, Complex III, Complex V, and p-AMPK/AMPK in (A) WAT and (B) BAT. (C) qRT-PCR was used to examine the effect of CT on protein expression levels above. Data were analyzed by one-way ANOVA (n = 3).
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; AKT, protein kinase B; HSL, hormone-sensitive triglyceride lipase; UCP1, uncoupling protein 1; AMPK, adenosine-5'-monophosphate (AMP)-activated protein kinase; WAT, white adipose tissue; BAT, brown adipose tissue; ANOVA, analysis of variance.
*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
Primary adipose MSCs were successfully isolated and identified
After 24 h of isolation and culture of mouse adipose tissue MSCs, the cells were short and spindle-shaped. The cells grew in clusters and the cell colonies gradually expanded at day 3. At day 4, the cells gradually fused and grew in a fibrillar, vortex-like pattern (Supplementary Fig. 1A). Oil Red O and alizarin red staining were used to detect the lipogenic and osteogenic differentiation abilities of the MSCs (Supplementary Fig. 1B and C). The morphology of cells induced by lipogenic differentiation changed from shuttle-shaped to oval-shaped with intracellular aggregates of lipid droplets of various sizes. Some of the intracellular lipid droplets were immaturely differentiated, with smaller lipid droplets in the form of rings, while the lipid droplets in the cells with mature differentiation were larger, and after Oil Red O staining, the lipid droplets were red (Supplementary Fig. 1B). The cells inducing osteogenic differentiation were locally aggregated and formed mineralized crystals, and small black granules of different sizes appeared in the cytoplasm. Mushroom-like scattered spherical nodules were visible after alizarin red staining, and the spherical nodules increased in size and changed their morphology to a square, rhombic decagonal shape after differentiation and maturation (Supplementary Fig. 1C). In addition, flow cytometry confirmed that adipose MSCs expression of high CD90, CD73, CD29 and low CD34, CD45, CD11b/c (Supplementary Fig. 1D).
CT inhibits lipid droplet formation and mRNA levels of brown genes in MSCs through AMPK activation
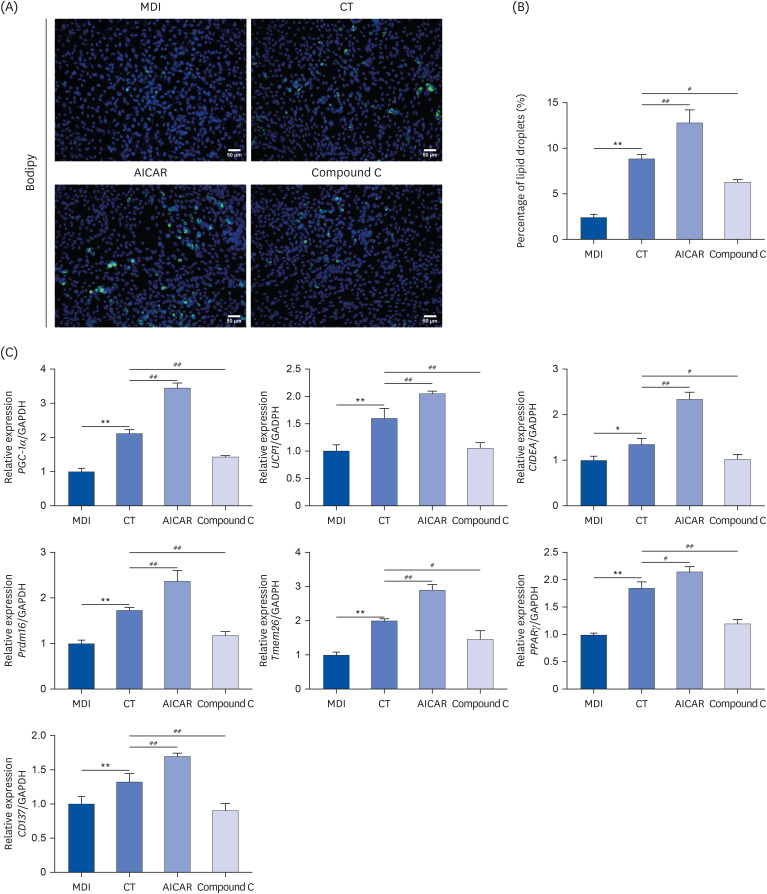
Immunofluorescence was used to detect lipid droplet formation in each group of adipose MSCs (Fig. 6A and B). The larger, single lipid droplets were mostly seen in the MDI group, while a large number of small lipid droplets, partially fused around the nucleus, were seen in the CT, AICAR, and Compound C groups (P < 0.05; Fig. 6A and B). Compared with the MDI group, the expressions of PGC-1α, UCP1, CIDEA, Prdm16, Tmem26, PPARγ, and CD137 mRNA were elevated in the cells in the CT group (P < 0.05). AICAR promoted the effect of CT (P < 0.05), while Compound C reversed the above changes (P < 0.05; Fig. 6C).
