

 PDF
PDF Citation
Citation Print
Print


Introduction
Most Plasmodium vivax infection in Korea occurs near the demilitarized zone (DMZ). From 2014, about 500 cases of P. vivax infections have occurred in Korea every year. Long-term military chemoprophylaxis since 2005 has contributed to a decrease in malaria cases. However, this might have facilitated the development of chloroquine (CQ)-resistant P. vivax strains [1].
CQ has been used for decades as a first-line treatment for uncomplicated P. vivax cases. Since the first report of CQ treatment failures in P. vivax malaria in Papua, Indonesia in 1989 [2,3], reports of antimalarial treatment failures related to genetic polymorphisms in P. vivax dihydrofolate reductase (pvdhfr), P. vivax multidrug resistance protein 1 (pvmdr1) and P. vivax hydroxymethylpterin pyrophosphokinasedihydropteroate synthetase (pvdhps) have increased, causing worldwide concerns about drug-resistant strain spread [4-9]. Especially, strains with multi-polymorphisms are thought to be the cause of antimalarial treatment failure. Despite the continuous occurrence of malaria infections in Korea, studies of CQ resistance polymorphisms were not reported. Here, we profiled known polymorphisms of P. vivax genes associated with drug resistance from clinical specimens in a secondary hospital in Korea.
Go to : 
Materials and methods
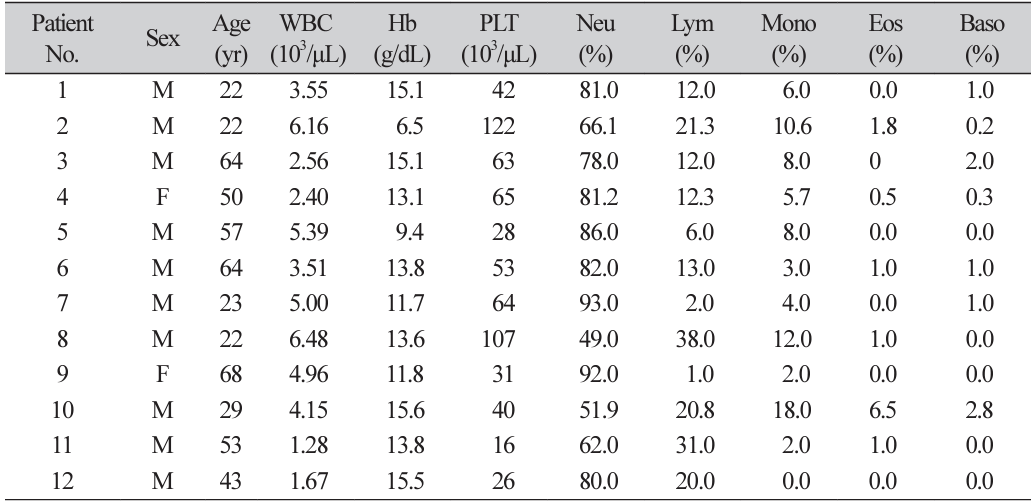
Patients with confirmed P. vivax infections in Uijeongbu St. Mary’s Hospital were enrolled from July 2014 to June 2017. A total of 12 P. vivax infection samples were collected. P. vivax infections were confirmed through SD Bioline Malaria Ag Pf/Pan™ rapid diagnosis test kits (Abbott, Chicago, IL, USA) and referral Plasmodium spp. PCR (polymerase chain reactin; SMLab, Seoul, Korea). Electronic medical records and complete blood count (CBC) findings were reviewed for all patients. In the 2015 World Health Organization (WHO) guidelines [10], antimalarial drug resistance is defined as the survival or increased number of parasites despite the use of an antimalarial drug at an usual recommended dose. According to the definition by the WHO, treatment failure in this study was defined as the persistence or an increase in parasite count despite the first-line therapy.
