

 PDF
PDF Citation
Citation Print
Print



Introduction
Bone remodeling is a dynamic process, finely orchestrated by osteoblasts and osteoclasts, maintaining a balance between bone formation and resorption [1, 2]. Any disruption to this equilibrium can lead to bone loss and the gradual deterioration of skeletal microstructure, ultimately resulting in osteoporosis—a condition characterized by bone fragility and increases susceptibility to fractures [3, 4]. In menopausal women, estrogen deficiency disrupts the natural bone turnover process, further exacerbating the risk of osteoporosis [5].
Vitronectin (VN), an extracellular matrix (ECM) protein, is abundantly present in serum, liver, cancellous bone, and the mineralized matrix of cortical bones [6, 7]. It is recognized for its considerable interactions within the mineralized bone matrix, hinting at its role in the regulation of bone metabolism [7]. Numerous studies have highlighted the multifaceted role of VN-derived peptides in bone remodeling, actively contributing to both bone formation and resorption processes. In vivo investigations have demonstrated that VN-derived peptides accelerate osteoblast differentiation and activity, thereby promoting bone formation [8]. In vitro studies have also shown that VN peptides support osteoblast differentiation. Furthermore, a specific VN-derived peptide has been observed to enhance osteoblastogenesis while simultaneously inhibiting osteoclastogenesis, as well as suppressing the expression of receptor activator of NF-κB ligand (RANKL) and interleukin-17A in vivo [9]. Additionally, VN has been found to act as a protective shield against bone degradation by inhibiting the secretion of cathepsin K by activated osteoclasts [10]. Conversely, VN induces tartrate-resistant acid phosphatase (TRAP) release by osteoclasts, accompanied by distinctive features like ruffled borders and clear zones that are characteristic of resorbing osteoclasts [11]. These collective findings suggest that VN might serve as a crucial endogenous modulator of bone remodeling. However, despite the existing body of research on the effects of VN on bone remodeling, its precise in vivo role remains largely ambiguous.
In this study, we aimed to elucidate the impact of VN on osteoporosis and bone remodeling in VN-deficient ovariectomy (OVX) mice.
Go to : 
Materials and Methods
Animals
Wild-type (WT) C57BL/6J female mice (8 weeks old) were acquired from Charles River Laboratory in Kanagawa, Japan. Vitronectin knockout (VNKO) female mice were generously supplied by Dr. David Ginsburg at the University of Michigan. All animal experiments received approval from the Institutional Animal Care and Use Committee of Ochanomizu University, Japan (Animal protocol nos. 21023R and 23024), and were conducted in accordance with the guidelines established by the Ministry of Education, Science, and Culture in Japan.
Ovariectomized mice
Eight-week-old female mice underwent ovariectomy or sham surgery using bilateral ovariectomy to establish an experimental mouse model of menopause. The procedure was performed under anesthesia, employing a mixture of 0.75 mg/kg Domitor (Nippon Zenyaku Kogyo), 4 mg/kg midazolam (Sandoz), and 5 mg/kg Vetorphale (Meiji Seika Pharma). A small abdominal incision was made, and the oviduct and ovarian blood vessels were tied and sectioned for ovariectomy. Sham-operated mice underwent a similar procedure, excluding the sectioning of the oviducts and removal of the ovaries [12].
Bone metabolism marker measurements
Plasma was obtained from blood by centrifugation and stored at –20°C. Alkaline phosphatase (ALP) and TRAP activities in plasma were measured using a TRACP & ALP Assay Kit MK301 (Takara Bio Inc.) following the procedures detailed in the manufacturer’s instructions.
Bone histomorphometry
Bone histomorphometry was conducted in accordance with the method previously described by Ikegami et al. [13]. In brief, femurs were fixed overnight at 4°C using 4% paraformaldehyde. Subsequently, the fixed specimens underwent decalcification in a 10% EDTA solution (pH 7.4) for a duration of 3 days at 4°C. Following cryoprotection with 20% sucrose, femur sections were obtained at a thickness of 5 μm using cryostats (Leica CM1850; Leica Microsystems).
For ALP staining, sections were incubated at 37°C for 20 minutes in a 0.1 M Tris-HCl buffer (pH 8.5) containing naphthol AS-MX phosphate and Fast Blue BB Salt hemi (zinc chloride) salt (Merck). Meanwhile, for TRAP staining, sections were exposed to a 0.1 M sodium acetate buffer (pH 5.0) containing 50 mM sodium tartrate, naphthol AS-MX phosphate, and Fast Red Violet LB Salt (Merck) at room temperature for 25 minutes. Methyl green was used for counterstaining. Subsequently, slides were examined under a light microscope (BZ-X700; Keyence). To assess bone formation and resorption, the ratio of osteoblast perimeter to bone perimeter (Ob.Pm/B.Pm) and the number of osteoclasts (N.Oc) per B.Pm were measured in an area located 1.0 mm from the growth plate in the proximal metaphysis of the femur.
