

 PDF
PDF Citation
Citation Print
Print



Highlights
• GLP-1 increases ABCA1 mRNA and apo AI, mediating intracellular cholesterol efflux.
• GLP-1 induces FNDC5 expression in β-cells via CREB, promoting WAT to BAT conversion.
• GLP-1 oxidates fatty acids and reduces its influx into hepatocytes, inhibiting NAFLD.
• GLP-1R/GIPR dual agonists improve lipid metabolism, with greater potential than GLP-1RAs.
Go to : 
The DEVELOPMENT AND USE OF GLUCAGON-LIKE PEPTIDE-1
Glucagon-like peptide-1 (GLP-1) is a 30-amino acid peptide hormone produced by intestinal L cells through differential processing of the proglucagon (PG) gene [1]. Kimball and Murlin [2] discovered PG in 1923. GLP-1 was discovered in 1983 when Bell et al. [3] analyzed the genetic sequence of glucagon. PG gene sequence consists of six exons and five introns, and GLP-1 (72–108) is one of the main domains. PG gene is expressed in pancreatic α-cells, intestinal L cells, and some nerve cells, and through tissue-specific translation modification, different end products are finally formed. While GLP-1 is mainly expressed and translated in intestinal L cells. In 1987, GLP-1 (7–36) was recognized as a physiological incretin in the studies of Kreymann et al. [4]. GLP-1 is derived from the PG precursor, which is a class of highly efficient incretins released by the body in response to nutrient intake. Such as the release of GLP-1 can be directly stimulated by nutrients such as glucose and fat in the intestinal lumen [5]. In 2005, exenatide was approved as a kind of GLP-1 receptor agonists (GLP-1RAs) for the clinical treatment of type 2 diabetes mellitus (T2DM) as the first international incretin drug [6]. Following the application of exenatide, other GLP-1 receptor (GLP-1R) activators were developed such as liraglutide (Lira). Meanwhile, the application of GLP-1RAs in non-diabetic diseases has also been launched.
Go to :
STRUCTURE, DISTRIBUTION, AND PHYSIOLOGICAL FUNCTION OF GLP-1
GLP-1 is composed of 30 amino acid residues, which primary structure begins with histidine and ends with glycine. There are a variety of molecular forms in the body which main active forms are GLP-1 (7–36) NH2 and GLP-1 (7–37), of which about 80% exist in the form of GLP-1 (7–36) NH2, which is first decomposed and inactivated in the blood circulation. Then it is eliminated by liver metabolism and renal excretion [7]. GLP-1 has a variety of effects, originating from the presence of its receptors in specific nuclei of the lung, pancreas, gastrointestinal tract, kidney, heart, bone, central nervous system, etc. In recent years, studies have found that the expression of GLP-1R is also found in liver and adipose tissue (Fig. 1) [8]. In the lung, GLP-1 binding to its receptors improves lung function and reduces mortality in patients with obstructive pulmonary disease [9]. In the pancreas, GLP-1 binding to its receptor stimulates the proliferation and regeneration of islet β-cells and inhibits their apoptosis [10]. In the gastrointestinal tract, GLP-1 binding to its receptors delays gastric emptying, reduce appetite, and increase satiety [11]. In the kidney, GLP-1 binding to its receptor has extensive renal protective properties. This characteristic may improve renal function by increasing renal blood flow, urine flow rate, preventing elevated plasma creatinine, reducing tubular necrosis, increasing renal interstitial fluid and glomerular filtration rate, and cytoprotective and anti-inflammatory effects [12,13], and may improve glomerular hyperosmosis in patients with T2DM in some cases [14]. In bone, GLP-1 binding to its receptor promotes bone marrow mesenchymal stem cell osteoblast differentiation and inhibits its differentiation into fat cells, thereby promoting bone formation and improving osteoporosis [15]. In the central nervous system, the action of GLP-1 and GLP-1RAs on its receptors can affect a variety of brain functions including satiety, thermogenesis, influence blood pressure, neurogenesis, neurodegeneration, retinal repair, and change energy homeostasis [16-20]. And its neuroprotective effect has become a research hotspot in recent years. In the heart, GLP-1 binding to its receptor enhances myocardial performance and has important physiological effects on cardiac function through direct receptor-mediated [21-24]. Several studies have shown that the clear link between glucose metabolism and GLP-1 through the protein kinase A (PKA) pathway [25-27]. In addition, GLP-1 has now been proved to be associated with nutrient intake and can be linked to lipid metabolism [28-30]. This article will review the role of GLP-1 in lipid metabolism in detail.
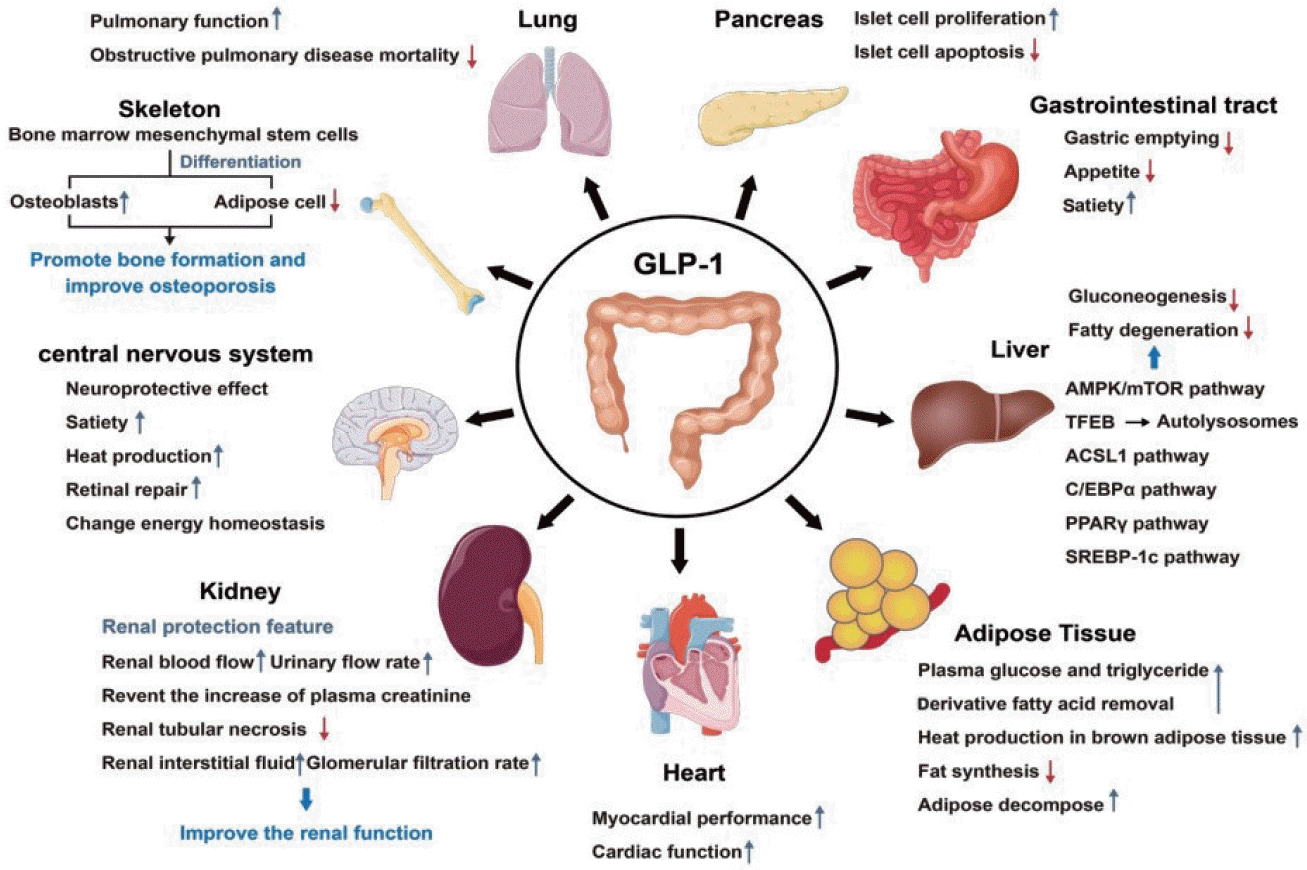 | Fig. 1.Physiological function of glucagon-like peptide-1 (GLP-1). AMPK, AMP-activated protein kinase; mTOR, mammalian target of rapamycin; TFEB, transcription factor EB; ACSL1, acyl-CoA synthetase long chain family member 1; C/EBPα, CCAAT/enhancer binding protein α; PPARγ, peroxisome proliferator-activated receptor-γ; SREBP-1c, sterol regulatory element binding protein-1c.
