

 PDF
PDF Citation
Citation Print
Print



Introduction
Acute kidney injury (AKI) is a common clinical condition that affects individuals of all ages ranging from children to those who are older [1]. Hospitalized patients, especially those in intensive care units, are at higher risk of developing AKI because of conditions such as surgeries, infections, and medications that can trigger kidney injury. AKI is associated with high morbidity and mortality rates. Severe AKI may progress to end-stage kidney disease or require kidney replacement therapy [2]. Nevertheless, therapies for AKI remain conservative, highlighting the urgent need for innovative therapeutic approaches.
Kidney ischemia-reperfusion (IR) injury is a pathological condition characterized by an initial reduction in blood supply to the kidneys and subsequent reoxygenation [3]. Counterintuitively, this restoration of blood flow often exacerbates tissue damage, resulting in glomerular and tubular damage, and triggering a severe inflammatory response, termed “reperfusion injury.”
Mitochondria, the powerhouses of cells, are pivotal for adenosine triphosphate (ATP) generation through oxidative phosphorylation. In normal mitochondrial physiology, the process encompasses not only ATP synthesis via the tricarboxylic acid cycle and redox management but also the handling of reactive oxygen species (ROS). Additionally, processes such as mitochondrial biogenesis and dynamics involving fission, fusion, and distribution are crucial for balancing mitochondrial network maintenance, quality control, and mitochondrial DNA distribution, which are linked to metabolic status [4]. Cellular and organismal health depends on this balance, with significant alterations occurring in response to stress and disease. The mitochondrial death pathway also plays a vital role in maintaining mitochondrial integrity and overall cell health [5].
Emerging research suggests that mitochondrial dysfunction plays a pivotal role in the pathogenesis of kidney IR injury [6]. The kidneys are high energy-demanding organs because of their roles in filtration, reabsorption, and secretion [7], and they depend heavily on mitochondria. Because mitochondria are primary suppliers of cell energy in the form of ATP, their dysfunction can critically impair the energy supply required for optimal kidney function [6]. Recent studies have suggested that the aberrant ROS levels implicated in tubular and glomerular injuries are associated with mitochondrial dysfunction [8,9].
In this review, we highlight insights from studies on the interaction between mitochondria and kidney IR injury, elucidating the intricate pathophysiology of the disease and therapeutic strategies. Specifically, we discuss the detrimental roles of mitochondrial dynamics (fusion-fission), mitophagy, and metabolic rewiring, along with the effects of mitochondrial ROS on kidney IR injury.
Molecular mechanisms of kidney ischemia-reperfusion injury: mitochondrial perspective
Kidney IR injury occurs when there is a temporary reduction in blood supply to the kidneys, followed by restoration. It can be triggered by surgical procedures, trauma, or shock. The interruption of blood flow can lead to oxygen and nutrient deprivation, resulting in cellular stress. When the blood flow is restored, a burst of ROS is triggered, which exacerbates oxidative stress and inflammation. Eventually, this sequence of events can damage kidney tubular and endothelial cells, contributing to the development of AKI. Under these conditions, significant pathological changes in the mitochondria occur, manifesting as impaired mitochondrial energetics, excessive mitochondrial fragmentation, defective mitophagy, and ROS bursts within the mitochondria.