Immunofluorescence staining of the femurs
Tissue sections of murine femurs were prepared as described in the bone histomorphometry section. The primary antibodies used were anti-VN (1:200 dilution; 50585-R019; Sino Biological Inc.), anti-tartrate-resistant acid phosphatase type 5 (TRACP5) (1:200 dilution; NBP2-4529; Novus Biologicals), and anti-runt related transcription factor 2 (RUNX2, 1:200 dilution; 692802; BioLegend) antibodies. The secondary antibodies used were as follows: Alexa Fluor 488 goat anti-rabbit immunoglobulin G (IgG), Alexa Fluor 568 goat anti-mouse IgG, and Alexa Fluor 568 goat anti-rat IgG (all from Thermo Fisher Scientific). Additionally, 4’,6-diamidino-2-phenylindole dihydrochloride (Roche Diagnostics) was used for nuclear staining. Images were captured with a confocal microscope (LSM700; Carl Zeiss).
Western blot analysis
To detect VN, the protein in the serum was quantified using a bicinchoninic acid protein assay kit, the amount was adjusted to 50 μg, and subjected to electrophoresis on a 10% polyacrylamide gel and transferred onto polyvinylidene difluoride membranes. The membranes were blocked with 5% skim milk and incubated with primary anti-VN antibodies (1:5,000 dilution; gifted by Prof. Masao Hayashi, Ochanomizu University) overnight, and then with horseradish peroxidase-labeled secondary antibodies for 1 hour each. The bands were visualized with an enhanced luminescent reagent (Ez WestLumi; ATTO) and captured using the iBright CL1500 Imaging System (Thermo Fisher Scientific). The density of each band was quantified using the ImageJ software (National Institutes of Health).
X-ray micro-computed tomography analysis of femurs
The femurs were subjected to cone-beam X-ray micro-computed tomography (CT) scanning (CosmoScan FX; Rigaku) under the following conditions: a tube voltage of 90 kV, a tube current of 88 μA, a slice thickness of 10 μm, and a voxel size of 10 μm. Three-dimensional bone parameters were computed using 3-dimensional image analysis software (Analyze 12.0; AnalyzeDirect). To analyze the distal femurs, we selected a region of interest consisting of 100 slices containing both cortical and trabecular bone, beginning from the end of the growth plate. The following parameters were employed for scanning the femurs:
For trabecular bone: bone volume (BV), total volume (TV), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp), trabecular number (Tb.N), marrow area (Ma.Ar), degree of anisotropy (DA), and the structural model index (SMI).
For cortical bone: cortical bone area (Ct.Ar), total cross-sectional area (Tt.Ar), cortical thickness (Ct.Th), and the ratio of Ct.Ar to Tt.Ar were determined.
Statistical analyses
Detailed information regarding sample size and the statistical tests used for each experiment is provided in the manuscript. In general, statistical significance was determined by the P-values obtained through a one-way ANOVA followed by the Tukey–Kramer test or Student’s t-test using Prism Software 9.00 (GraphPad Software). Significance levels were defined as follows: P<0.05, P<0.01, P<0.001, and P<0.0001.
Go to :
Results
Effect of ovariectomized on body weight and uterus weight
To investigate the effect of VN on osteoporosis, we performed an OVX operation. Female mice were subjected to OVX or sham surgery at 8 weeks of age and were euthanized 4 weeks post-surgery (Fig. 1A). We recorded the initial body weight of each mouse on the day of ovariectomy and subsequently measured their body weights weekly for both the OVX and sham-operated groups during the 4 weeks post-operation period. The body weights of OVX-operated mice were significantly higher than those of sham-operated mice after surgery (P<0.05) (Fig. 1B). Furthermore, we recorded the uterine weight of each mouse in both the OVX and sham-operated groups, and the OVX-operated mice exhibited significant uterine tissue atrophy compared to the sham-operated mice (P<0.0001), confirming the success of the surgical procedure (Fig. 1C).
