

 PDF
PDF Citation
Citation Print
Print


Introduction
Pseudomonas aeruginosa, one of the antimicrobial-resistant ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, P. aeruginosa, and Enterobacter species) pathogens, represents a global threat to human health [1]. P. aeruginosa usually shows multidrug resistance. Its co-resistance to carbapenems, aminoglycosides, polymyxins, and tigecycline has also increased [2]. There are limited treatment options for serious infections caused by multidrug-resistant P. aeruginosa. Many combination therapies have been tried [3]. Recently, it has been reported that combination therapy including ciprofloxacin is correlated with a lower mortality. A combination of a beta-lactam and ciprofloxacin has been proposed for ciprofloxacin-susceptible P. aeruginosa bacteremia [4].
According to data from the Korean global antimicrobial resistance surveillance system (Kor-GLASS) [5], of P. aeruginosa blood isolates in 2017-2019, 12.3% showed resistance to piperacillin, 13.1% to ceftazidime, 12.9% to cefepime, 21.3% to imipenem, 22.1% to meropenem, 9.6% to amikacin, and 18.9% to ciprofloxacin. Most the carbapenemase-producers exhibited co-resistance to amikacin. However, data about its co-resistance to ciprofloxacin are unavailable. This study provides the newest data about the resistance mechanism to fluoroquinolone of carbapenemase-producing P. aeruginosa.
Resistance mechanisms of P. aeruginosa to fluoroquinolone are known mostly through the acquisition of mutations in genes encoding target proteins of fluoroquinolone and regulators of efflux pumps, leading to overexpression of these pumps [6]. Quinolone resistance may also be attributable to mutations of target enzymes of topoisomerases II and IV encoded by gyrA and parC, respectively [7]. These mechanisms are known to be chromosomally mediated. However, plasmid-mediated quinolone resistance (PMQR) in P. aeruginosa is also important. It is associated with qnrA-E, qnrS, qnrVC, qepA, oqxAB (efflux pump), and acc(6′)-Ib-cr (quinolone-modifying enzyme) [7]. Recently, CrpP enzymes have been described as a novel ciprofloxacin-resistance mechanism. The crpP gene encodes a novel protein, capable of specifically conferring resistance to ciprofloxacin in Escherichia coli through an adenosine triphosphate-dependent mechanism that involves phosphorylation of the antibiotic [8].
The purpose of this study was to report on the current situation of crpP gene spread in carbapenemaseproducing P. aeruginosa blood isolates in Korea.
Materials and methods
In this study, non-duplicated P. aeruginosa blood isolates were isolated from nine nationwide general hospitals (National Health Insurance Service Ilsan Hospital, Gangnam Severance Hospital, Chonnam National University Hospital, Chungbuk National University Hospital, Busan Paik Hospital, Jeju National University Hospital, Hallym University Dongtan Sacred Heart Hospital, Wonju Severance Christian Hospital, Keimyung University Dongsan Hospital) and two nursing homes according to the Kor-GLASS manual [5] in 2020. Briefly, pure colonies of P. aeruginosa were collected in 10% skim milk and stored at -70°C before all collected isolates were transferred to a single analysis center of the Korea Disease Control and Prevention Agency for analyses using approved methods [5]. Bacterial species were verified using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight mass spectrometry (Bruker Biotyper, Bruker Daltonics GmbH, Bremen, Germany). Antimicrobial susceptibility was mainly determined by the disk diffusion test according to the Clinical and Laboratory Standards Institute (CLSI) guidelines [9].
To detect carbapenemase-producers, P. aeruginosa isolates showing nonsusceptibility to imipenem or meropenem were PCR-sequenced to detect blaKPC , blaNDM , blaOXA-48 , blaVIM , blaIMP , and blaGES . For wholegenome sequencing, DNAs of freshly subcultured isolates were extracted using a GenElute™ Bacterial Genomic DNA Kit (Sigma-Aldrich, St. Louis, MO, USA). Then 8 μg of input genomic DNA was used to sequence the entire genome using NextSeq 550 instrument (Illumina, San Diego, CA, USA). Sequences were assembled with Spades (version 3.11.1) and annotated with Prokka (version 1.13.7). Resistance genes were obtained with ResFinder 4.1 from the website of the center for genomic epidemiology [10]. A phylogenic tree was generated based on whole-genome multilocus sequence typing using a BioNumerics software, version 7.6.3 (Applied Maths, St Martens Latem, Belgium) [11] (Fig. 1).
