

 PDF
PDF Citation
Citation Print
Print



Introduction
Melatonin, a hormone produced by the pineal gland, is secreted in a rhythmic manner that follows the body’s circadian and seasonal cycles. This hormone is derived from serotonin and is not only secreted by the pineal gland but also by other parts of the body, including the testicles, ovaries, and peripheral nerves. The biosynthetic pathways for melatonin production appear to be consistent across these different locations. Melatonin performs a pleiotropic function and is involved in various physiological processes [1-5]. Melatonin has a significant impact on the immune system by regulating it. It also safeguards fatty acids from oxidation and has anti-apoptotic and anti-aging effects on cells through cell signaling and response. Melatonin is known to reduce the amount of reactive oxygen species (ROS) production in cells, thereby preventing DNA mutation caused by oxidative stress [6-8]. Because melatonin has both hydrophilic and lipophilic properties, it can aid in safeguarding mammalian gametes and embryos while they are being cultured. This is achieved by mitigating the harm caused by oxidative stress [9, 10]. Melatonin carries out its physiological function through G-protein signal transduction pathways. Researchers have identified three subgroups of melatonin based on the methodology used in vertebrates. MT1 and MT2 melatonin receptors are G-protein coupled receptors. They are expressed in mouse, rat, and human testis cells that contain germ cells in the testis, the tail and head of spermatids, and the epithelial cells of the epididymis, as well as in Leydig and Sertoli cells. This indicates the wide activity of this hormone produced by the pineal gland [2, 11, 12]. In many articles, melatonin has been added as a supplement to the spermatogonial stem cells (SSCs) culture system, and the role of this factor in proliferation and differentiation has been investigated [6, 13, 14]. In this article, we aim to compile articles that employ melatonin as a supplement in laboratory experiments, animal testing, or freezing solutions and explore the signaling role of this substance as an antioxidant and growth enhancer. The results of the articles show that melatonin has several mechanisms on Sertoli cells and Leydig cells or SSCs to promote the proliferation of spermatogonial stem cells and prevent the apoptosis of these cells and has a protective role that these mechanisms are described below and is presented in Tables 1-4.
Review
The presence of melatonin as a supplement in SSCs medium culture