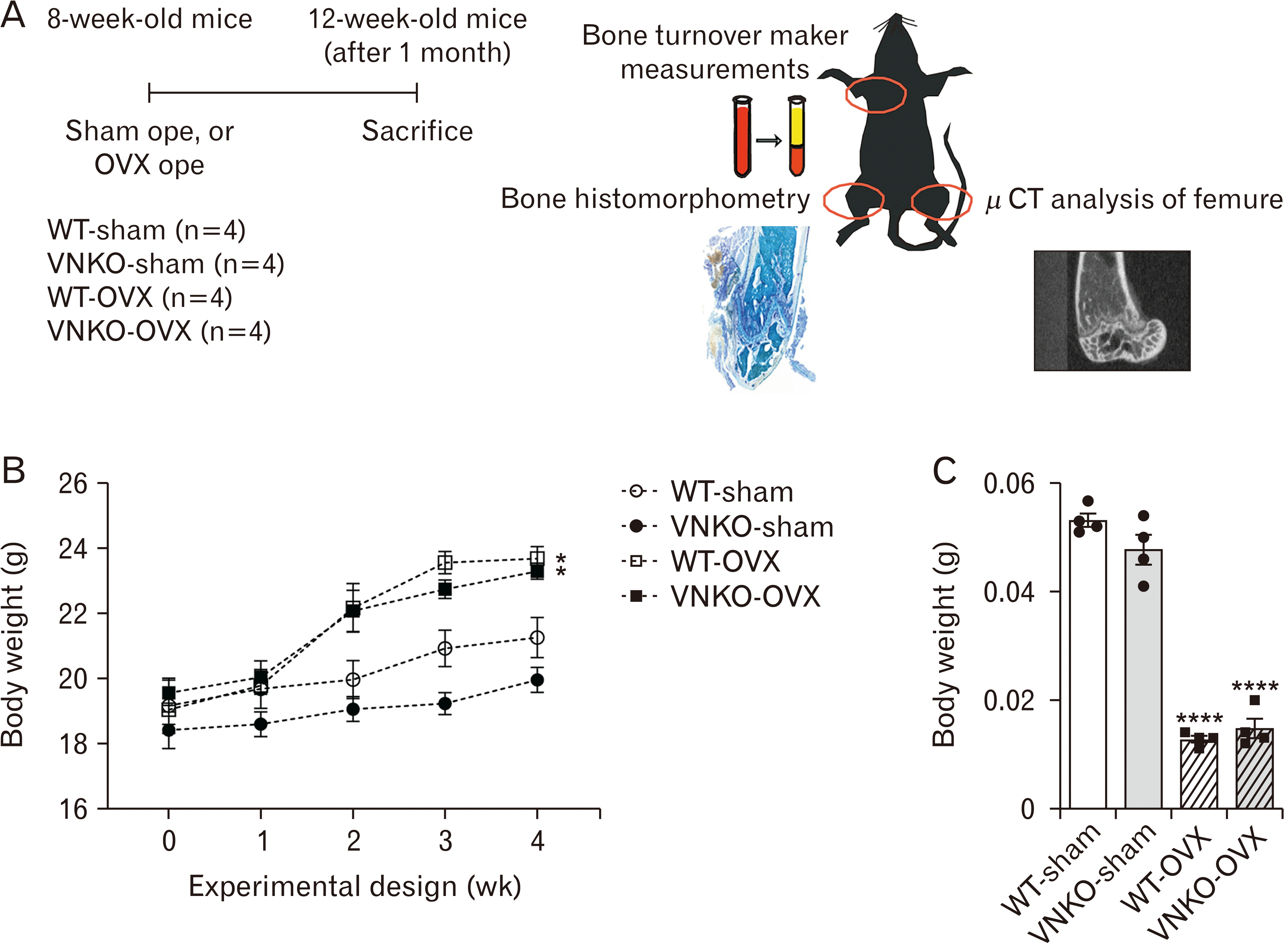 | Fig. 1Effects of ovariectomy on the body and uterine weights of ovariectomized (OVX) mice. (A) Experimental design of this study. Wild-type (WT) female C57BL/6J mice and vitronectin (VN) knockout (KO) female mice were randomly divided into four groups (n=4 mice/group), namely sham-operated WT group (WT-sham), sham-operated VNKO group (VNKO-sham), OVX WT group (WT-OVX), and OVX+VNKO group (VNKO-OVX). The sham and OVX operations were performed in 8-week-old mice, which were sacrificed at 4 weeks after surgery. Plasma was separated from blood by centrifugation and stored at –20°C. The right femur was stored at –20°C for micro-computed tomography (micro-CT) analysis, whereas left femur was fixed in 4% paraformaldehyde at 4°C overnight and subjected to histomorphometry. (B) Body weight change in sham-operated or OVX-operated mice. Body weight was measured in WT-sham, VNKO-sham, WT-OVX, and VNKO-OVX. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. sham-operated group. (C) Uterine weights of sham- or OVX-operated mice. The uterine weight was measured in WT-sham, VNKO-sham, WT-OVX, and VNKO-OVX groups. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: ****P<0.0001 vs. sham-operated group. Dots indicate the results from each animal.
|
Expression and localization of vitronectin in the bone matrix and serum of ovariectomized mice
To confirm the localization of VN in bone remodeling and its interaction with osteoclasts and osteoblasts, we first investigated VN expression in femur sections of sham-operated and OVX-operated mice. Immunofluorescent staining was performed using anti-VN antibody, a marker of osteoclasts, TRACP5, and a marker of osteoblast, RUNX2. VN staining was localized in the bone matrix close to osteoclasts; in contrast, VN staining was detected around the osteoblasts (Fig. 2A). Our results suggest that the osteoclasts adhered to VN on the bone surface and VN is localized around the osteoblasts. The western blot analysis results showed that the expression of VN in the serum was not significantly different between sham-operated and OVX-operated WT mice (Fig. 2B). Furthermore, no immunoreactivity against VN was observed in the bone matrix and serum of VNKO mice (Fig. 2A, B).