Fig. 6
CT inhibits lipid droplet formation and mRNA levels of brown genes in MSCs through activation of AMPK. (A, B) Immunofluorescence assays were used to detect lipid droplet formation in MSCs (magnification, 200×). (C) qRT-PCR was used to examine the effect of CT on the brown genes PGC-1α, UCP1, CIDEA, Prdm16, Tmem26, PPARγ, CD137 mRNA levels in MSCs. Data were analyzed by one-way ANOVA (n = 3).
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; MSC, mesenchymal stem cell; AMPK, adenosine-5'-monophosphate (AMP)-activated protein kinase; qRT-PCR, quantitative real-time polymerase chain reaction; PGC-1α, proliferator-activated receptor-gamma coactivator-1α; UCP1, uncoupling protein 1; CIDEA, cell death inducing DFFA like effector A; Prdm16, prdomain-containing 16; Tmem26, transmembrane protein 26.
PPARγ, peroxisome proliferator-activated receptor γ; CD, cluster of differentiation; ANOVA, analysis of variance.
*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
CT promotes brown gene expression and improves insulin sensitivity and lipolysis in adipose tissue through AMPK activation
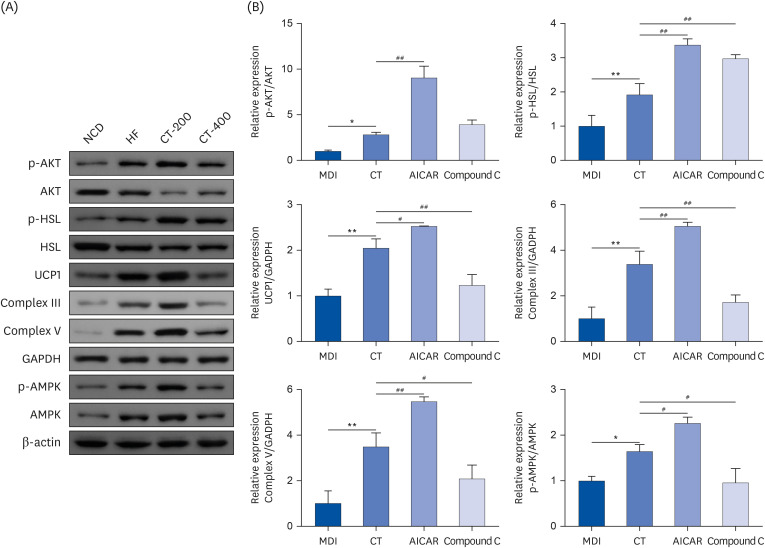
The western blot suggested that the protein expressions of p-AKT/AKT, p-HSL/HSL, UCP1, Complex III, Complex V, and p-AMPK/AMPK were elevated in the cells of the CT group compared with the MDI group (P < 0.05) and that the expressions of the above molecules increased in the AICAR group compared with those in the CT group (P < 0.05; Fig. 7). However, Compound C reversed the effect of CT (P < 0.05; Fig. 7).
Fig. 7
CT promotes brown gene expression and improves insulin sensitivity and lipolysis in adipose tissue through activation of AMPK. (A, B) Western blot was used to detect the effect of CT on protein expression levels of p-AKT/AKT, p-HSL/HSL, UCP1, Complex III, Complex V, p-AMPK/AMPK in MSCs. Data were analyzed by one-way ANOVA (n = 3).
Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).
CT, Cryptotanshinone; AMPK, adenosine-5'-monophosphate (AMP)-activated protein kinase; AKT, protein kinase B; HSL, hormone-sensitive triglyceride lipase; UCP1, uncoupling protein 1; MSC, mesenchymal stem cell; ANOVA, analysis of variance.