Recent research has shown that the presence of melatonin as a supplement in the culture medium has a significant effect on improving the culture and survival of SSC colonies. However, more studies are required to find the signaling pathway. Fig. 1 shows a set of the most important signaling pathways of melatonin as a supplement in the culture medium. Table 1 also mentions the growth factors (GFs) that play a role in the proliferation of SSCs along with melatonin as a supplement, as well as endpoints, sources, and cell types.
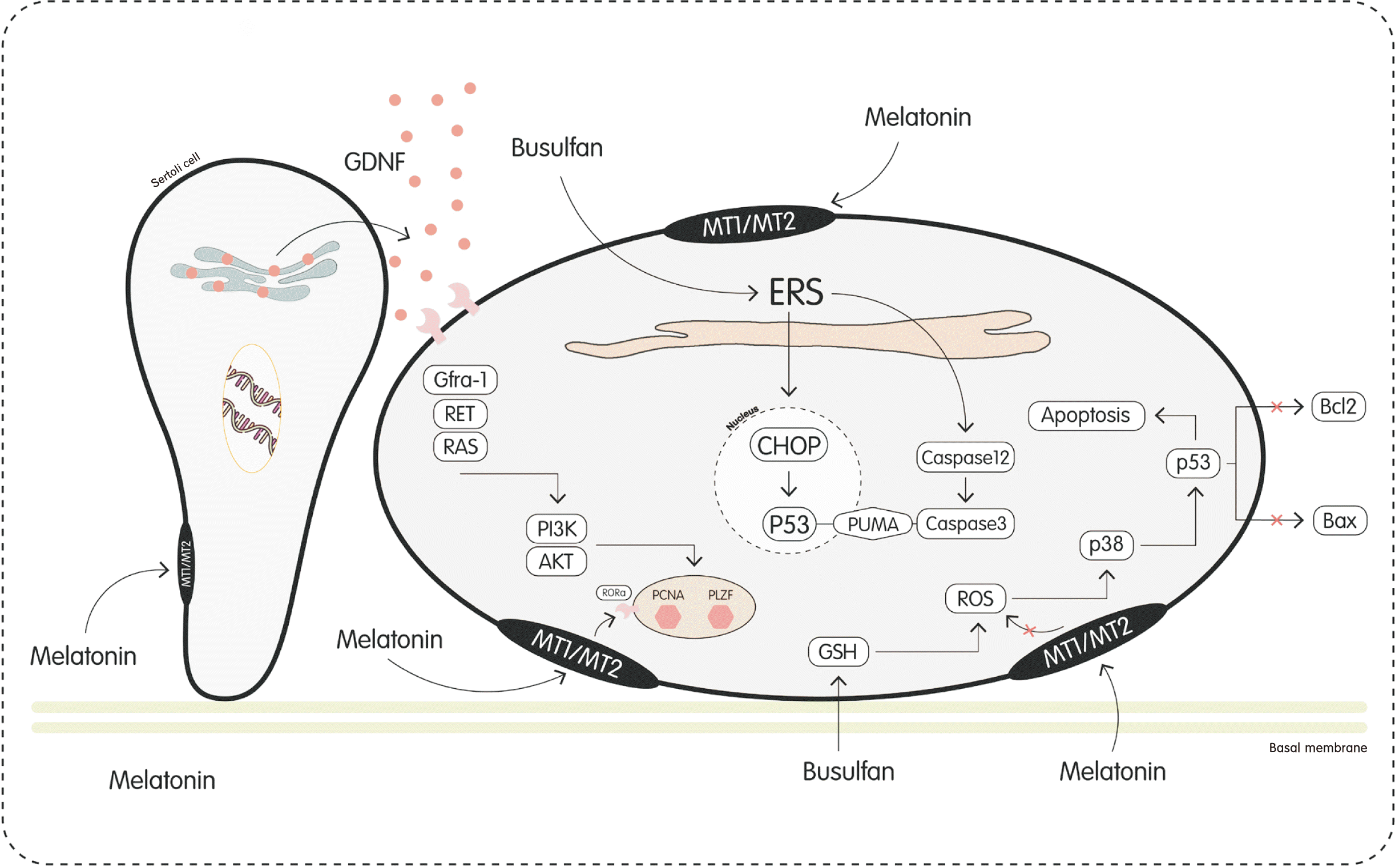
In 2016, Niu et al. [15] introduced melatonin for the first time in goat SSCs culture medium. They demonstrated that the presence of melatonin in the amount of 1 μM and 1 nM, respectively, improves the culture of SSCs and increases the proliferation of these cells. The results of their research indicate that the addition of melatonin to the culture medium reduces the level of mRNA expression of differentiating genes and stimulates Sertoli cells to produce glial cell line-derived neurotrophic factor (GDNF). GDNF activates the GDNF-GDNF family receptor α 1 (GFRα1)-RET-phosphatidylinositol 3-kinase (PI3K)-AKT pathway, which increases the proliferation of SSC (Fig. 1). After separation, SSCs were cultured in Dulbecco’s modified Eagle medium/nutrient mixture F-12 (DMEM/F12; Gibco) using GF. These factors include fetal bovine serum (FBS), knockout serum replacement (KSR) medium, GDNF, epidermal growth factor (EGF), basic fibroblast growth factor (bFGF), and GFRα1 [15].
In 2016, Deng et al. [16] investigated the effect of melatonin (10-7 M) on sheep’s SSCs. Their findings show that melatonin initially suppresses the level of cAMP, but in the next stage, it promotes its production level, as a result, testosterone production increases. The level of meiotic anaphase marker genes Dnmt3a and Bcl2 increases. After that, it leads to the differentiation of spermatogonia and finally the creation of sperm. The sperm is fertilized with oocytes MІІ in vitro, which developed successfully to the blastocyst stage. Testicular cells containing SSCs, Sertoli cells, were cultured in DMEM culture medium with GFs containing FBS, stem cell factor (SCF), bFGF, EGF, GDNF [16].
In 2017, Navid et al. [6] conducted a study on the culture and colonization of SSCs. They found that the use of 100 μM melatonin as a supplement in the culture medium can have a significant effect on improving the culture and colonization of SSCs. In addition to melatonin as an antioxidant, they also used FBS, GDNF, and human recombinant leukemia inhibitory factor (LIF) as GFs. The researchers stated that the presence of LIF in the medium as an inhibitor prevents the differentiation of SSCs. Therefore, they presented a new protocol for the culture of SSCs, which can be useful for the treatment of infertility caused by pre-puberty cancer diseases. This protocol greatly helps SSC banking [6].