|
In liver tissue, GLP-1 levels are considered to be highest in human organs [31]. The induction of GLP-1 or its analogs in the liver can regulate varieties of processes, including hepatic gluconeogenesis, glycogen synthesis, and glycolysis [32]. However, the expression of GLP-1R in liver tissue remains controversial. To date, studies in animals and humans suggest that GLP-1 may improve hepatic lipid accumulation through signal transduction of the AMP-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) pathway [33]. Downregulating the expression of miR-23a and increasing the expression of the mitochondrial protective gene peroxisome proliferators-activated receptor-γ coactivator-1 (PGC-1α) can exert an anti-apoptotic effect on human hepatocytes [34]. Meanwhile, GLP-1R has been shown to exist in human liver cells. GLP-1 analogs (exendin-4) can also improve hepatic steatosis [35] in ob/ob mice both in vivo and in vitro [36]. Lira (GLP-1R) can reduce the occurrence of hepatic steatosis in vivo and in vitro by enhancing autophagy and lipid degradation through the GLP-1R-transcription factor EB (TFEB)-mediated autophagy lysosomal pathway [37]. These evidences all suggest that the action of GLP-1R in liver cells may be directly stimulated by ligand-receptor action. However, some scholars have proposed that the effect of GLP-1 in the liver is indirectly mediated by multiple mechanisms, such as reducing fat absorption or inhibiting fat cells through acyl-CoA synthetase long chain family member 1 (ACSL1), CCAAT/enhancer binding protein α (C/EBPα), peroxisome proliferator-activated receptor-γ (PPARγ), and sterol regulatory element binding protein-1c (SREBP-1c) pathways to achieve the purpose of reducing liver lipids [38]. Alternatively, hepatic steatosis can be alleviated by regulating insulin clearance in the liver [39]. Although the expression of GLP-1R in liver tissue is controversial, GLP-1RAs have been shown to act as regulatory pathways affecting metabolic disorders.
GLP-1 has a direct effect on adipose tissue. GLP-1 may affect systemic energy metabolism by regulating adipocyte development, accelerating plasma clearance of glucose and triacylglycerol-derived fatty acids, improving insulin signaling, and stimulating thermogenesis in brown adipose tissue (BAT) [40-45]. For example, GLP-1R in adipocytes can directly regulate apoptosis and preadipocyte proliferation by activating the adenylate cyclase/cyclic adenosine monophosphate (AC/cAMP) signaling pathway and various cell signaling cascades, including extracellular signal-regulated kinase (ERK), protein kinase C (PKC), and protein kinase B (AKT) [41,46]. GLP-1 also regulates lipid metabolism in BAT and white adipose tissue (WAT) through the brain-adipocyte axis [43,44]. In differentiated human adipocytes in vitro, GLP-1 can regulate the expression of adipogenesis and lipolytic genes [47]. The direct effect of GLP-1 on adipose tissue has been demonstrated in many studies. However, the specific receptors that mediate and participate in it have not been determined, which are needed to be verified by establishing multiple experimental models.
Go to :
GLP-1 MODULATES LIPID METABOLISM
GLP-1 modulates lipid metabolism in the liver
GLP-1 Modulates fat synthesis in the liver
The liver is the main site of fat synthesis. Hepatic fat synthesis is a process by which carbohydrates and amino acids are converted into fatty acids and condensed with glycerol to form triglycerides (TG) [48]. Hepatic fat transport is the transport of TG out of the liver in the form of very low-density lipoprotein (VLDL) to participate in the oxidative breakdown of tissues or storage in adipose tissue [49]. The synthesis and transport of liver fat is the key to maintaining the homeostasis of TG in the liver. And its anabolic-related genes include fibroblast growth factor-21 (FGF21), SREBP-1c, fatty acid synthase (FAS), acetyl-CoA carboxylase (ACC-a), liver X receptor (LXR), suppressor of cytokine signaling-3 (SOCS-3), etc.
A recent study pointed out that GLP-1 can reduce liver fat synthesis. After giving long-term reared ovariectomy (OVX) mice with different gradient protein concentrations (standard, low protein, high protein) diet, it was concluded that a large number of lipid droplets were visible in the model group of hepatocytes hematoxylin and eosin (HE) staining and oil red O staining compared with the control group mice. In contrast, the expression of GLP-1 mRNA in the colon tissues of mice in the model group decreased, and the content of GLP-1 in serum decreased. Interestingly, FGF21, SREBP-1c, and FAS mRNA in the liver tissues of the model group of mice and protein expression was increased compared with the control group, while liver kinase B1 (LKB1) mRNA expression was reduced. The above conclusions all proved that a long-term high protein diet could reverse the above changes in OVX mice, while there was no significant difference between the low protein group and the model group. In this experiment, the high protein diet can promote intestinal secretion of GLP-1, improve FGF21 resistance, upregulate LKB1 to activate the AMPK pathway, downregulate the expression of SERBP-1c mRNA and protein in the liver, and ultimately inhibit liver lipid synthesis [50].
More interestingly, male Sprague-Dawley rats were randomly divided into the normal control (NC) group, high-fat (HF) group, and HF+Lira (GLP-1RA) group. Cell experiments showed that the arrangement of liver cells in rats in the HF group was disordered, and some hepatocytes were degenerated. There were more lipid droplets and inflammatory cell infiltration in hepatocyte cells. However, the fatty alteration of hepatocytes in the HF+Lira group was significantly improved. There were a small number of fat droplets in liver cells, and the infiltration of inflammatory cells was improved. Compared with the NC group, animal experiments found that the serum aspartate transaminase (AST), alanine transaminase (ALT), total cholesterol (TC), and TG were increased in the HF group, and the level of liver TG was also increased. Compared with the HF group, the HF+Lira group decreased serum AST, ALT, and TC, serum TG levels decreased significantly, the mRNA expression of SOCS-3 and SREBP-1c decreased significantly, and the liver TG level was also reduced. The above studies suggest that GLP-1 can downregulate the expression of SOCS-3 and SREBP-1c in rats with nonalcoholic fatty liver disease (NAFLD), reduce hepatic TG deposition. GLP-1 is also consistent with the conclusion that GLP-1 can inhibit liver lipid synthesis [51]. Boland et al. [52] gave diet-induced obesity (DIO) mice daily GLP-1R/Gcg receptor (GcgR) double agonist (cotadutide), Lira, Gcg agonist (g1437), and Lira+g1437 respectively. They found that all groups receiving GcgR agonists showed lower liver lipid content than the control group. In the hyperglycemic clamping experiment, cotadutide was also found to reduce the de novo fat production of the liver during clamping. It was concluded that cotadutide can inhibit the production of key fats in the liver of mice and reduce the lipid content of the liver. In animal studies by Burmeister et al. [53], GLP-1R knockout group in the hypothalamus was also activated and the liver TG increased in rats compared with the control group. Besides, KKAy mice were selected and divided into the Lira group and the HF feeding group. The results were shown that the serum TC and TG contents of mice in the model group and the Lira group were significantly increased compared with the control group. Interestingly, compared with the model group, the TC and TG contents of the Lira group had a decreasing trend but the difference was not statistically significant. The results of HE staining showed that compared with the control group, the model group had swollen hepatocytes and vacuolar changes (mainly steatosis). Compared with the model group, hepatocytes in the Lira group had less swelling, fewer vacuolar changes, more nuclei, and less inflammation. Compared with the model group, hepatocytes in the Lira group had less swelling, fewer vacuolar changes, more nuclei, and less inflammation. Phosphoinositide 3-kinase (PI3K) protein expression was significantly increased in the Lira group compared to the model group [54]. Similarly, animal experiments have shown that the expression of liver tissue FAS and ACC-a mRNA in rats in the Lira group was significantly higher than the control ratio [55]. At present, several studies have found that GLP-1 analog Lira may improve lipid metabolism and fat deposition by downregulating the expression of lipid metabolism-related genes FAS, and ACC-a mRNA, or promoting the expression of PI3K protein in mouse liver. The above results suggest that GLP-1 is important for inhibiting liver fat synthesis, and is likely to be related to AMPK, SERBP-1c, FAS, ACC-a, PI3K, and other signaling pathways.