DNA extraction, PCR, and Sanger sequencing of pvdhfr, pvmdr1, and pvdhps genes were performed following methods reported in previous studies [6,11-13]. The following primers were used for PCR and Sanger sequencing [13]: primer PV1F (sense) 5′-CAGTGAAGGGACAAAGAATGAACC-3′, primer PV1R (antisense) 5′-ACTCGGGGAAGAAGACGTCAC-3′ for pvdhfr (560 bps), primer PV7F (sense) 5′-GCCATGTTCATTTCTGAGACGCTG-3′, primer PV7R (antisense) 5′-TCGCTCTGATGGCAAACACTC-3′ for pvmdr1 (337 bps), primer PV9F (sense) 5′-GCGGTTTATTTGTCGATCCTGTG-3′ and primer PV9R (antisense) 5′-TTTTTCCTGGCATCACTTGCTG-3′ for pvdhps (244 bps). Amino acid sequences were compared with reference wild-type sequences, where insertions and deletions were manually verified. Identified single-nucleotide polymorphisms (SNPs), insertions, and deletions were compared with reported SNPs known to be associated with antimalarialresistance in previous studies [5,6,8,11,14,15].
Go to :
Results
Ten of 12 patient were males. Initial hemoglobin level varied. CBCs on the first hospital visit of the twelve patients showed no specific pattern, although all cases showed cytopenia including various degrees of thrombocytopenia (Table 1). All 12 specimens had polymorphisms in pvdhfr. Only one out of 12 specimens had a polymorphism in pvdhps. No polymorphism was present in pvmdr1. Four previously reported nonsynonymous polymorphisms (F57L, S58R, H99S, S117N) and one previously reported in-frame deletion (c.292_309del, = T98A) were detected in pvdhfr. A nonsynonymous polymorphism (M399I) was detected in pvdhps. F57L in pvdhfr was only found as double polymorphisms, with c.292_309del found in 5 out of 12 cases. Two cases had C.292_309del and three cases had H99S in pvdhfr. One case had both S58R and S117N in pvdhfr. Only one case had M399I in pvdhps, with C.292_309del in pvdhfr (Table 2). All species with H99S did not have tandem repeat insertions or deletions (e.g., C.292_309del) in pvdhfr.
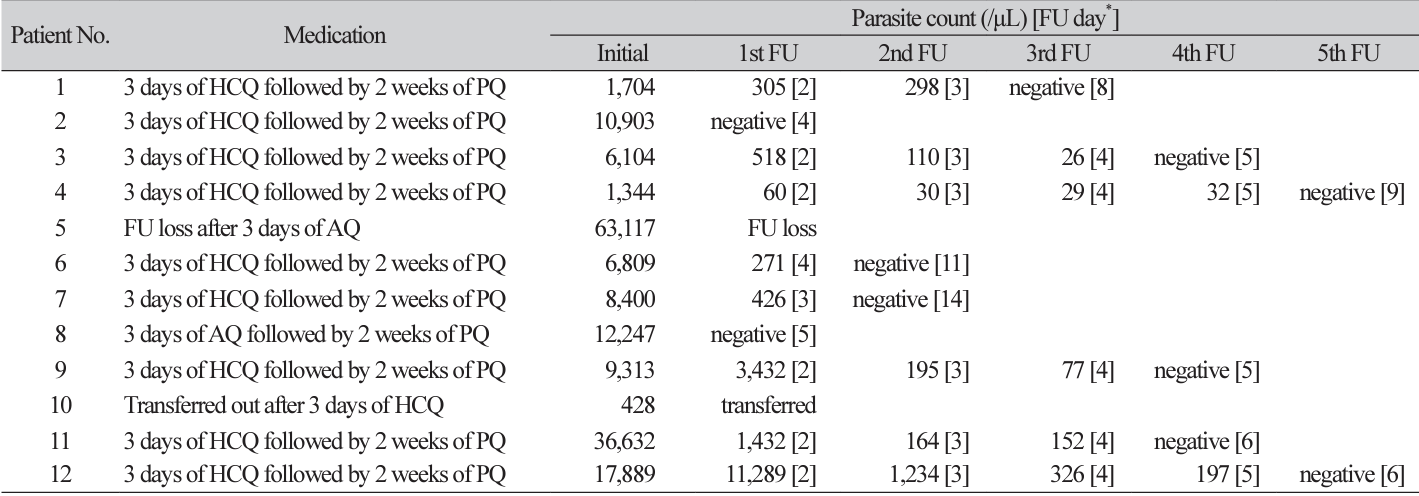
Nine out of 12 cases were cured with first-line regimen of three days of hydroxycholoroquine (HCQ) followed by two weeks of primaquine (PQ). The patient No. 8 with the S58R/S117N double polymorphism in this study recovered after three days of atovaquone (AQ) followed by two weeks of PQ. Patient No. 10 who was transferred out after three days of HCQ was confirmed to be cured later (Table 3).