In 2017, Reda et al. [17] cultivated tested testicular tissue pieces of neonate mice (3 days old) with different supplements such as melatonin and Glutamax, and KSR. The results showed that the melatonin supplement along with Glutamax has a positive effect on the maturation of male germ cells. They also concluded that the concentration of at least 10% KSR plays a decisive role in the maturation of germ cells and testosterone production [17].
In 2018, Zhang et al. [18] studied the effect of melatonin on neonatal mice that were under the influence of bisphenol A or di (2‐ethylhexyl) phthalate. These substances disrupt the self-renewal and the normal process of spermatogenesis and meiosis of SSCs. This disorder is due to the reduction of G9-dependent methylation. Melatonin maintains demethylation in these cells by maintaining histone 3 lysine 9 trimethylation (H3K9me1). Therefore, the normal process of spermatogenesis and meiosis is preserved. In this research, testes were cultured in agarose gel. The culture medium is DMEM/F-12 and FBS GF was used for the growth and cultivation of cells [18].
Deng et al. [19] investigated the effect of melatonin on sheep Leydig and Sertoli cells in this vitro. Melatonin reduces the expression of SCF and insulin-like growth factor-1 and estrogen synthesis in Sertoli cells. The increase in the expression of SCF is through the retinoic acid receptor-related orphan receptor-α (RORα), which is dependent on the MEK/extracellular signal-regulated kinase pathway. By increasing the expression of insulin-like growth factor-1 and decreasing the expression of estrogen in Sertoli cells, the expression of testosterone in Leydig cells increases [19].
Palmitic acid (PA) is one of the saturated fats in the diet, which leads to obesity-related infertility in male. In 2020, Xu et al. [20] investigated the effect of melatonin on mice that were drowned under the influence of PA. The research results showed that melatonin stimulated the expression and nuclear translocation of SIRT1 through its receptors and prevented the production of ROS caused by PA and mitochondrial dysfunction through the sirtuin type 1 (SIRT1) signaling pathway. Therefore, by activating SIRT1, melatonin reduces oxidative stress, endoplasmic reticulum stress (ERS), mitochondrial dysfunction, and DNA damage in SSCs [20].
Lv et al. [21] investigated the effect of melatonin, as an antioxidant, on mice SSCs that were affected by chromium (Cr[VI]) (toxin and mutagenic substance, especially in this vivo), and the research results show that Cr increases the ROS and decreases the expression of fusion proteins in the mitochondrial membrane such as MFN1, MFN2, and OPA1, and in the reverse direction, it increases the protein fused in the mitochondrial membrane called DRP1. As a result, disorders are created in the mitochondrial membrane. Mitochondria fuse with lysosomes and undergo autophagy. The presence of melatonin in the culture medium leads to a decrease in ROS production. In addition, with the production of methyltransferase-like 3 (METTL3)-mediated RNA m6A, it leads to an increase in the fused proteins in the mitochondrial membrane, Mfn2 and Opa1, and it also causes a decrease in the expression of the autophagy genes Bnip3 and Nix, which in total disrupts and damages the mitochondrial membrane and autophagy of this organelle, which ultimately leads to the stop of apoptosis of SSCs [21]. In line with this research, in 2017, the same researcher investigated the effect of melatonin on global histone methylation. H3K27me3 epigenetic activated the apoptotic signaling pathway in SSCs. The results showed melatonin alleviated Cr(VI)-increased ESET expression and Histone H3 lysine 27 trimethylation (H3K27me3) level. External administration of melatonin protected mice against Cr(VI)-induced changes in testicular tissue and apoptosis of SSCs, which helped to maintain the regular spermatogenesis process [22].
The presence of melatonin as a supplement in SSCs three-dimensional culture system
Navid et al. [13] cultured mouse spermatogonial cells in a three-dimensional (3D) soft agar culture model. The 3D soft agar culture system (SACS) consists of two layers. It contains an upper layer with 0.37% (w/v) agar and a lower solid layer with 0.5% (w/v) agar. The cell suspension for culture is added to the top layer. In the culture medium, they added melatonin as a supplement. The culture medium containing GFs FBS, GDNF, LIF. Their results show that the presence of melatonin along with LIF and SACS creates a testicular-like nutrient microenvironment that improves the culture of SSCs. The presence of melatonin in the 3D culture system (SACS) plays an important role in reducing ROS (Table 2) [13].