 | Fig. 2Localization and expression of vitronectin (VN) in bone remodeling. (A) Immunofluorescent images of femur sections of sham- or ovariectomized (OVX)-operated mice. Top panels: VN (green) and tartrate-resistant acid phosphatase type 5 (TRACP5) (red) expression in the femur sections, with merged images indicating areas of overlap. The arrowheads indicate the sites where VN is close to osteoclasts. Bottom panels: VN (green) and runt related transcription factor 2 (RUNX2) (red) expression in the femur sections. 4’,6-diamidino-2-phenylindole (grey) was used for nuclear counterstaining. Scale bar: 5 μm. (B) Western blot analysis of VN expression in the sera of sham- or OVX-operated mice. Fifty micrograms of protein was loaded into each lane. Relative expression levels of VN protein in wild-type (WT) mice. Data are presented as mean±SEM, n>2. The results were analyzed using the t-test. WT-sham, sham-operated WT group; WT-OVX, OVX WT group; A.U., arbitrary unit; n.s., not significant.
|
Effect of vitronectin deficiency on bone metabolism in ovariectomized mice
To investigate the effect of VN on bone metabolism in OVX mice, we assessed the activity of ALP in osteoblasts and TRAP in osteoclasts in both sham- and OVX-operated mice (Fig. 3). Interestingly, the OVX surgery had no discernible impact on ALP activity in either WT or VNKO mice (Fig. 3A). However, we did observe a significant increase in TRAP activity in VNKO-OVX mice compared to WT-OVX mice (P=0.0445) (Fig. 3B).
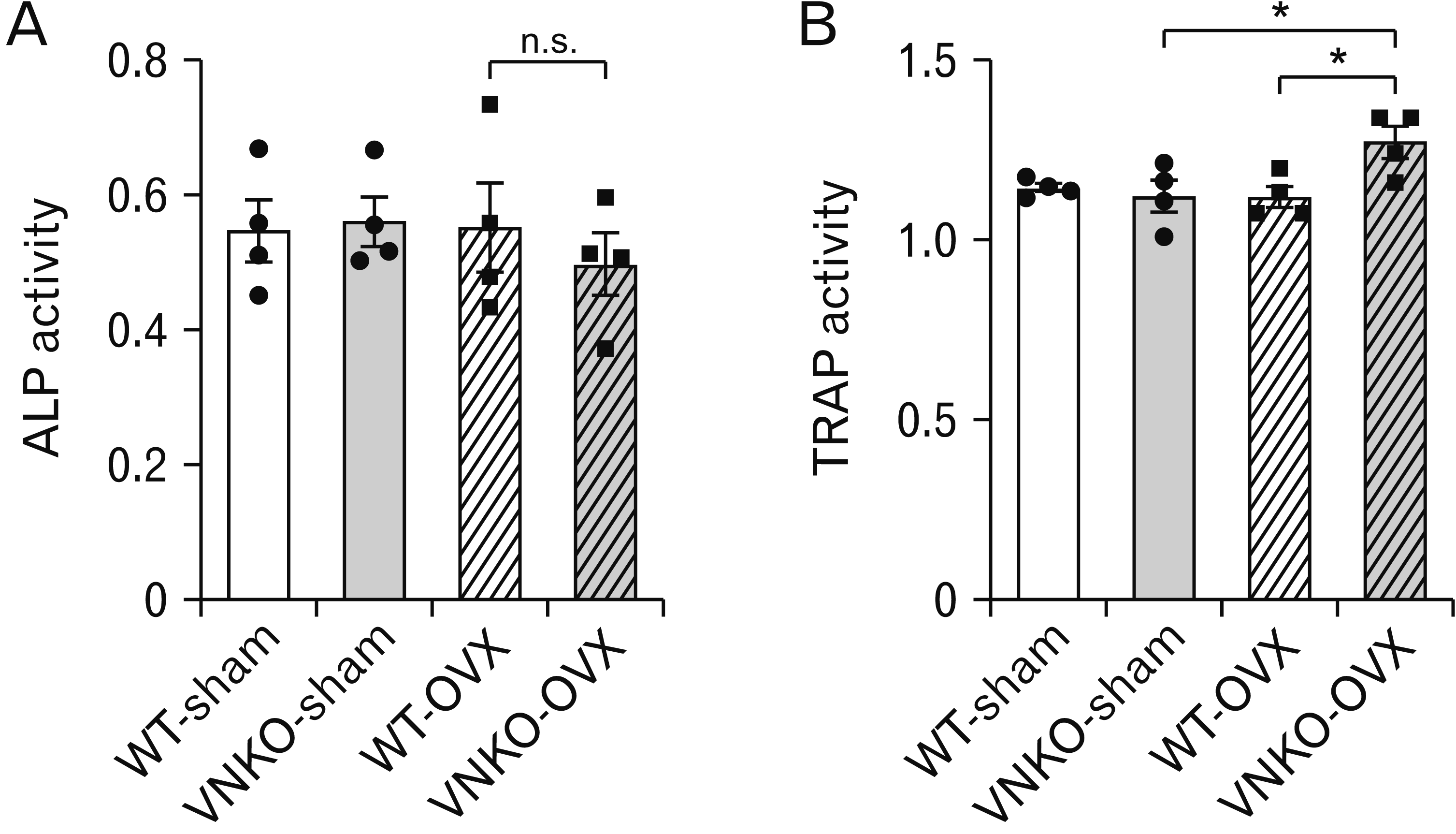 | Fig. 3Effects of vitronectin (VN) deficiency on bone metabolism markers in sham- or ovariectomized (OVX)-operated mice. (A) The activity of plasma alkaline phosphatase (ALP), marker for osteoblasts, in sham- or OVX-operated mice. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA. Dots indicate the results from each animal. (B) The activity of plasma tartrate-resistant acid phosphatase (TRAP), marker for osteoclasts, in sham- or OVX-operated mice. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. OVX+VNKO group (VNKO-OVX). Dots indicate the results from each animal. VNKO, vitronectin knockout; n.s., not significant.