1. Mitochondrial dysfunction and related cell death
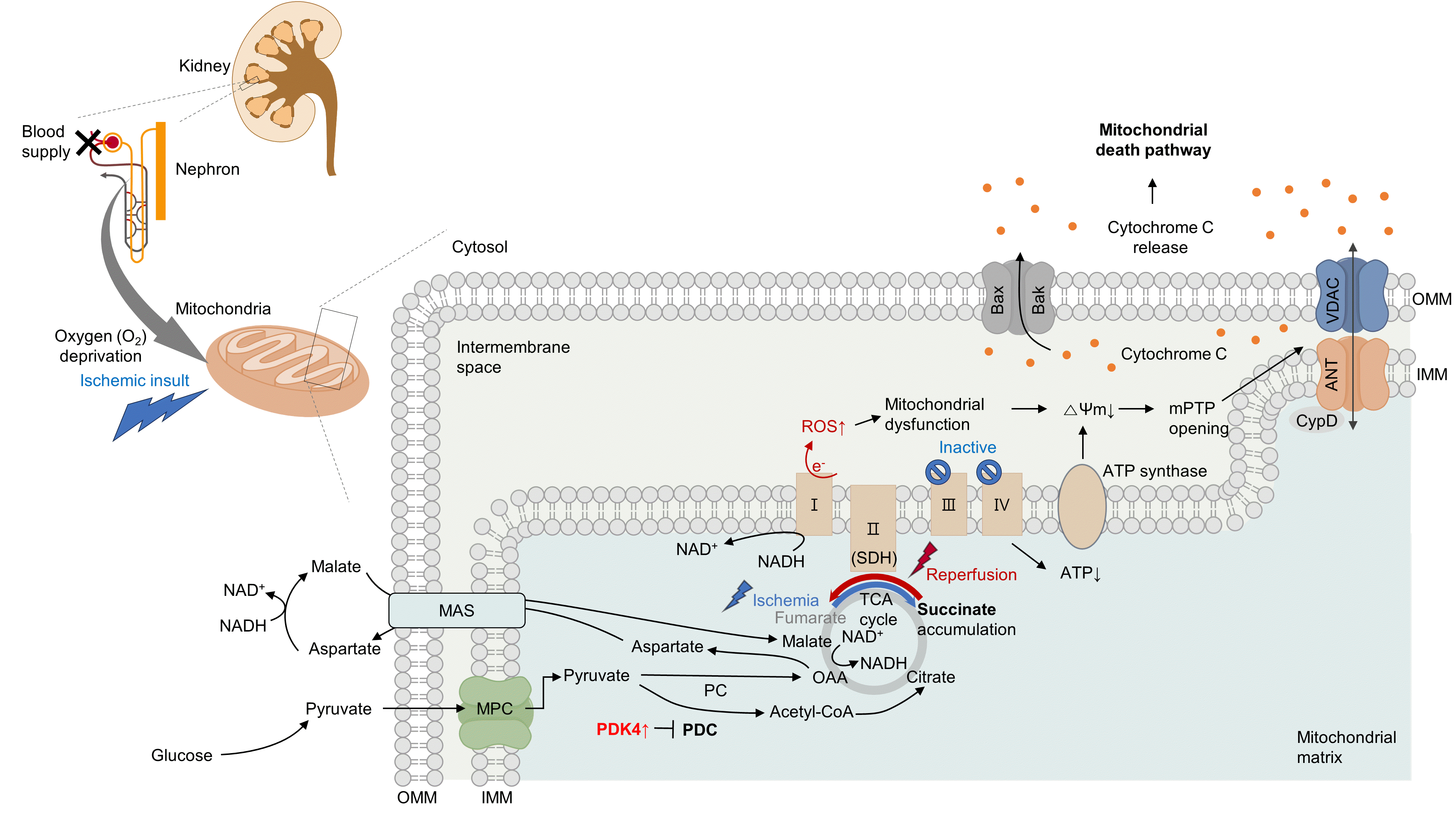
Several studies have suggested that mitochondrial energetics are compromised following an ischemic insult. This is manifested by a marked reduction in mitochondrial ATP synthesis and mitochondrial membrane potential, and subsequent opening of the mitochondrial permeability transition pore (mPTP) [10] (Fig. 1). Indeed, upon ischemia induction, ATP levels in proximal tubule cells decrease to a nadir within a few minutes [11]. Decreased ATP levels can reduce the activity of Na+/K+-ATPase, leading to the accumulation of intracellular Na+. Subsequently, this accumulation triggers Ca2+ influx and accumulation within the mitochondria. Following reperfusion, excessive increases in ROS and Ca2+ levels occur within the mitochondria, which can induce mitochondrial dysfunction [12,13]. This triggers cytochrome c release through the mPTP, initiating a caspase cascade and activating the mitochondrial death pathway [14,15]. Cyclophilin D plays a key role in the regulation of mPTP. Cyclophilin D knock-out (KO) mice are resistant to IR injury [14]. The endoplasmic reticulum (ER) and mitochondria share close physical and functional connections. Downregulation of X-box binding protein 1 (XBP1), an ER stress response protein, protects the kidneys from IR injury. Mechanistically, 3-hydroxy-3-methylglutaryl-coenzyme A reductase degradation 1 (HRD1), a protein downstream of XBP1, serves as an E3-ligase and facilitates the downregulation of nuclear factor erythroid 2-related factor 2 (NRF2) through the ubiquitination-degradation pathway [16]. XBP1 can also upregulate transcription from the NLR family pyrin domain containing 3 (NLRP3) promoter, highlighting the role of the XBP1-NLRP3 axis in regulating ER-mitochondrial crosstalk in kidneys under IR stress [17].