In vitro studies, HepG2 cells treated with palmitic acid (PA) were found that dulaglutide significantly reduced liver lipid accumulation and gene expression related to lipid drop-binding proteins, de novo adipogenesis and TG synthesis. And dulaglutide increased the expression of proteins associated with lipolysis and fatty acid oxidation, as well as family with sequence similarity 3 member A (FAM3A) [56]. In the HepG2 fatty liver cell model, hypoxia-inducible factor-2alpha (HIF-2α)/PPARα pathway was found to be essential for Lira to inhibit liver lipid synthesis and reduce lipid-induced hepatic steatosis [57]. In addition, in the HepG2 cell experiment, some extrahepatic factors, such as erythroid-2-related factor 2 (NRF2) pathway [58], intestinal metabolite sodium butyrate (NaB) [59], Src homology region 2 domain-containing phosphatase-1 (SHP1)/AMPK [60], sirtuin 1 (SIRT1)/heat shock factor 1/heat shock protein (HSP) [61], and other signaling pathways have also been found to be involved in the regulation of liver lipid synthesis. Lira was found in PA-induced Kupffer cells to reduce HF diet-induced hepatic steatosis by inhibiting NOD-like receptor thermal protein domain associated protein 3 (NLRP3) inflammasome activation [62]. Gupta et al. [63] first reported the existence of G protein-coupled receptor (GPCR) GLP-1R in human hepatocytes in 2010, and provided evidence to prove the direct effects of GLP-1 protein on cells. GLP-1R binds to ligands to internalize GLP-1R, and can independently reduce TG accumulation in hepatocytes by activating key signaling molecules downstream of insulin receptor substrate 2 (IRS2). Since then, a number of studies have also been conducted in human primary hepatocytes, providing evidence for the direct effect of GLP-1 binding with GLP-1R in hepatocytes on reducing liver lipids [64]. In the culture system of various isolated hepatocytes (mouse primary hepatocytes, HepG2 human hepatocytes, human primary hepatocytes, etc.), Liu et al. [65] found that Lira’s stimulation of liver FGF21 expression may require the expression of GLP-1R in extrahepatic organs, and the increase of liver FGF21 is also necessary for Lira to improve hepatic lipid homeostasis. In cell experiments, GLP-1 was found to have more mechanisms that may inhibit liver fat synthesis, such as FAM3A, NRF2, NaB, SHP1/AMPK, NLRP3, FGF21, IRS2, and other signaling pathways, which also provided direct evidence for the role of GLP-1R in the regulation of lipid metabolism in the liver.
GLP-1 modulates hepatic lipolysis
Lipolysis is the process of converting TG into glycerol and fatty acids under the action of hydrolase, such as adipose triacylglyceride lipase (ATGL), hormone-sensitive lipase (HSL), and monacylglycerol lipase, in which fatty acids can provide energy for the body through β-oxidation or other oxidation pathways. And lipolysis is also affected by many lipid-associated proteins, such as SIRT1, α-subunit of PGC-1α, fatty acid-binding protein 4 (FABP4), etc. [66].
In a Lira-related study, healthy male C57BL/6 rats were evenly divided into three groups, namely NC group, model control group (OC group), and Lira group. Mice in the NC group were fed standard feed, while mice in the OC group and mice in the Lira group were fed HF feed. Compared with the NC group, the liver TG level, total liver TC level, and liver weight of the OC group were increased. And the mRNA expression levels of SIRT1, PGC-1α, and phosphoenolpyruvate carboxykinase (PEPCK) in the liver of the mice were significantly reduced. Compared with the OC group, the liver TG level, total liver TC level and liver weight of the mice in the Lira intervention group were significantly reduced. And the mRNA expression levels of SIRT1, PGC-1α, and PEPCK in the liver of the mice were significantly increased. This indicates that Lira can improve liver fatty acid oxidation in obese mice, reduce liver fat accumulation, and promote hepatic lipolysis by activating the liver SIRT1/PGC-1α/PEPCK pathway [67]. However, a small sample found that Lira did not significantly change levels of the hepatic fat oxidation marker β-hydroxybutyrate, although it reduced liver fat [68]. This conclusion should not be overlooked as a distraction factor with small sample size. Therefore, the effect of GLP-1 on liver lipolysis in clinical studies can be further developed.
GLP-1 modulates liver cholesterol metabolism
As the central organ of cholesterol metabolism, the liver directly affects the body’s cholesterol metabolism homeostasis. Hepatic cholesterol metabolism involves multiple parts, including cholesterol uptake, synthesis, biotransformation, and efflux. And it is related to cholesterol metabolism genes such as LXRα, ATP-binding cassette transporter A1 (ABCA1), ATP-binding cassette G1 (ABCG1), acyl-coenzyme A oxidase 1 (ACOX1), apolipoprotein AI (apo AI) (Fig. 2) [69]. Ben-Shlomo et al. [70] found in vivo and in vitro studies that GLP-1 and its analog exendin-4 were able to induce an increase in ABCA1 mRNA, increase the expression of apo AI mRNA and promoter, thereby increasing apo AI secretion, indicating that GLP-1 in hepatocytes may mediate the outflow of intracellular cholesterol to apo AI through the ABCA1 pathway. Recent studies have also demonstrated that exendin-4 is mediated by Ca2+/calmodulin (CaM)-dependent protein kinase kinase/CAM-dependent protein kinase IV/prolactin regulatory element binding (CaMKK/CaMKIV/PREB) pathway increases liver ABCA1 expression and reduces cholesterol accumulation [71]. Similarly, studies such as Ben-Shlomo et al. [70] have found that GLP-1 directly acts on LXRα in liver cells to mediate cholesterol outflow. These studies suggest that GLP-1 may mediate cholesterol outflow in hepatocytes through the LXRα-ABCA1 pathway, consequently avoiding large amounts of cholesterol accumulation in cells.