*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
DISCUSSION
This study found that CT promotes adipose tissue browning to increase body thermogenesis and thus prevent obesity. The underlying mechanism involved the activation of the AMPK pathway. In addition, animal and cellular experiments have confirmed that CT promotes brown gene expression in the adipose tissue and improves insulin sensitivity and lipolytic activity. “Browning of WAT” means that WAT can transform into beige adipocytes with morphological and functional characteristics similar to BAT, which plays a significant role in preventing obesity, inflammation, and diabetes [1628]. Earlier studies have reported that CT can inhibit inflammatory reactions in adipocytes [29] and inflammatory cell infiltration in T2DM [30]. Our research supports the overall role of CT in promoting browning and reducing inflammation.
Several studies have found that natural compounds, especially flavonoids, can promote thermogenesis by increasing the level of browning in the body [3132]. In our study, CT, an abietane type-diterpene quinone pigments from Salvia miltiorrhiza [33], significantly reduced body weight, blood glucose, and insulin resistance in obese mice. This is consistent with previous studies and further confirms the efficacy of CT in obesity-related metabolic disorders [2134]. In addition, our experimental results showed that the anal temperature of mice in the CT groups was significantly higher than that of mice in the HF group under cold stimulation, suggesting that CT could increase the adaptive thermogenesis of obese mice. Also, brown adipose and subcutaneous adipocyte volume and lipid droplets were reduced in the CT groups compared to the HF group. Through immunohistochemistry, qRT-PCR, and western blot, we further demonstrated that CT increased the expressions of UCP1 and other brown genes and promoted insulin sensitivity and lipolytic activity. Earlier studies have reported that CT treatment promoted brown-adipocyte-like features, including the increased gene expression of brown-fat signature genes and beige-cell-specific genes [24]. In this study, we observed that CT increased the expression of UCP1, CIDEA, Prdm16, PGC-1α (brown-fat signature genes), Tmem26, and CD137 (beige-cell-specific genes) in adipose tissue. Specifically, Prdm16 is a key molecule in BAT formation and regulates adipose biogenesis [35]. Additionally, high UCP1 expression in adipocytes dissipates energy through the mobilization of mitochondria [36]. To sum up, our study suggests that the ameliorative effects of CT on metabolism are achieved by activating BAT and the browning of WAT in vivo, but the exact molecular mechanism remains to be further investigated.
Studies have shown that AMPK activation is associated with brown fat activity and white fat browning [37]. It has been reported that palmitic acid and chicoric acid increase the activity of BAT and promote white fat “browning” by activating the AMPK pathway [3839]. In addition, AMPK promotes the survival of β cells, and inhibits insulin resistance, promotes glucose metabolism and uptake and its activation has been shown to ameliorate diabetic complications [40]. AMPKα1 deficient mice fed on a high-fat diet have been shown to gain weight and develop impaired glucose tolerance [41]. Aerobic training improves the markers of non-alcoholic fatty liver disease (NAFLD) and insulin resistance via the regulation of the AMPK-PPAR-α signaling pathway in obese mice [42]. AMPK is a key target for improving obesity.
Nagappan et al. [43] found that CT improved ethanol-induced liver injury through the AMPK pathway. The role of CT in browning has also been elucidated by other researchers, who found that CT may enhance brown-adipocyte-like features and increase mitochondrial mass by phosphorylating AMPKα [24], suggesting a close relationship between CT and AMPK. In both in vivo and in vitro experiments, we also found that CT increased AMPK activity in BAT and WAT tissues, while this effect was mitigated by Compound C, an AMPK inhibitor. Furthermore, the browning-promoting effect of CT was enhanced using the AMPK agonist AICAR. Our study supports the idea that CT promotes the browning of WAT and the activities of BAT to alleviate obesity, which is mediated through the activation of the AMPK pathway.
Our study has certain limitations. Results from the in vitro experiments support the premise that AMPK may be linked with the CT regulation of adipocyte browning. MDI was used in our study to induce the adipocyte differentiation of MSCs [44]. Researchers have found that the adipocyte differentiation model with MDI incubation shows both brown and beige cell-like characteristics [45]. This indicate that although MDI is a commonly used model, it may not be the most suitable one. Nevertheless, further studies using diverse models are required to clarify the exact role of CT and AMPK and uncover any additional crucial pathways.
In conclusion, this study combined in vivo and in vitro experiments to demonstrate that CT increases brown fat activity and promotes the “browning” of subcutaneous fat through AMPK activation, thereby increasing thermogenesis and energy expenditure and improving obesity-related elevated glucose and insulin resistance. This study provides a scientific basis for the treatment of obesity with CT.
 XML Download
XML Download