Under oxygen-deprived conditions, the activity of complex V is impaired, leading to defects in oxidative phosphorylation. In the IR milieu, excessive ROS production is always accompanied by a reduction in ATP production in the mitochondria [18,19]. However, whether mitochondrial dysfunction is a cause or consequence of excessive mitochondrial ROS production remains unclear. In conclusion, therapeutic strategies targeting kidney IR injury should include the prevention of mitochondria-related cell death.
2. Metabolic rewiring and aberrant succinate accumulation within the mitochondria
Previous studies have examined the role of metabolic rewiring in kidney IR injury [20-22]. During both the early and late stages of reperfusion following ischemia, kidney lactate, and pyruvate levels increase, along with an increase in hexokinase activity, indicative of enhanced glycolysis. Additionally, tubules in the process of normal regeneration and those undergoing atrophy show elevated expression of glycolytic enzymes and inhibitory phosphorylation of mitochondrial pyruvate dehydrogenase (PDH) [22]. Similarly, the progression of IR-induced AKI in human kidney proximal tubular cells (HK-2 cells) is characterized by the disruption of amino acid, nucleotide, and tricarboxylic acid cycle metabolism, and notably, a metabolic shift from fatty acid oxidation (FAO) to glycolytic conversion [20]. It could be hypothesized that the induction of a glycolytic phenotype can serve as an adaptive mechanism following tubular injury, as glycolysis can facilitate biomass synthesis crucial for cellular repair and proliferation. However, the continued presence of this phenotype seems to correlate with an elevated level of fibrosis [21].
Moreover, in proximal tubular cells, hypoxia can induce the reversal of complex II (succinate dehydrogenase, SDH) activity, resulting in excessive succinate accumulation [8,9]. Specifically, the primary source of succinate is the overflow of fumarate from purine nucleotides and partially from the reversal of the malate/aspartate shuttle [8]. Supporting this idea, our group recently discovered that inhibition of the malate/aspartate shuttle by activating PDH flux can attenuate succinate accumulation [9]. In the heart, however, succinate accumulates via the canonical Krebs cycle and not through reverse SDH activity [23]. Although the reason behind this discrepancy remains elusive (e.g., whether it is organ-specific), a reduction in either SDH activity or succinate accumulation is sufficient to mitigate damage to both the kidney and heart [8,9,23]. Consistent with this finding, a recent study revealed that fumarate can accept electrons through net reversal of the SDH complex [24].