Results
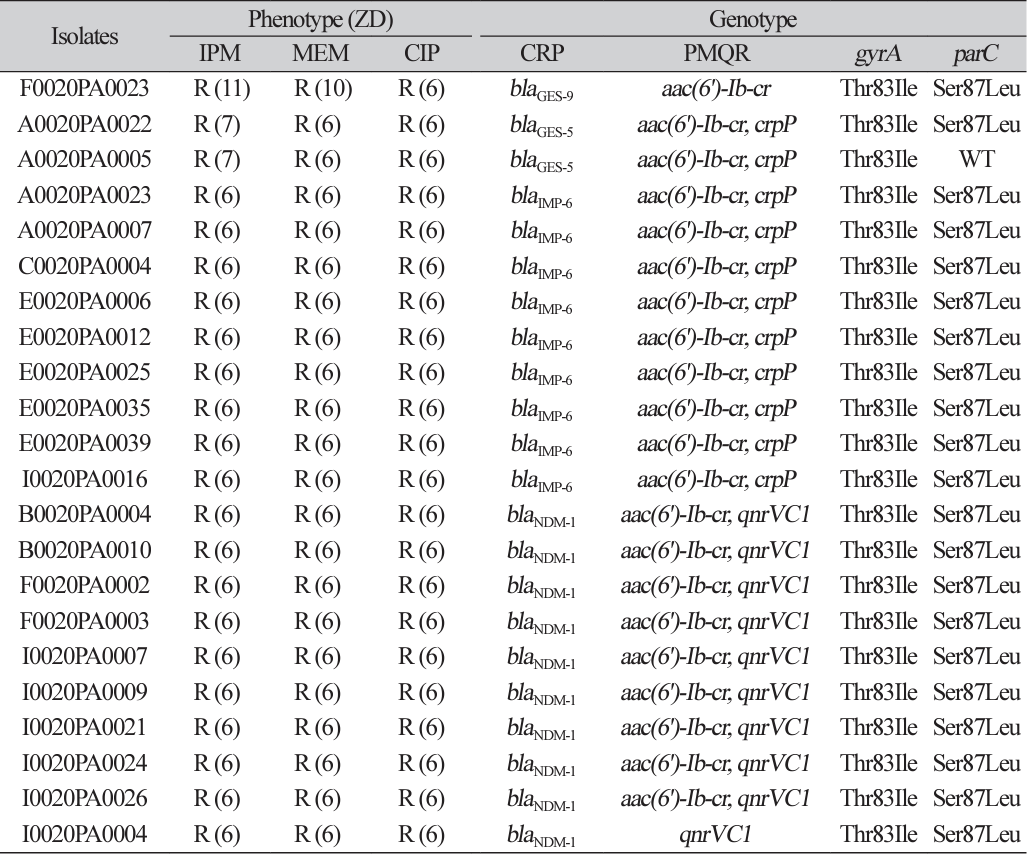
Among 212 non-duplicated P. aeruginosa blood isolates, 22 carbapenemase-producing P. aeruginosa isolates showed resistance to both ciprofloxacin and imipenem. A total of 11 carbapenemase-producing P. aeruginosa blood isolates (nine IMP-6 producers and two GES-5-producers) co-harbored aac(6')-Ib-cr and crpP (Table 1). A total of nine NDM-1-producers co-harbored aac(6')-Ib-cr and qnrVC1. One GES9-producer also carried aac(6')-Ib-cr and one NDM-1-producer also carried qnrVC1. The phylogenic tree showed no epidemiologic link among 22 carbapenemase-producing P. aeruginosa isolates.