|
Effect of vitronectin deficiency on osteoclastogenesis in ovariectomized mice
To confirm the effect of VN on bone histology, we conducted ALP and TRAP staining to visualize osteoblasts and osteoclasts in the femur (Fig. 4A, C). Osteoblasts were identified as ALP-positive cells that appeared blue, and we analyzed the ratio of Ob.Pm to B.Pm within a specific region located 1.0 mm distal to the growth plate in the proximal metaphysis of the femur, utilizing a light microscope (BZ-X700). As shown in Fig. 4A and B, there was no significant difference in the ratio of Ob.Pms among the groups, this is consistent with the observation that VN deficiency did not significantly affect ALP activity (Fig. 3A).
 | Fig. 4Alkaline phosphatase (ALP) and tartrate-resistant acid phosphatase (TRAP) staining of femur sections from sham- or ovariectomized (OVX)-operated mice. (A) ALP staining images of the femurs in OVX-operated groups. Scale bar: 100 μm. (B) Bone formation was assessed by measuring the ratio of ALP-positive osteoblast perimeter to bone perimeter (Ob.Pm/B.Pm) in an area 1.0 mm from the growth plate in the distal metaphysis of the femur. Data represent the mean±SEM, n=3. Statistics used one-way ANOVA. Dots indicate the results from each animal. (C) TRAP staining images of the femurs in OVX-operated groups. The arrowheads indicate the activity of osteoclasts. Scale bar: 100 μm. (D) Bone resorption was assessed by measuring the number of TRAP-positive osteoclasts (n>3 nuclei) per bone perimeter (N.Oc/B.Pm) in an area 1.0 mm from the growth plate in the distal metaphysis of the femur. Data represent the mean±SEM, n=3. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. WT-OVX, **P<0.01 vs. WT-sham or VNKO-sham. Dots indicate the results from each animal. WT-sham, sham-operated WT group; VNKO-sham, sham-operated VNKO group; WT-OVX, OVX WT group; VNKO-OVX, OVX+VNKO group; VNKO, vitronectin knockout; n.s., not significant.
|
Osteoclasts were identified as TRAP-positive cells with >3 nuclei on the bone surface [14]. The N.Oc per B.Pm in the proximal metaphysis of the femur was quantified within an area 1.0 mm from the growth plate. The OVX operation did not significantly affect the N.Oc in WT mice (Fig. 4D). However, VN deficiency in OVX-operated mice (VNKO-OVX) induced a significant increase in osteoclast number in the femur compared to any other mouse groups studied (P=0.003 for WT-sham, P=0.0017 for VNKO-sham, P=0.0192 for WT-OVX) (Fig. 4D), suggesting that VN deficiency may promote the OVX-induced osteoclastogenesis.
X-ray micro-computed tomography analysis in sham and ovariectomized groups
To investigate whether VN can regulate ovariectomy-induced osteoporosis, we analyzed the effects of VN deficiency on bone microarchitecture and biomechanical parameters in OVX- and sham-operated mice (Fig. 5A). Firstly, we confirmed the effect of OVX operation on bone microarchitecture. In micro-CT analysis, OVX-operated mouse groups (WT and VNKO) exhibited a significant increase in TV (P=0.0185 for WT, P=0.0057 for VNKO) and Ma.Ar (P=0.0162 for WT, P=0.006 for VNKO), but not in BV, in comparison to their respective sham-operated mice, indicating that the OVX operation increased the medullary cavity. Additionally, the OVX-operated groups showed a significant decrease in the ratio of Ct.Ar to Tt.Ar (P=0.0034 for WT-sham and P=0.0009 for VNKO-sham) compared to the respective sham-operated group. These results suggest that the OVX operation led to alterations in bone microarchitecture.