Following reperfusion, the accumulated succinate is rapidly reoxidized by SDH. Aberrant electron accumulation during the ischemia period spills over not only to complexes III, IX, and V but also to complex I during the reperfusion period. This phenomenon, known as reverse electron transport, results in a burst of ROS leading to cell death. Although strategies to reduce tubular damage are still being established, recent findings suggest that pharmacological targeting of SDH by administering dimethyl malonate during reperfusion can ameliorate kidney IR injury by blocking rapid oxidation by SDH [25]. Therefore, the optimal timing for SDH inhibition to prevent kidney IR injury requires further investigation (Fig. 1).
3. Excessive mitochondrial fragmentation with defective mitophagy
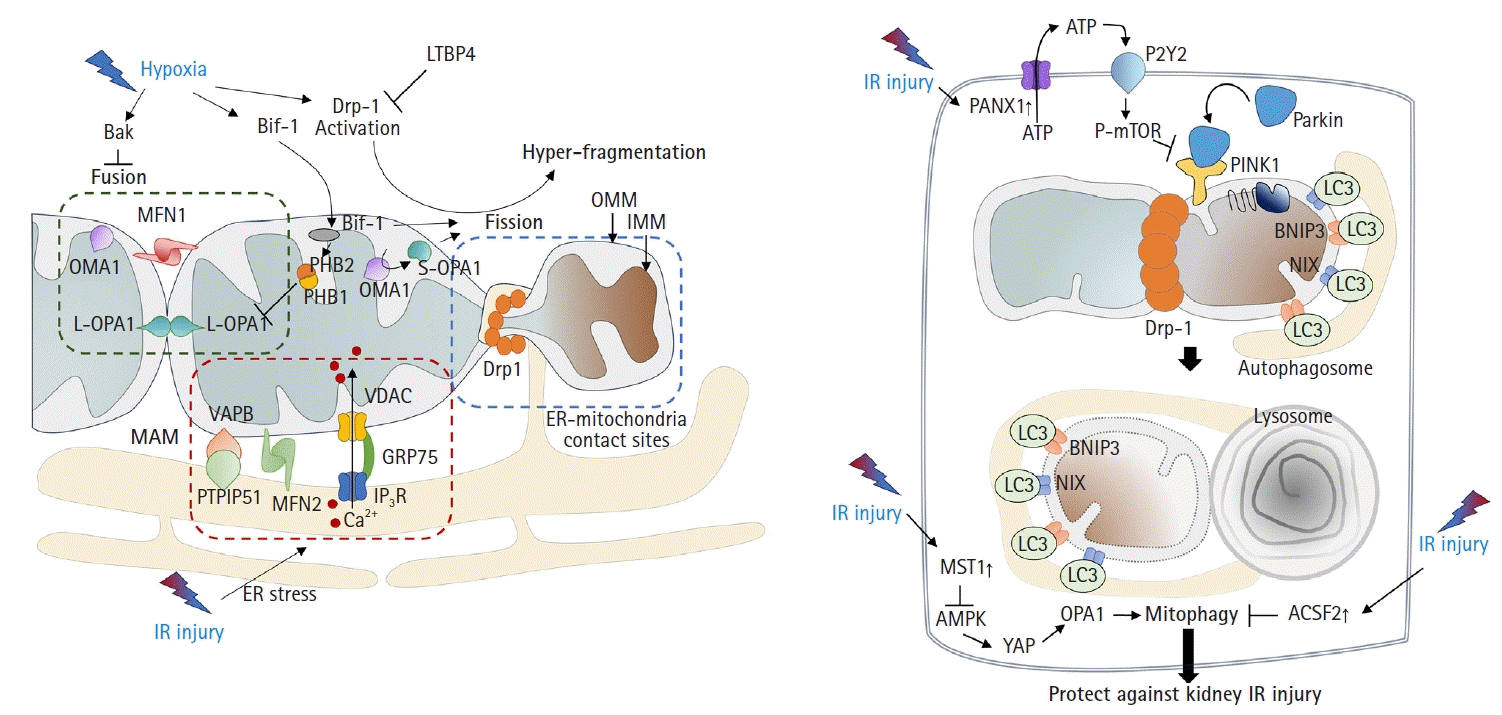
Mitochondria undergo fusion or division in response to various stimuli and stresses [26]. Environmental stresses, including hypoxia, can lead to mitochondrial hyperfragmentation. Previous studies have shown that mitochondria in proximal tubular cells undergo excessive fission [9,27]. Mechanistically, the activation of dynamin-related protein 1 (Drp1)-mediated fission and inhibition of Bak-mediated fusion can promote cleavage of the outer membrane [6,28]. In addition, recent findings have indicated that OMA1 zinc metallopeptidase-mediated proteolysis of optic atrophy 1 (OPA1), a key inner membrane fusion protein, contributes to inner membrane cleavage during cellular stress [29]. Tubular cell apoptosis and AKI can be attenuated by the expression of dominant-negative Drp1 or administration of mitochondrial division inhibitor 1 (mdivi-1), a Drp1 inhibitor [6] (Fig. 2A).
Consistent with these findings, OMA1 deficiency prevented ischemic AKI by inhibiting mitochondrial fragmentation [27]. In addition, deficiency of Numb, a multifunctional adaptor protein, can increase AKI severity by exacerbating mitochondrial fragmentation through phosphorylation of Drp1 at Ser637 [30]. These findings are supported by other studies. Under stress conditions such as IR injury, Bax-interacting factor 1 (Bif-1, a protein implicated in apoptosis and mitophagy) can regulate the mitochondrial inner membrane by interacting with prohibitin 2, which forms complexes in the inner membrane with prohibitin 1. This interaction, in tandem with prohibitin 1, can disrupt prohibitin complexes, leading to the proteolysis and inactivation of OPA1 [31]. A more recent study revealed that mitochondrial fragmentation can be induced and inhibited by uncoupling protein 2 (UCP2) gain-of-function and loss-of-function mutations, respectively, which correlated with kidney function in mice subjected to kidney IR injury [32]. Latent transforming growth factor-beta binding protein 4 (LTBP4) is upregulated in the kidney tissues of patients with AKI. LTBP4 also regulates transforming growth factor beta activity. Knockdown of LTBP4 aggravates injury by accelerating Drp1-dependent mitochondrial division, injury that can be ameliorated by mdivi-1 treatment [33].