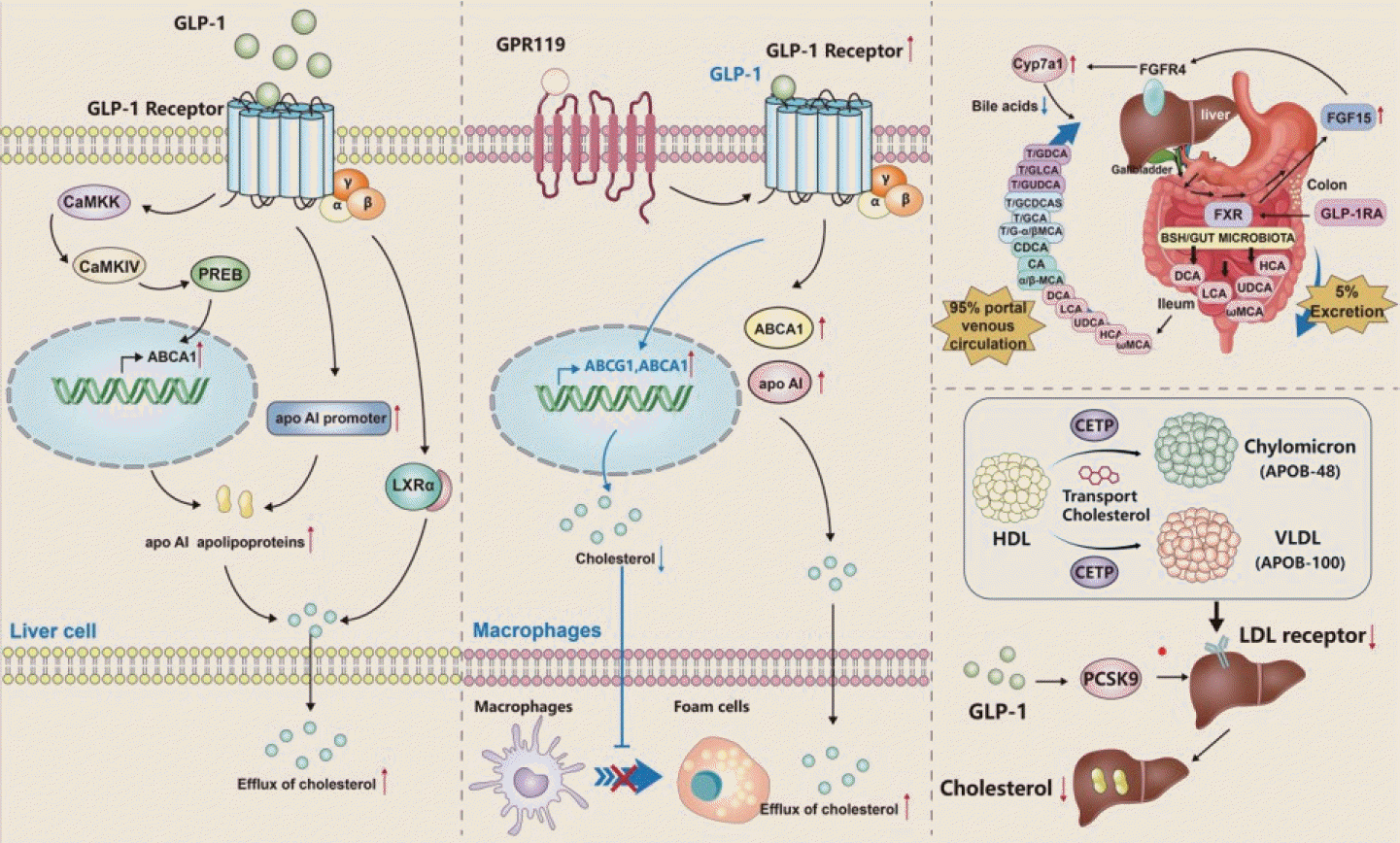 | Fig. 2.Possible mechanisms by which glucagon-like peptide-1 (GLP-1) modulates liver cholesterol metabolism. (Left) In liver cells, GLP-1 can mediate cholesterol efflux by acting directly on LXRα, or by inducing an increase in ATP-binding cassette transporter A1 (ABCA1) mRNA through Ca2+/calmodulin (CaM)-dependent protein kinase kinase/CAM-dependent protein kinase IV/Prolactin regulatory element binding (CaMKK/CaMKIV/PREB), increasing apolipoprotein AI (apo AI) mRNA and promoter expression, and increasing apo AI secretion, thereby mediating intracellular cholesterol efflux. (Center) In macrophages, GLP-1 can significantly increase the expression of GLP-1 receptor through G protein-coupled receptor 119 (GPR119), enhance the activity of ABCA1, lead to the increase of apo AI, and thus promote intracellular cholesterol outflow. The expression levels of ABCA1 and ATP-binding cassette G1 (ABCG1) can also be increased by GLP-1, which inhibits the transformation process of macrophages into foam cells and reduces the level of intracellular cholesterol. (Right) GLP-1 receptor agonist (GLP-1RA) reduces cholesterol accumulation by affecting the expression of genes in the hepato-enteral circulation and cholesterol reverse transport pathway, such as farnesoid X receptor (FXR), fibroblast growth factor-15 (FGF15), fibroblast growth factor receptor 4 (FGFR4), cytochrome P450 family 7 subfamily A member 1 (Cyp7a1), proprotein convertase subtilisin/kexin type 9 (PCSK9), low-density lipoprotein receptor (LDLR), etc. T/GDCA, T/glycodeoxycholic acid; T/GLCA, T/glucoronic acid; T/GUDCA, T/glycoursodexoycholic acid; T/ GCA, T/glycocholic acid; MCA, muricholic acid; CDCA, chenodeoxycholic acid; CA, cholic acid; DCA, deoxycholic acid; LCA, lithocholicacid; UDCA, ursodeoxycholic acid; HCA, hydroxy carboxylic acid; BSH, bile salt hydrolase; CETP, cholesteryl ester transfer protein; HDL, high density lipoprotein; VLDL, very low density lipoprotein.
|
Recent studies have pointed out that GLP-1 can increase the expression of ABCA1 and ABCG1 in ApoE-/- mice, thereby reducing the area of aortic plaque in HF raised ApoE-/- mice and realizing the reverse transport of aortic plaque. At the cellular level, GLP-1 can significantly increase the expression levels of ABCA1 and ABCG1, and decrease the level of intracellular cholesterol, which inhibits the transformation of macrophages into foam cells [72]. Studies have also shown that GLP-1 can significantly increase the expression of GLP-1R through G protein-coupled receptor 119 (GPR119), enhance the activity of ABCA1, lead to the increase of apo AI, and thus promote intracellular cholesterol outflow [73]. The GLP-1 analog Lira has been shown to have similar effects on cholesterol metabolism with GLP-1 [74].
Meanwhile, in the latest animal studies, some researchers found that exendin-4 can reduce levels of serum VLDL cholesterol and low-density lipoprotein cholesterol (LDL-C) by reducing liver sterol regulatory element binding protein 2 (SREBP2) level and cholesterol absorption [75]. The hepato-enteral circulation theory of bile acids is well known; that is, cholesterol is converted to bile acids in the liver, 95% bile acids are absorbed by the distal ileum and returned to the liver, and 5% bile acids are excreted in the stool. In order to maintain the bile acid pool, therefore, the amount of bile acid excreted is almost equal to the amount newly synthesized by the liver. Therefore, a possible mechanism proposed by the authors is that the activation of the farnesoid X receptor (FXR) by exendin-4 stimulates the expression of FGF15 in the small intestine and inhibits the expression of cytochrome P450 family 7 subfamily A member 1 (Cyp7a1) in the liver through fibroblast growth factor receptor 4 (FGFR4), thereby reducing fecal bile acid excretion and liver bile acid synthesis [75]. Interestingly, Lira has also been shown to attenuate negative feedback inhibition of bile acids by inhibiting intestinal FXR activity and ultimately reduce bile acid synthesis in the liver [76]. It has also been reported that cholesteryl ester transfer protein (CETP) can transport cholesterol from high-density lipoprotein to lipoprotein containing apolipoprotein B (ApoB) and rich in TG, such as chylomicron remains (containing ApoB-48) and VLDL remains (containing ApoB-100). These cholesterol can then be absorbed into the liver via low-density lipoprotein receptors (LDLRs) or LDLR-associated proteins [77]. In theory, if the LDLR is inhibited, the reverse transport of cholesterol is also inhibited, and less cholesterol is absorbed into the liver, thus reducing the accumulation of cholesterol in the liver. There are varieties of studies that confirm this conclusion. For example, Yang et al. [78] found that Lira inhibited proprotein convertase subtilisin/kexin type 9 (PCSK9) expression in HepG2 cells and db/db mice through hepatocyte nuclear factor 1 (HNF1)-dependent mechanism, and may inhibit LDLR expression in db/db mice through PCSK9 independent pathway, and ultimately reduce liver cholesterol accumulation. Animal studies have also shown that portal vein GLP-1 weakens intestinal fat absorption and chylomicron production through the vagal enteric-brain-liver axis, and reduces postprandish and fasting blood lipids [79].