 | Fig. 5Micro-computed tomography (CT) analysis of femurs in sham- or ovariectomized (OVX)-operated mice. (A) Micro-CT images of the distal femurs obtained after sham- or OVX-operation. Sham-operated wild-type (WT) group (WT-sham), sham-operated VNKO group (VNKO-sham), OVX-operated WT group (WT-OVX), and OVX-operated vitronectin knockout (VNKO) group (VNKO-OVX) were compared; the trabecular bone (orange), the cortical bone (red), and the cortical pore (green). (B, C) Quantitative micro-CT analysis of femurs from sham- or OVX-operated mice. (B) The graph shows parameters of trabecular area of femurs; bone volume (BV), total volume (TV), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp), trabecular number (Tb.N), marrow area (Ma.Ar), degree of anisotropy (DA), and the structural model index (SMI). Data represent the mean±SEM, n>3. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. WT-OVX, **P<0.01 vs. VNKO-sham, †P<0.05 vs. WT-sham, ††P<0.01 vs. WT-OVX, †††P<0.001 vs. VNKO-sham. Dots indicate the results from each animal. (C) The graph shows parameters of cortical bone area (Ct.Ar) of femurs; Ct.Ar, total cross-sectional area (Tt.Ar), cortical thickness (Ct.Th), and the ratio of Ct.Ar to Tt.Ar. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: **P<0.01 vs. WT-sham, ***P<0.001 vs. VNKO-sham. Dots indicate the results from each animal.
|
Next, we examined the effects of VN deficiency on bone resorption. VN deficiency significantly suppressed the Ma.Ar and TV in both sham-operated and OVX-operated mice (Fig. 5B). We also confirmed the effects of VN deficiency on the DA and SMI. DA was increased by OVX operation, and the OVX-induced DA increase was suppressed by VN deficiency (P=0.0089) (Fig. 5B). SMI is used to determine the plate- or rod-like geometry of trabecular structures. OVX operation increased the SMI value (Fig. 5B), indicating that the OVX operation increased the plate-to-rod transition (SMI=0 for plates and 3 for rods). VN deficiency suppressed the increase in OVX-induced SMI value (Fig. 5B), suggesting that it mitigated bone resorption.
Go to :
Discussion
VN is present throughout the mineralized bone matrix of cancellous and cortical bones, suggesting its involvement in bone remodeling through bone formation and resorption [7]. Previous studies have largely focused on the exogenous effects of VN in osteoporosis and lay a crucial foundation for understanding the potential therapeutic applications of VN. However, the specific endogenous role of VN, particularly as a component of the ECM in regulating bone remodeling and resorption, remains poorly understood. In this study, we aimed to examine the role of endogenous VN in osteoporosis using a comprehensive in vivo approach. Our study revealed that VN deficiency not only suppresses bone resorption but also increases osteoclastogenesis, providing intriguing insights into how VN regulates the pathogenesis of bone resorption in a mouse model of osteoporosis. This study highlights the potential clinical significance of inhibitors of VN activity and its potential effect on the bone remodeling process.
To induce osteoporosis in the femur, we performed an OVX procedure in mice. The OVX mice exhibited significant weight gain and uterine tissue atrophy compared to the sham-operated mice (Fig. 1). Micro-CT analysis confirmed a significant increase in the TV of the femurs in the OVX group, with a particular enlargement of the Ma.Ar, indicating an expanded medullary cavity (Fig. 5B). The expansion of the medullary cavity results from increased bone resorption in the lumen [15, 16]. Additionally, the ratio of Ct.Ar to the Tt.Ar decreased in the OVX group (Fig. 5C), indicating a tendency toward cortical bone loss and aligning with structural changes observed in the cortical bone [17]. These results confirmed the successful induction of osteoporosis in the femur by ovariectomy.
From the results of ALP and TRAP activity in the plasma of OVX- and sham-operated mice (Fig. 3), we observed that VN deficiency did not affect the activity of osteoblast but rather promoted the activity of osteoclasts. Consistent with this result, histochemical analysis of ALP- and TRAP-stained femurs indicated that VN deficiency did not affect the ratio of Ob.Pm to B.Pm but increased the N.Oc in OVX-operated mice (Fig. 4). Pathological bone diseases, such as osteoporosis, are characterized by an imbalance caused by increased activity and the N.Oc, leading to bone destruction [18]. TRAP, an enzyme expressed in osteoclasts, is released on the basolateral surface of cells [19] and is used as a histochemical marker for osteoclasts. We observed that the activity and N.Oc in the femur were increased by VN deficiency (Figs. 3, 4). A possible mechanism by which VN suppresses osteoclastogenesis in OVX mice is via the inhibition of macrophage colony-stimulating factor (M-CSF) signaling. Previous studies have reported that signals from M-CSF and RANKL are required for osteoclastogenesis [20, 21]. The association between these signals and VN was recently reported. A VN-derived peptide, VnP-8 (residues 206–217 in human VN), has been shown to impede M-CSF signaling through the degradation of its receptor c-Fms [22]. Furthermore, VnP-8 reduces the expression of RANK, a receptor for RANKL. These findings support the notion that VN may suppress the signaling pathways of RANKL and M-CSF and thereby suppress osteoclastogenesis (Fig. 6A). In addition, a VN-derived peptide (VnP-16) (residues 270–281 in human VN) has been shown to suppress osteoclastogenesis by initially inhibiting the RANKL–RANK–TRAF6 pathway and subsequently downregulating the JNK–c-Fos–NFATc1 signaling cascade, crucial for osteoclast maturation (Fig. 6A) [8, 23]. Moreover, it has been shown that VN reduces TRAP activity and pre-osteoclast number [24], consistent with our results. Despite VN deficiency leading to increased osteoclast activity, our results showed no significant effect on osteoblast activity (Figs. 3, 4). This distinct effect of VN on osteoblasts could be explained by the difference in the functional regions of VN between osteoblasts and osteoclasts. Previous studies have reported that a VN-derived peptide, VnP-8 (residues 206–217 in human VN), inhibits the pathway of RANKL–RANK–TRAF6s in osteoclasts and suppresses osteoclastogenesis, but does not show reproducible attachment to osteoblasts [22]. The effects of distinct regions of VN on osteoblasts and osteoclasts likely reflect its differential roles in modulating the activities of osteoclasts and osteoblasts through distinct regions of VN.