One of the three H99S patients was lost to follow-up after the first visit. However, the other two patients recovered after three days of HCQ followed by two weeks of PQ. All c.292_309del cases with or without F57L in pvdhfr recovered after three days of HCQ followed by two weeks of PQ. The patient with c.292_309del in pvdhfr and M399I in pvdhps was transferred out after three days of HCQ medication. It was confirmed that this patient recovered later.
Go to :
Discussion
Among polymorphisms detected in pvdhfr (F57L, S58R, H99S, S117N, and c.292_309del), the S58R/ S117N double polymorphism has been previously reported in Indonesia, Thailand, Ethiopia, China, East Timor, Philippines, Vanuatu, Vietnam, Papua New Guinea, Madagascar, Iran [16], and French Guiana with in vitro resistance to drug combination of sulfadoxine and pyrimethamine (SP) [11]. Because the S58R/ S117N double polymorphism case in this study was treated with AQ and PQ, we could not assume that this polymorphism resulted in SP or CQ resistance in vivo.
It has been reported that the F57L polymorphism in the pvdhfr gene can increase resistance of P. vivax to antifolate agents such as pyrimethamine [4,9]. F57L strains are known to be less prevalent in the region where CQ is used as a first-line regimen [17], which means low selection pressure of antifolate agents. In a previous study [18], pvdhfr F57L was found in 23% (22/97) of P. vivax strains in Korea. In the present study, it was found in 42% (5/12) of P. vivax strains (Table 2). This might be due to the difference in sample size. Actual change within the period between the two studies is also possible.
M399I in the pvdhps gene was reported in a previous report [19], although its association with drug resistance was unknown. This polymorphism was unlikely to cause in vivo CQ resistance because the patient recovered after HCQ medication (Table 3).
Unlike previous reports from other geographic locations, multi-polymorphisms exceeding double polymorphisms were not found in the present study. These reports of multi-polymorphisms are from the region in which antimalarial treatment failures and are thought to be the cause of treatment failures [11,15,16]. This justifies further monitoring of genetic profile and drug resistance of P. vivax in Korea.
Although every specimen had single or double polymorphisms known to cause antimalarial drug resistance, there was no reported treatment failure with first-line therapy. Because all cases except two with AQ therapy were cured with first-line HCQ therapy and none of the 12 cases had polymorphism in pvmdr1, the current regimen of three days of HCQ followed by two weeks of PQ seems to be proper as a first-line therapy in Korea.
To the best of our knowledge, this is the first study analyzing polymorphisms in pvdhfr, pvdhps and pvmdr1 genes altogether in Korea. Although all cases were susceptible to first-line therapy, pvdhfr F57L proportion in this study was higher than that in a previous report. Whether this change was true without a selection pressure by antifolate drugs or it was just an error due to small sample size was unclear. This is a limitation of this study. To clarify this change and to surveil emergence of multi-polymorphism strain reported abroad, further monitoring of genetic profile and drug resistance of P. vivax is needed.
Go to :

 XML Download
XML Download