The presence of melatonin as a supplement in SSCs freezing medium
In this section of the collection of essays, we examine the effect of melatonin as an additive in the cryoprotectant solution. Subsequently, the cells are cultivated in the culture medium under the influence of GFs. The ability of SSCs is assessed using various techniques or by transplanting them into azoospermic adult mice. The GFs and cell origins are specified in Table 3.
In 2014, Gholami et al. [24] investigated the effect of melatonin on SSCs in the freezing medium and showed that melatonin leads to cell proliferation and induces apoptosis in damaged cells [24]. In 2015, in a similar study, the same researcher added melatonin to the freezing medium of mouse spermatogonial cells, which contains glycerol, and obtained similar results [25]. In 2016, Saki et al. [26] investigated the effect of melatonin as a strong and natural antioxidant on the cryopreserved testicular cells in mouse. They used lactate dehydrogenase activity test to obtain cytotoxicity and flow cytometry to investigate apoptosis. The results showed that melatonin has the property of scavenging free radicals [26].
In 2020, Feng et al. [27] investigated the effect of melatonin in the cryopreservation of SSC. The results of the research showed that melatonin increases the secretion of enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-Px). As a result, it activates the immune system, so it has an antioxidant role. This extensive antioxidant activity prevents the swelling and vacuolation of the mitochondrial membrane, thus preventing the release of cytochrome c from the mitochondria into the cytoplasm, thus preventing the release of caspase-3. As a result of which apoptosis of SSCs is inhibited. By regulating proteins related to autophagy, including light chain 3 (LC3)-I, LC3-II, P62, Beclin1, and ATG7, it prevents the formation of autophagosomes and autophagy of SSCs. It also decreases the expression of Bax gene and in a negative correlation increases the expression level of BCL2, so it shows another aspect of melatonin’s anti-apoptotic function [27].
Kazemzadeh et al. [28] added 100 μM melatonin to the basic medium of the freezing medium for long-term preservation of SSCs, then cultured SSCs obtained from the freezing medium in α-mem medium containing FBS, GDNF, bFGF, LIF GFs. Then, the proliferative quality of SSCs before transplantation was checked by the real-time technique of the expression level of PLZF and GFRα genes. The expression level of these genes was much higher in the medium containing melatonin. Then, they checked the quality of transplanting these cells into adult male azoospermic mice. The results showed that the presence of melatonin in the frozen base medium has an effective role in improving the quality of transplanting SSCs into adult male into mature azoospermic mice [28]. The same author conducted a similar research in 2021 and added melatonin to the basic freezing medium containing SSCs and checked the quality of these cells after transplanting into adult azoospermic male mice and reached a similar result [14].
The presence of melatonin as a therapeutic factor in vivo
In this section, we reviewed a collection of articles that prescribe melatonin as a therapeutic factor for animals. Subsequently, the animal is sacrificed, and the testicle tissue or testicular cells are examined using various techniques to assess the ability and quality of SSCs. The details of these articles are mentioned in Table 4. Additionally, Fig. 1 illustrates the most important signal paths in vivo.
Hemadi et al. [30] investigated the effect of melatonin on SSCs. SSCs were then injected into the testes of adult azoospermic mice. For 9 weeks after transplantation, melatonin (20 mg kg−1 per day) was administered to the treated group. The ultrastructural studies of the testis in the presence of melatonin showed that the degenerative and apoptotic changes in the basement membrane and epithelial cells of the spermatogenic tubules were significantly less than the untreated group [30].
In 2013, Abd El Aziz and Metwally [31] investigated the effect of melatonin on rats under the influence of busulfan. The results obtained from testicular tissue sections show that in the group without the presence of melatonin, the increase in the average number of caspase-3 positive cells in the seminiferous tubules is a sign of the decrease of germ cells. However, in the group treated with melatonin, in the tissue section of the testis, the almost complete regeneration of germ cells with the appearance of round and elongated spermatids in seminiferous tubes indicates the improvement and increase of germ cells [31].