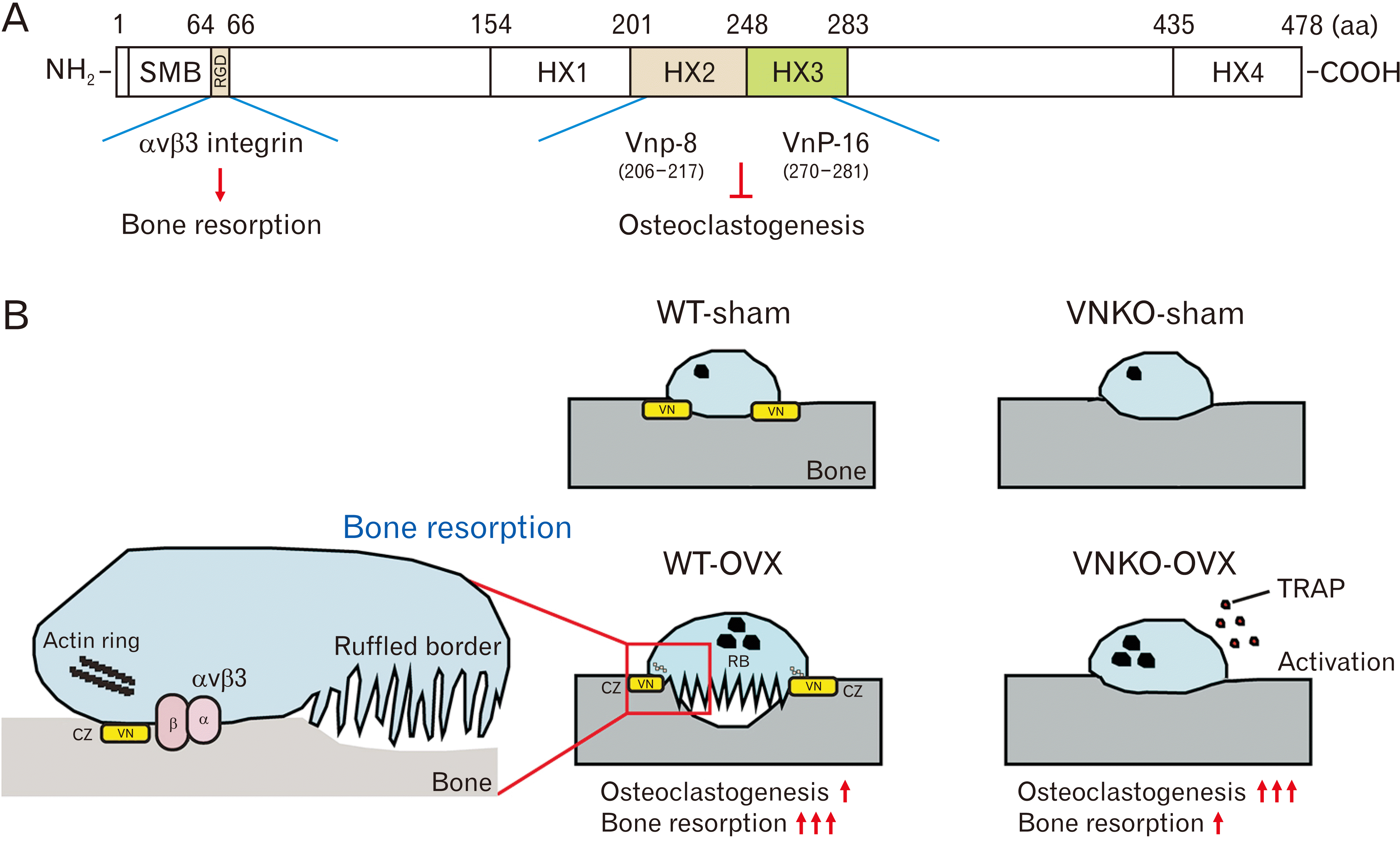 | Fig. 6Functional regions of vitronectin (VN) and the roles in osteoclastogenesis and bone resorption. (A) The amino acid (aa) positions and the domain structure of full-length VN are indicated. The schematic delineates specific regions of VN peptides (VnP-8: residues 206–217 and VnP-16: residues 270–281 in human VN), which are known to inhibit osteoclastogenesis. In contrast, the RGD motif (residues 64–66) is suggested to facilitate the attachment of osteoclasts to VN via αvβ3 integrin on the bone matrix, potentially augmenting bone resorption (B) Diagrams showing the role of VN in osteoclastogenesis and bone resorption in WT-OVX or VNKO-OVX mice. VN regulates osteoclastogenesis and the adhesion of osteoclast to bone surface. In VNKO-OVX mice, VN deficiency increased the activity of tartrate-resistant acid phosphatase (TRAP) and osteoclastogenesis. On the other hand, VN also suppresses osteoclastogenesis. However, VN deficiency suppressed bone resorption in VNKO-OVX mice. VN may regulate the attachment of osteoclasts to bone surface via αvβ3 integrins and induce the actin-rich clear zone (CZ) formation. Furthermore, VN may induce the formation of ruffled borders (RB), leading to bone resorption in WT-OVX mice. WT, wild-type; OVX, ovariectomized; WT-OVX, OVX-operated WT group; VNKO-OVX, OVX-operated vitronectin knockout group; WT-sham, sham-operated WT group; VNKO-sham, sham-operated VNKO group; RGD, Arg-Gly-Asp.
|
Regarding the effect of VN deficiency on bone resorption, our results showed that VN deficiency significantly suppressed the OVX-induced up-regulation of the medullary cavity, as evidenced by our micro-CT analysis of the parameters TV and Ma.Ar (Fig. 5B). Furthermore, we found that VN deficiency suppressed the OVX-induced upregulation of DA (surface roughness of the bone) (Fig. 5B). The surface roughness of bone has been reported to be important in initiating bone resorption by osteoclasts [25], and DA has been positively correlated with osteoporosis [26]. Our data supported the hypothesis that VN enhances bone resorption. Additionally, a radical change in trabecular bone from plate-like to rod-like has been observed in osteoporosis [27]. In WT-OVX mice, the SMI showed an increased tendency from 0, which represents a typical plate, to 3, which is typical for rods; this OVX-induced upregulation of the SMI value was suppressed by VN deficiency (Fig. 5B). These results suggested that VN increased