Mitophagy, defined as the clearance of damaged mitochondria from cells to maintain a healthy mitochondrial population, is required to overcome the pathology of kidney IR injury. In particular, the mitochondrial kinase PTEN-induced kinase 1 (PINK1) and the E3-ubiquitin ligase parkin can serve as sensors of mitochondrial quality. They are activated following membrane depolarization [34]. PINK1 accumulates on defective mitochondria, and its subsequent homodimerization on the outer mitochondrial membrane facilitates the recruitment of parkin from the cytosol, mediating the clearance of damaged mitochondria [35,36]. Once recruited to depolarized mitochondria, parkin-dependent ubiquitination and proteasomal degradation of outer membrane proteins ultimately leads to mitophagy [37] (Fig. 2B).
Recent studies have shown that activation of mitophagy is essential for mitigating IR injury. Mitophagy is induced in kidney proximal tubular cells in both in vitro and in vivo models of ischemic AKI [38], and genetic disruption of mitophagy exacerbates the injury [38,39]. Another study demonstrated that Beclin-1 peptide pretreatment, which induces both autophagy and mitophagy, can protect mice against kidney IR injury [40]. Consistent with this idea, a recent study showed that the genetic deletion of pannexin 1 can prevent kidney tubular cell death, oxidative stress, and mitochondrial damage after IR injury. This protection was attributed to enhanced mitophagy, which modulates the ATP-P2Y-mammalian target of rapamycin signaling pathway [41]. The depletion of STE20-like kinase 1 can ameliorate kidney IR injury via AMP-activated protein kinase (AMPK)-yes-associated protein (YAP)-OPA1-dependent activation of mitophagy [42]. Furthermore, inhibition of acyl-CoA synthetase family member 2 (ACSF2), which is highly expressed in the mitochondria of kidney tubular cells, protects against kidney IR injury by activating mitophagy in proximal tubular cells [43]. Bcl-2 interacting protein 3 (BNIP3), a noncanonical regulator of mitophagy, has also been implicated in kidney IR injury [44]. Taken together, these findings suggest that the regulation of mitophagy is an important therapeutic strategy against kidney IR injury.
4. Ferroptosis
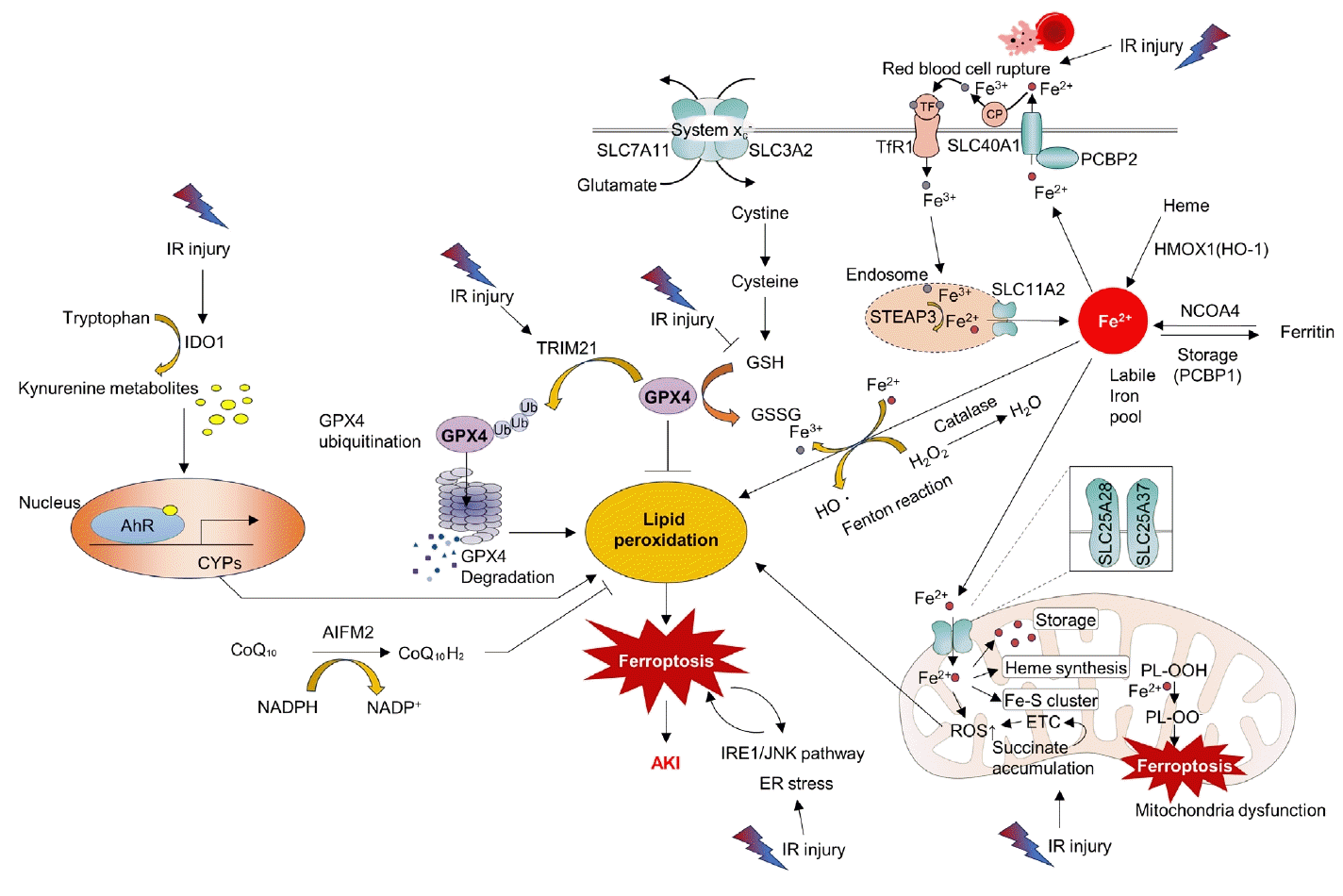
Ferroptosis is a relatively newly discovered form of cell death [45]. It is an iron-dependent cell death process characterized by the accumulation of lipid peroxides. This process is distinct from that of apoptosis and necrosis [45]. Mitochondria in ferroptotic cells display pathological changes in morphology such as shrinkage and loss of cristae [46]. As suggested by its name, ferroptosis can be triggered by various stimuli that lead to iron overload. A pathological increase in the cellular iron content can accelerate the Fenton reaction, resulting in the production of hydroxyl radicals (•OH) and other ROS. Additionally, excess polyunsaturated fatty acids, deprivation of glutathione, and reduced function of mitochondrial glutathione peroxidase 4 (GPX4), an enzyme that decreases lipid peroxides, can induce ferroptosis by increasing lipid ROS (Fig. 3) [46].