These studies show that GLP-1 is closely related to cholesterol metabolism. GLP-1 can promote intracellular cholesterol outflow, reduce intracellular cholesterol content, and maintain intracellular cholesterol homeostasis through the ABCA1 pathway. While GLP-1 has not been clearly studied and reported through other ways to regulate intracellular cholesterol outflow. In addition, GLP-1 can reduce liver cholesterol accumulation by inhibiting SREBP2 and PCSK9, and reduce liver bile acid synthesis by weakening the negative feedback inhibition of bile acid after inhibiting FXR.
GLP-1 modulates fat lipid metabolism
GLP-1 modulates adiposynthetic and differentiation
Fat synthesis includes three aspects: de novo synthesis of saturated fatty acids, extension of fatty acid carbon chains, and synthesis of unsaturated fatty acids. The differentiation of adipose precursor cells consists of two stages, the former stage is the differentiation of adipose precursor cells into immature adipocytes, and the latter stage is the transformation of immature adipocytes to mature adipocytes, which associated genes include PPARγ, C/EBPα, adipocyte fatty acid-binding protein (AP2), lipoprotein lipase (LPL), etc.
After treating 3T3-L1 cells with GLP-1, some scholars found that the number of small and medium-sized adipocytes increased. And oil red O staining showed that GLP-1 had no significant effect on lipid accumulation. Compared with the control group, the treated cells can enhance the phosphorylation of the Akt signal 24 hours before differentiation [80]. Overall, these results suggest that GLP-1 may regulate 3T3-L1 adipogenesis through the Akt signaling pathway.
GLP-1 has also been found to increase mRNA and protein expression levels of AP2 and PPARγ in a dose-dependent manner during pre-3T3-L1 adipocyte differentiation. The real-time reverse transcription polymerase chain reaction (RT-PCR) results showed that GLP-1 promoted the expression of C/EBPα and LPL [80]. Recent studies have also found that the promotion of GLP-1 on 3T3-L1 pre-adipocyte differentiation depends on the long non-coding RNA protein folding activity of the ribosome (lncRNA PFAR), which can target miR-138, remove the inhibition of miR-138 on Yes-associated protein 1 (YAP1) mRNA, and then upregulate the expression of YAP1 [81]. These data suggest that GLP-1 can promote the differentiation of pre-3T3-L1 adipocytes by promoting the expression of the specific markers LPL and AP2, as well as the transcription factors PPARγ and C/EBPα. Activation and phosphorylation of signaling pathways such as Akt, and yes-associated protein 1-TEA domain transcription factor (YAP1-TEADs) also play a key role in fat differentiation.
GLP-1 modulates fat browning
Adipose tissue is produced by the aggregation of a large number of fat cells. Adipose tissue is divided into WAT with a small number of mitochondria in the cell and BAT with a large number of mitochondria in the cell. WAT mainly plays the role of storing energy, while the main role of BAT is to decompose lipid droplets for energy. WAT can also be further divided into visceral adipose tissue (VAT) and subcutaneous adipose tissue [82]. Fat browning refers to the high plasticity of white fat cells, which can be transformed into beige fat cells with many similar morphological and functional characteristics to brown fat cells under the stimulation of exercise, cold exposure, adrenergic receptor stimulation, and other factors [83]. Since BAT has the function of thermogenesis and heat generation, promoting TG clearance and glucose metabolism. White fat browning is a good way to promote white fat consumption and accelerate human metabolism, which is associated with uncoupling protein 1 (UCP1), nitric oxide synthase (NOS), SIRT1, signal transducer and activator of transcription 3 (STAT3) (Fig. 3) [84].
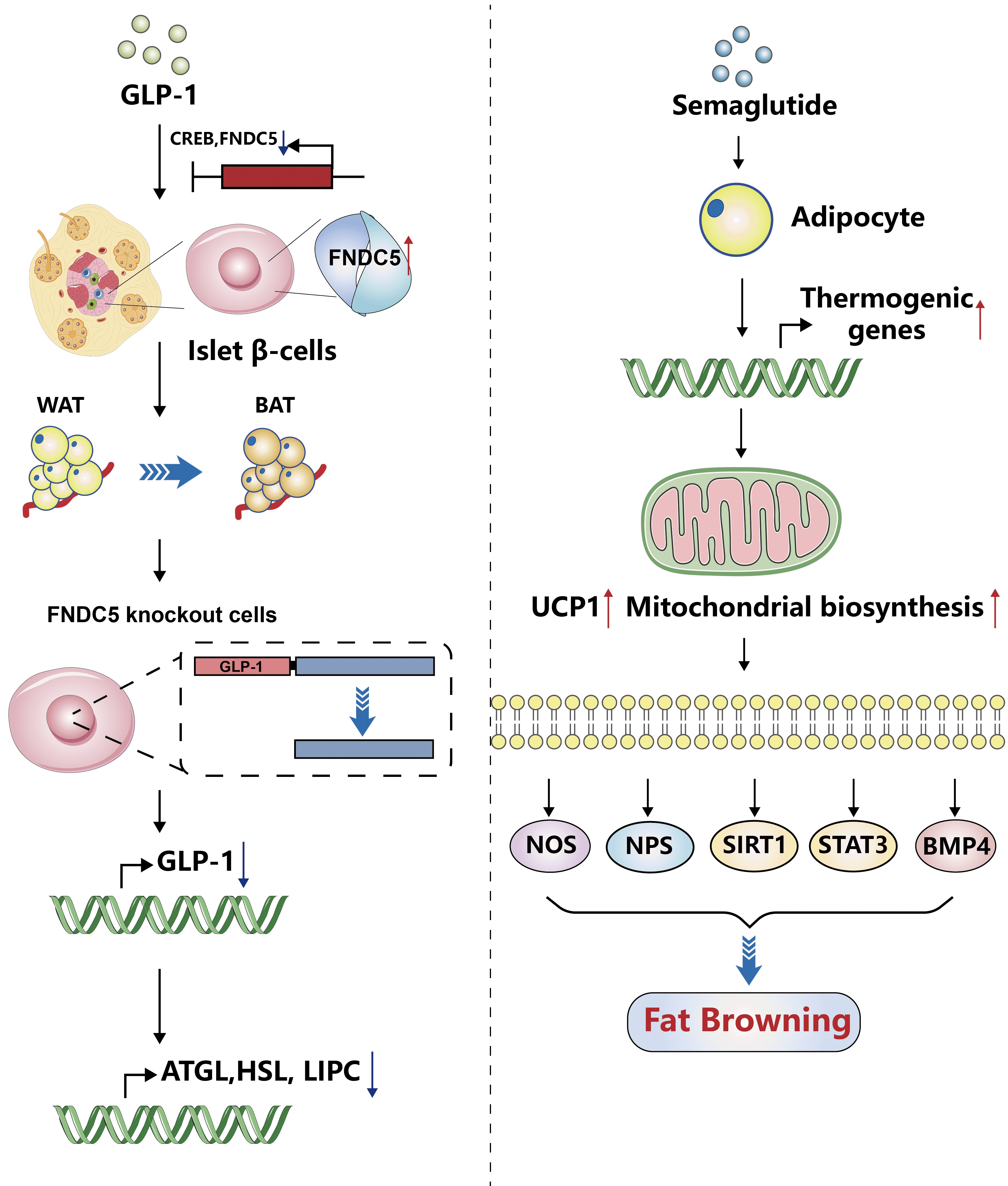