bone resorption in the femur. To discuss the role of VN in bone resorption behavior, we can explain the process of bone resorption as follows: First, osteoclasts adhere to VN on the mineralized surface of bone through αvβ3 integrin (Fig. 6) [28]. This adhesion induces c-Src-dependent tyrosine phosphorylation of Pyk2 and p130Cas, leading to the formation of a large molecular complex. This complex activates downstream signaling molecules to induce cytoskeletal reorganization [29]. The latter enables the formation of an actin-rich clear zone (actin ring) in osteoclasts (Fig. 6B) [30-32]. After the actin ring is formed, a ruffled border is formed in the osteoclasts, and bone resorption begins (Fig. 6B) [33]. Previous studies had revealed that VN is mediated with the binding between osteoclasts and the bone surface through αvβ3 integrin receptor, known as the VN receptor and expressed in osteoclasts [34, 35]. These findings support the notion that VN plays a critical role in osteoclast adhesion to the bone surface. Additionally, VN-coated coverslips have been reported to induce osteoclasts not only to form ruffled borders but also to secrete hydrolytic enzymes and digest extracellular substrates [28]. Osteoclasts adhere and spread when coated with VN, and VN has been described as inducing bone resorption [11, 28]. In other words, VN can trigger bone resorption. Although VN deficiency increased osteoclastogenesis, it critically suppressed the adhesion of osteoclasts to the bone surface, thereby inhibiting the formation of actin rings and ruffled borders (Fig. 6B). This is a possible mechanism by which VN deficiency increases osteoclastogenesis, but suppresses bone resorption in OVX-operated mice (Fig. 6), as observed in this study.
In this study, we showed for the first time that VN suppresses osteoclastogenesis and promotes bone resorption in OVX-operated mice, serving as an osteoporosis model. These distinct functions of VN could be attributed to several functional regions in VN (Fig. 6A). Our study revealed that the Arg-Gly-Asp (RGD) motif could be a new target for anti-resorptive agent in OVX-operated mice. Besides VnP-8 and VnP-16, it is suggested that the RGD motif facilitates bone resorption via αvβ3 integrin and could be a new clinical target for osteoporosis treatment. This discovery highlights the potential of VN in treating osteoporosis and related bone disorders characterized by excessive osteoclast activity.
Go to :
 XML Download
XML Download