Discussion
Quinolones have been commonly used as a treatment option for a large number of bacterial infections. Ciprofloxacin is the most active antibiotic in this group. The increase in quinolone resistance has limited the effect of quinolones. There was no report about the prevalence of quinolone resistance genes in P. aeruginosa in Korea to the best of our knowledge. Recently, novel mutations in gyrA and parC genes were first found in P. aeruginosa isolated from companion dogs in South Korea [12]. In that study, a total of 84 nonduplicated P. aeruginosa strains were obtained from healthy dogs and infected dogs. The resistance rate was 14.3% for levofloxacin and 13.1% for ciprofloxacin. The resistance was commonly associated with gyrA mutations [12].
The information about the prevalence of quinolone resistance genes of clinically important pathogens in Korea is limited. For K. pneumoniae, mutations in gyrA and parC were found in 78.9% and 65.5% of 142 extended-spectrum β-lactamases-producers, respectively [13]. The common PMQR gene was qnrB-aac(6')Ib-cr-oqxAB (58/142, 40.8%) [13]. Lee et al. [14] have reported the presence of PMQR genes in Salmonella enterica isolated from human salmonellosis patients in South Korea from 2016 to 2019 [14]. Among 34 Salmonella strains with reduced susceptibility to quinolones, 25 strains harbored one or two of qnrA, qnrB, qnrS (most common), and aac(6')-Ib-cr genes.
Out of 22 carbapenemase-producers from nationwide collections, we detected 11 P. aeruginosa blood isolates that co-harbored crpP and carbapenemase genes. This is the first report about the current situation of crpP gene spread in carbapenemase-producing P. aeruginosa blood isolates in Korea. Because all strains also have aac(6')-Ib-cr and chromosomal mutations in gyrA or parC, the effect of the presence of crpP gene on quinolone resistance is hardly speculated in this study.
The crpP gene obtained from the pUM505 plasmid isolated from a P. aeruginosa clinical isolate was identified in 1986 [8]. Regarding the expression of CrpP in the transconjugants test, CrpP proteins increased minimal inhibitory concentration values of ciprofloxacin concerning E. coli J53-3. However, these changes were not enough to be considered as resistance according to the breakpoint value (≥ 4 mg/L) reported for this interpretation by the CLSI [8]. Nevertheless, the crpP gene conferring low-level resistance could facilitate the selection of mutants with a higher level of quinolone resistance. In addition, plasmids contain other genes in addition to the crpP gene, which encodes additional quinolone resistance mechanisms [8].
In this study, the type of carbapenemase of crpP gene-carrying P. aeruginosa was mostly IMP-6, known to be a common type in Korea [15]. NDM-1-producing P. aeruginosa isolates did not carry crpP genes, which seemed to become one of the majority types of carbapenemase in P. aeruginosa. The shift in the molecular epidemiology of carbapenemase genes might change the molecular epidemiology of PMQR genes.
Mobile genetic elements play a key role in the spread of resistance genes, and high-risk clones frequently integrate such determinants into their genomes [16]. In this study, the linkage of crpP and aac(6')-Ib-cr gene with blaIMP-6 is suspected. And there is high possibility that aac(6')-Ib-cr and qnrVC1 are associated with blaNDM-1 in P. aeruginosa.
If only PMQR is present, it is expected to be susceptible or low-grade resistance phenotypes [7,8]. Unfortunately, the only effect of crpP gene is hard to know in this study, because all isolates had chromosomal mutations in gyrA or parC, in addition to PMQRs. These strains are usually associated with a large reduction in biological fitness with the accumulation of more resistance-associated mutations [17]. One more thing to consider is that NDM-1 carrying P. aeruginosa is mostly separated from hospitals in Daegu or Busan and the regional differences could have an effect. Therefore, we suggest continuous monitoring of the change of PMQR in P. aeruginosa.
In conclusion, the spread of P. aeruginosa isolates, coharboring crpP and carbapenemase genes were found in this study. This is the first report about the current situation of crpP gene spread in carbapenemaseproducing P. aeruginosa blood isolates in Korea.

 XML Download
XML Download