Cui et al. [32] investigated the destructive effect and apoptosis of SSCs and testicles of mice under the influence of busulfan and found that the presence of melatonin (10 mg/kg/day) can reduce the apoptosis of SSCs and destructive effect of testicular tissue. Busulfan is used for chemotherapy. During the chemotherapy process, serious damage is done to SSCs and testicular tissue. To prevent infertility, melatonin has been used as a broad-spectrum antioxidant in this research. The process is such that busulfan causes the secretion of ERS. Busulfan is used for chemotherapy. During the chemotherapy process, serious damage is done to SSCs and testicular tissue. To prevent infertility, melatonin has been used as a broad-spectrum antioxidant in this research. The process is such that busulfan causes the secretion of ERS. The presence of melatonin as a therapeutic factor block the secretion of proteins caspase-3, caspase-12, and C/EBP homologous protein (CHOP) as well as causes a decrease p53 upregulated modulator of apoptosis (PUMA) and P53 proteins are secreted. In addition to, melatonin has other therapeutic effects, such as stimulating the secretion of gonadotropins and testosterone, which is useful in the treatment of infertility (Fig. 1).
In another study conducted by Deng et al. [33] in 2017, focus on melatonin receptors. According to Immunocytological techniques, MT1 and MT2 receptors (intracellular receptors) are located in the primordial germ cells and Leydig cells. Retinoic acid receptor-RORα is located in the perinuclear areas of Leydig cells and at lower levels of Sertoli cells and in primordial germ cells. This receptor (nuclear receptor) under the influence of melatonin increases the level of expression of GATA-4, then it increases the expression of SF1 and StAR, which as a result increases the concentration of testosterone in vivo. Therefore, RORα pathway plays a role in inhibiting the proliferation and differentiation of SSCs. This pathway is contrary to the MT1 and MT2 pathway, which leads to the proliferation of SSCs [33].
Similar to the previous research, Li et al. [34] investigated the effect of melatonin on apoptosis SSCs and testicular damage in vivo. In this research, melatonin and busulfan were injected intraperitoneally in different doses to male mice. Then SSCs obtained from the testes of mice were evaluated in this vitro. The results show that melatonin increased the expression of MnSOD and SIRT1, which decrease ROS and p53, respectively (Fig. 1) [34].
Du et al. [34] showed in a research on diabetic mouse model that high glucose leads to induction of apoptosis on Leydig cells, which leads to testicular damage and ERS reactions. The presence of melatonin in the treatment group, it protects the Leydig cells. Therefore, the induction of apoptosis is inhibited by high glucose concentration. At the same time, the secretion of CSF-1 by Leydig cells induces the self-renewal process of SSCs. Therefore, the presence of melatonin in the diabetic model leads to the improvement of infertility [35].
In 2019, Zhang et al. [36] investigated the effect of melatonin on the effects of chemotherapy. They continuously injected melatonin to the mice that were under the influence of busulfan. The results of the tests showed that melatonin destroys the ROS due to the production of Sod, Gpx, and Cat. Therefore, it brings the production of P-ATM and P53 to the normal level and stops cell arrest and apoptosis in SSCs [36].
In 2021, Yang et al. [37] investigated the effect of melatonin in mice exposed to lipopolysaccharide (LPS) associated with ERS. LPS leads to testicular damage, apoptosis, ERS, and inflammatory response in SSCs. Toll-like receptor 4 and inflammatory responses are increased by LPS treatment. Melatonin significantly affects the expression of apoptotic genes related to ERS, has a reparative and anti-infection role in testicular damage caused by LPS [37].
In 2022, Li et al. [38] investigated the effect of melatonin on spermatogonial cells in contrast to hexavalent chromium (Cr[VI]). C57BL/6 J male mice were injected intraperitoneally for 14 days in the control group with only Cr(VI) and in the experimental group with melatonin and Cr(VI) and they were sacrificed on day 21. Their results showed that exposure to Cr(VI) can cause DNA damage, G2/M arrest and apoptosis in SSCs. Inspiringly, melatonin protected SSCs from Cr(VI)-induced damage by scavenging ROS, by suppressing ATM-p53 phosphorylation and the MAPK pathway, as well as by restoring H3K9me3 levels in cell cycle promoter and apoptosis-related regions [38]. Based on the research conducted, melatonin, as a hormone and antioxidant factor, can effectively improve the colonization, anti-apoptotic, and even differentiation of SSCs. All the mechanisms obtained from the conducted research confirm these results.
Conclusion
The signaling pathway of melatonin is composed of complex molecular mechanisms. However, the present study aimed to examine the signaling pathways of melatonin in various culture media, including 2D and 3D, as well as frozen conditions. It also explored the potential of melatonin as a supplement and therapeutic agent. Considering the significance of melatonin as a hormone and antioxidant, further research is required to uncover the mechanisms of action of melatonin on testicular cells, such as SSCs, Sertoli, and Leydig cells.
 XML Download
XML Download