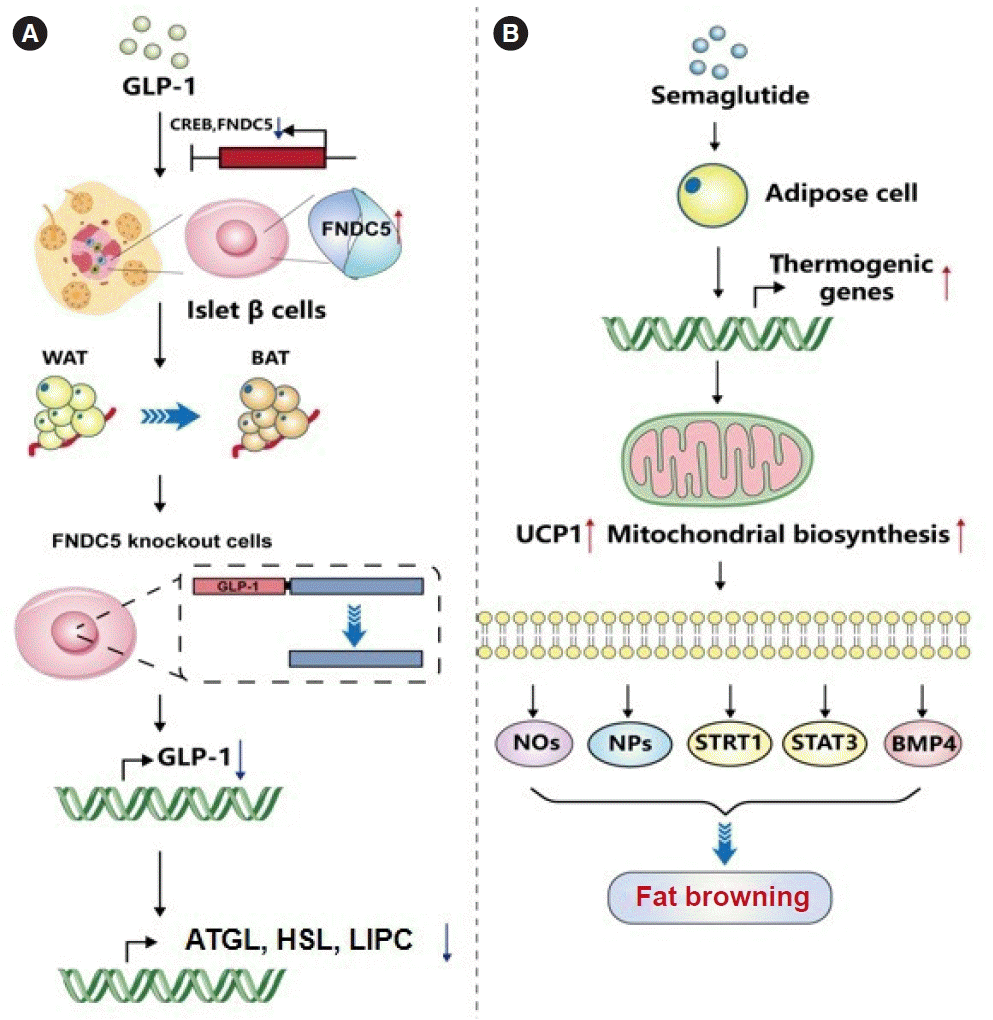 | Fig. 3.Possible mechanisms by which glucagon-like peptide-1 (GLP-1) modulates the browning of white fat. (A) GLP-1 can induce fibronectin type III domain-containing protein 5 (FNDC5) expression in pancreatic β-cells through the interaction of its downstream transcription factor cAMP-response element binding protein (CREB) with binding elements in the FNDC5 promoter, thereby promoting the conversion of white adipose tissue (WAT) to brown adipose tissue (BAT). GLP-1 expression was downregulated in wild-type β Lox5 cells knocked out by FNDC5, and the induced expression of lipolysis and autophagy-related genes and proteins were also inhibited, such as adipose triacylglyceride lipase (ATGL), hormone-sensitive lipase (HSL), and hepatic lipase deficiency gene (LIPC). (B) Semaglutide, as a GLP-1 analog, can directly act on fat cells, increase the expression of thermogenic genes for browning phenotype maintenance, improve the expression of uncoupling protein 1 (UCP1), mitochondrial biosynthesis, and thermogenic marker, or promote fatty browning through nitric oxide synthase (NOS), neuropeptides (NPs), sirtuin 1 (STRT1), signal transducer and activator of transcription 3 (STAT3), and bone morphogenetic protein 4 (BMP4) signaling.
|
The latest research pointed out that irisin is derived from its precursor protein fibronectin type III domain-containing protein 5 (FNDC5), which can be released from various tissues such as skeletal muscle, fat, and pancreas, mainly involved in the transformation of WAT to BAT, thereby increasing the body’s energy expenditure. GLP-1 can induce the expression of FNDC5 in pancreatic β-cells through the interaction of its downstream transcription factor cAMP-response element binding protein (CREB) with binding elements in the FNDC5 promoter, thereby promoting the conversion of WAT to BAT and enhancing energy metabolism [85]. In addition, GLP-1 expression was down-regulated in wild-type β Lox5 cells knocked out by FNDC5, and the induced expressions of lipolysis and autophagy-related genes and proteins were also inhibited, such as ATGL, HSL, and hepatic lipase deficiency gene (LIPC), suggesting that GLP-1 expression is also regulated by FNDC5, and may also provide therapeutic targets for lipid metabolism-related diseases.
Animal studies have shown that GLP-1RAs activate fat cell browning and may regulate BAT through the central nervous system. Martins et al. [86] found that semaglutide, as a GLP-1 analog, can directly act on fat cells, increase the expression of thermogenic genes for browning phenotype maintenance, improve the expression of UCP1, mitochondrial biosynthesis and thermogenic marker, thereby contributing to weight loss. So how does GLP-1 modulate BAT? Lockie et al. [44] observed the activity of BAT in the shoulder blade region of C57BL/6 mice by injecting GLP-1 (7–36) amide into the lateral ventricle of C57BL/6 mice. And they found that the mice lost weight, increased sympathetic activity, increased BAT temperature in the scapular region, and increased thermogenesis. However, the same dose of the above drug intervention was given by intraperitoneal injection, and there was no significant change in the temperature of BAT in the scapular area. This study suggests that GLP-1 can enhance the function of brown fat, increase heat production in BAT, and reduce body weight in mice by modulating the activity of the sympathetic system through binding with GLP-1R in the center. In recent years, extensive literature has reported that a variety of signaling pathways and transcription factors are involved in the fat differentiation process of WAT, such as the NOS pathway, neuropeptide S [87] pathway, SIRT1 [88] pathway, STAT3 [89], bone morphogenetic protein 4 (BMP4) [90] pathway, etc. These pathways and signaling pathways can promote BAT heat production. The above studies show that GLP-1 and GLP-1RAs can enhance BAT function or promote WAT browning through the center. However, the specific pathways or pathways involved are not clear enough and need further research and exploration.
Go to :
CLINICAL APPLICATION OF GLP-1 IN LIPID METABOLISM AND ADIPOSE-RELATED DISORDERS
Clinical application of GLP-1 in obesity
Obesity is a major health problem worldwide. Studies in human and animal models suggest that GLP-1 may play a role in lipid metabolism and obesity. Similar to rodents, GLP-1 also plays a key role in human lipid metabolism and obesity [91]. Most studies have found a decrease in GLP-1 secretion in obese individuals [92], and some studies have found no difference in GLP-1 levels between obese and normal-weight controls [93]. The mechanism of action of GLP-1RA to promote weight loss mainly includes the reduction of fat deposition through regulation of adipose tissue (lipolysis, fatty acid oxidation, and adipocyte differentiation) and the reduction of food intake through the central and peripheral nervous system.
In obese patients, acipimox can inhibit adipose tissue lipolysis by activating protein upregulated in macrophage by INF-γ (PUMA-G) (also known as the hydroxycarboxylic acid receptor 2 [HCA2] and GPR109a) receptors. In addition, an inverse correlation between circulating free fatty acids and GLP-1 concentrations have been found in vivo and in vitro experiments, and systemic GLP-1 levels can be increased in obese patients after treatment with acipimox [94]. Studies demonstrated that the fat-damaging effects of GLP-1 are effective for weight loss and may also improve insulin resistance in obese patients [95]. There is also evidence that GLP-1RA (such as Lira, etc.) can significantly reduce food intake and promote fatty acid oxidation, thus promoting weight loss and fat mass reduction [16,96]. In addition, in vitro models of primary human adipose stem cells, researchers detected intracellular fat content and quantitative expression of early and mature adipocyte markers (PPARγ, FABP4, and HSL), demonstrating that glucagon can significantly reduce adipocyte differentiation, inhibit adipocyte proliferation and adipogenesis. Moreover, this effect can be reversed by specific GLP-1R (exendin-9) antagonists, which means that GLP-1RA may inhibit the proliferation and differentiation of human adipocyte precursors [97,98]. These studies suggest that GLP-1RA can achieve weight loss by interfering with the proliferation and differentiation of human fat precursors.
Besides, Baggio and Drucker [99] and Allegretti et al. [100] investigated the relationship between intestinal flora and obesity. They found that intestinal flora may mediate the release of the intestinal hormone GLP-1 through the short-chain fatty acid (SCFA)-GPCR pathway, thereby increasing satiety and slowing gastric emptying, ultimately reducing obesity. There is also evidence that GLP-1RA has positive effect on metabolic syndrome and abdominal obesity. In a recent randomized, double-blind, placebo-controlled trial, obese adults without diabetes were assigned to the exercise group, the GLP-1RA (Lira) group, and the exercise+Lira group. The results showed that abdominal fat percentage decreased by 2.6, 2.8, and 6.1 percentage points in the exercise, Lira, and combination groups compared to placebo. The combination of adherent exercise and Lira treatment reduced abdominal obesity and may therefore reduce obesity risk more than the individual treatments [101]. The difference between elevated or unchanged GLP-1 levels in obese patients may be related to differences in metabolic status and detection methods, but the significant correlation between GLP-1 and lipid metabolism suggests that it can control weight and regulate lipid metabolism in HF diet groups, and GLP-1 has the potential to treat obesity caused by HF diet [102].
Clinical application of GLP-1 in fatty liver
NAFLD is one of the most common metabolic syndromes in today’s society and has become the common chronic liver disease that seriously affects the health of adults and even children worldwide. There are currently no specific agents for NAFLD [103]. Previous studies have shown that GLP-1 can inhibit the formation and progression of NAFLD by enhancing lipid metabolism through oxidation and reducing the influx of fatty acids into hepatocytes [104]. The mechanisms of GLP-1 to improve NAFLD include improving lipid metabolism by regulating metabolism-related signaling pathways such as AMPK pathway [33] and JNK pathway [105]. And GLP-1 can inhibite hepatocyte apoptosis which induced by NLRP3 through autophagy in mitochondria, weakening the intrahepatic inflammatory response, thereby inhibiting the transition from simple fatty liver to nonalcoholic steatohepatitis (NASH). Finally, GLP-1 can achieve the purpose of preventing the deterioration of NAFLD [106,107]. Recent research data has shown that the expression of lipid metabolism genes adiponectin (APN), Adipo R, AMPK, PPARα mRNA and protein is significantly increased after 48 hours of treatment with different concentrations of GLP-1 compared with the model group of adipose hepatocytes [108]. GLP-1 alleviates hepatocyte steatosis and reduces intracellular lipid droplets by activating the expression of the above lipid metabolism genes. This also shows that GLP-1 can significantly improve clinical indexes, reduce inflammatory indicators, improve liver fat deposition, and be effective in the treatment of fatty liver.
In addition, Sharma et al. [64] studied GLP-1 and receptors on hepatocytes and found that the combination of them can reduce the lipid stress response in hepatocytes, reduce the lipid load of hepatocytes, delay steatosis, and ultimately prevent NAFLD. Several other studies have also provided evidence that GLP-1RA can reduce hepatic fat accumulation and reduce steatosis in liver tissue by regulating the expression of genes and proteins related to the fat synthesis pathway [109,110]. However, there are no targeted drugs specifically designed to treat NAFLD. According to this characteristic, GLP-1RA gradually entered the clinic. Hopkins et al. [111] treated T2DM patients with exenatide and found that the liver fat content (LFC) of patients decreased from 21.3%±19.3% to 12.7%±10.6%, a trend that suggested that GLP-1RA drugs could improve liver lipid metabolism. Similarly, Gastaldelli et al. [112] found a decrease in LFC in patients with moderate to severe NAFLD with T2DM using exenatide in combination with dapagliflozin. Additionally, a further follow-up study was conducted in patients with poor T2DM control and found a significant 31% reduction in LFC in patients treated with Lira. Recent studies suggest that treatment with semaglutide (GLP-1RA) in combination with firsocostat (ACC-a inhibitor) and/or cilofexor (nonsteroidal FXR agonist) can further improve hepatic steatosis and thus NAFLD compared with semaglutide alone in exploratory efficacy analyses [113]. Flint et al. [114] randomized subjects to semaglutide or placebo to observe the change of liver fat from baseline to week 48, and found that semaglutide significantly reduced hepatic steatosis, improved liver enzymes and metabolic parameters, and had a positive effect on the diagnosis and treatment of NAFLD compared with placebo. The above studies suggest that GLP-1 can significantly reduce LFC and inflammatory response in patients with T2DM and NAFLD, and even slow down the process of liver fibrosis [115]. And the emergence of GLP-1RA, such as semaglutide, exenatide, and efinopegdutide [116], and so on, can accelerate the drug development of NAFLD which has great therapeutic potential for the treatment of NAFLD [117].
Interestingly, there are studies on the safety and effectiveness of Lira in the treatment of NAFLD. It is found that in the clinical follow-up studies of NAFLD patients, compared with the control group, hepatic fat content and ALT levels were not significantly reduced in Lira group and were associated with an increased incidence of gastrointestinal adverse events [118]. These studies suggest that despite the potential benefits of Lira, current evidence does not advocate the use of Lira in patients with NAFLD.
In addition to NAFLD, NASH is also of concern, with limited treatment options. If NASH is left untreated, the liver can deteriorate over time. In a recent study, Newsome et al. [119] conducted a phase II clinical trial in patients with NASH, that is, a placebo-controlled trial of subcutaneous sommarutide in the treatment of NASH, and found that sommarutide treatment resulted in a significantly higher proportion of patients in remission of NASH compared with placebo. A number of studies have also validated the therapeutic potential of sommarutide in NASH [113,120,121]. While sommarutide is undoubtedly an exciting potential treatment option, there is still a lack of clear clinical data on the mechanism of action of GLP-1 agonists in NASH, such as whether GLP-1 agonists have a direct, independent role in improving NASH, or simply by improving body weight, insulin resistance, and so on.
Clinical application of GLP-1 in atherosclerosis
There is substantial evidence that GLP-1 is involved in the function of the cardiovascular system [9]. GLP-1 has multiple beneficial effects in cardiovascular diseases such as hypertension, myocardial ischemia, atherosclerosis, and heart failure. And it may prevent the occurrence of cardiovascular disease. This article mainly reviews the anti-atherosclerotic effect of GLP-1 in clinical applications. A clinical trial of sitagliptin in a small sample of patients with impaired glucose tolerance and mild T2DM found significant reductions in carotid intimal thickness in sitagliptin compared with diet control alone [122]. In addition, a multicenter, randomized controlled trial in patients with T2DM without cardiovascular disease also found that the mean carotid intimal thickness and left carotid intimal thickness after sitagliptin treatment were significantly reduced from baseline, with no significant difference on the right side. The results suggested that sitagliptin has a delayed effect on the progression of carotid atherosclerosis (CAS). This is consistent with the results of GLP-1 in animal experiments conducted by Burgmaier et al. [123] to reduce intraplaque inflammation and increase plaque stability. Also, data analysis showed that the serum GLP-1 level of patients with newly diagnosed T2DM combined with CAS was significantly reduced compared with the control group. And it was related to the severity of CAS. GLP-1 levels can predict CAS in patients with newly diagnosed T2DM to some extent, and the optimal GLP-1 level of it is 6.59 pmol/L. However, Koska et al. [124] conducted a double-blind, practical trial in which participants were randomly assigned to receive exenatide or placebo to measure changes in carotid plaque volume and composition at 9 and 18 months of treatment, and found no difference in mean change in plaque volume in the exenatide group compared with placebo. This suggests that the antiatherosclerotic effects of short-term GLP-1 may not play a central role in atherosclerosis. Therefore, studies that extend the follow-up time and increase the sample size to observe the role of GLP-1 in atherosclerosis can be further developed. It can also provide new ideas for the prevention and treatment of cardiovascular diseases.
Go to :
GLP-1 AND DUAL AGONISTS
Effects of glp-/glucose-dependent insulinotropic peptide dual agonists on lipid metabolism
GLP-/glucose-dependent insulinotropic peptide (GIP) dual agonists are a new type of oral hypoglycemic drug, which can activate both GLP-1 and GIP incretin at the same time, thereby exerting a more comprehensive blood glucose control effect [125]. Recent studies have shown that GLP-/GIP dual agonists also have significant lipid-regulating effects. Frias et al. [126] demonstrated in a drug clinical study in 2018 that GLP-/GIP dual agonists could significantly reduce TG and TC levels in patients compared with placebo. In mouse models of NASH, GLP-/GIP dual agonists have also been found to significantly reduce TG and LDL-C and increase high-density lipoprotein cholesterol (HDL-C) [127]. Specifically, tezepatide can reduce TC, LDL-C and TG levels by 12.9%, 15.5%, and 24.9%, respectively, and increase HDL-C levels by 0.9% [128]. In addition, more and more studies have proved that such drugs can reduce TC, LDL-C and TG, and increase HDL-C, so as to effectively correct lipid metabolism disorders [129].
In rodent models, a number of studies have found that GLP-/GIP dual agonists group mice have reduced fat mass, decreased circulating cholesterol, and lost weight, which has excellent weight loss effect. Furthermore, the weight loss effect of GLP-/GIP dual agonists is achieved by combining an increase in energy expenditure with a decrease in food intake [130], and its potential in the treatment of obesity has been validated in clinical trials. In addition, a large number of clinical studies have shown that GLP-/GIP dual agonists can reduce LFC, VAT, and body weight, and have an improvement effect on both NAFLD and NASH [131,132]. These studies all showed that GLP-/GIP dual agonists had similar effects on lipid metabolism to GLP-1, and even had better effects on lipid reduction and weight loss.
Comparison of GLP-/GIP dual agonists and GLP-1
GLP-1 and GIP, both major members of the incretin family, play a physiological role by binding and activating corresponding receptors to promote postprandial insulin secretion [125]. GIP consists of 42 amino acids and is synthesized and secreted by endocrine K cells located mainly in the proximal duodenum and jejunum. GIP receptor (GIPR) is also a type B G-protein-coupled receptor, which exists in various tissues such as pancreas, adipose tissue, gastric mucosa, heart, adrenal cortex, bone and brain. Binding of GIP to its receptor also stimulates glucose-dependent insulin secretion and produces a greater proportion of the incretin effect than GLP-1 (about 45% for GIP and 29% for GLP-1) [133]. It is worth mentioning that, GLP-1 inhibits glucagon secretion in α-cells during hyperglycemia and normal glycemia states, but does not have this effect at low glycemia levels. Unlike GLP-1, GIP can promote glucagon secretion in normal and hypoglycemic states, and inhibit it in hyperglycemia, thus acting as a bifunctional hormone to maintain blood glucose homeostasis. In addition, GIP enhances the lipid buffering capacity of WAT in lipid metabolism and promotes suitable lipid allocation, thereby reducing ectopic fat deposition [134]. However, GLP-1 regulates lipid metabolism by enhancing the function of BAT or promoting WAT browning. GLP-/GIP dual agonist innovatively integrates the effects of two incretin into one molecule, which can simultaneously activate GLP-1R and GIPR. The main representative drug of it is tirzepatide [135], which is a representative dual agonist commonly studied. There are also many multi-target dual agonists under development, such as GLP-1R/GCGR, GLP-1R/fibroblast growth factor 21 receptor (FGF21R), etc. Compared with GLP-1, dual agonists can avoid the degradation of dipeptidyl peptidase-4, thereby prolongating drug administration. A number of studies have shown that GLP-/GIPR dual agonists can achieve better blood glucose control, insulin sensitivity, lipid metabolism and weight loss compared with GLP-1 alone [136,137]. GLP-/GIPR dual agonists have fewer side effects and greater safety, which means that the combination therapy has greater metabolic benefits than single use. Moreover, although GLP-1 has a significant effect on blood glucose control, its effect on lipid metabolism is relatively weak. In contrast, GLP-/GIP dual agonists, while controlling blood sugar, can more comprehensively regulate lipid metabolism and reduce the risk of cardiovascular diseases [138,139]. In addition, all currently available GLP-1R therapies are peptide agonists, most of which require subcutaneous administration [140]. However, GLP-/GIP dual agonists have been proved to be orally effective in animal experiments, and its state in plasma is more stable than GLP-1RA [141], and this oral administration method has gradually been applied to the clinic [142]. Therefore, GLP-/GIP dual agonists have advantages in terms of administration mode, and their oral administration mode is more convenient than GLP-1 injection administration. The development of double agonists and even multiple agonists has also improved the limitations of GLP-1R in the treatment of metabolic diseases and provided better treatment options.
In summary, GLP-/GIP dual agonists have shown superior potential over GLP-1 in improving lipid metabolism. With the deepening of clinical research, we expect that this kind of new drug can bring more comprehensive treatment benefits to the majority of diabetes patients. At the same time, GLP-/GIP dual agonists may also be an effective treatment option for non-diabetic patients with abnormal lipid metabolism.
Go to :
CONCLUSIONS
In summary, the research progress of GLP-1 in lipid metabolism shows its potential in the treatment of metabolic disorders such as NAFLD, especially combined with its significant weight loss effect, making the application of GLP-1 in obese patients with NAFLD more worthy of in-depth exploration. However, the use time and effect of GLP-1RAs, and the adverse reactions after long-term use must also be fully paid attention to and studied before clinical application. In addition, attention should also be paid to the effect of GLP-1 at different sites of administration and blood concentrations. Therefore, basic and clinical trials further reveal the mechanism of GLP-1 on lipid metabolism, which is helpful to improve our understanding of GLP-1 function and is essential for advancing its clinical application. The development of GLP-/GIP dual agonists provides a more complete scheme for the treatment of metabolic diseases, but the safety of more dual agonists or multiple agonists in clinical trials is also worthy of attention.
Go to :
 XML Download
XML Download