Accumulating evidence has shown that ferroptosis is an important molecular target for the treatment of kidney IR injury. ROS bursts during reperfusion injuries are associated with ferroptosis [47]. Researchers have found that the aryl hydrocarbon receptor (AhR) is activated during reoxygenation, which leads to ROS production, lipid peroxidation, and ferroptotic cell death [47]. The same researchers have also shown that AhR-mediated ferroptosis is detrimental not only during the reperfusion phase but also during the ischemic phase [48]. Tripartite motif-containing protein 21 (TRIM21) is upregulated in the kidney during IR injury and promotes ferroptosis by ubiquitinating and degrading GPX4. TRIM21 inhibition may be a strategy to reduce IR injury [49]. Inhibition of the ER stress inositol-requiring enzyme 1 (IRE1)/ c-Jun N-terminal kinase (JNK) pathway in kidney tubular epithelial cells (TECs) attenuates ferroptosis in AKI. Intriguingly, inhibition of ferroptosis can also attenuate IRE1/JNK signaling, suggesting a feed-forward loop [50]. Moreover, preserving heme oxygenase 1 (HO-1) expression by inhibiting miR-3587 can attenuate ferroptosis and kidney IR injury, suggesting that ferroptosis is a key detrimental pathological mechanism of IR injury [51].
Therapeutic strategies of kidney ischemia-reperfusion injury targeting mitochondria
1. Targeting mitochondrial function including reactive oxygen species, biogenesis, and related mitochondrial death pathway
Several studies have suggested that enhancing mitochondrial quantity or function can be beneficial for retarding or preventing IR injury. Approaches such as mitochondrial gene delivery [52], coenzyme Q10 administration [53], and mitochondrial transplantation [54,55] have been shown to be able to attenuate kidney injury. Supporting these findings, the overexpression of reduced nicotinamide adenine dinucleotide (NADH):ubiquinone oxidoreductase core subunit V1 (NDUFV1), which encodes a 51-kDa subunit of complex I, can attenuate kidney IR injury by improving mitochondrial function [56].
Recent studies have revealed that a deficiency in oxidized nicotinamide adenine dinucleotide (NAD+) can contribute to mitochondrial dysfunction, leading to inflammation and kidney disease progression. NAD+ supplementation prevents inflammation during kidney injury. This protective mechanism is attributed to the prevention of mitochondrial RNA leakage into the cytosol and inhibition of cytosolic pattern recognition receptor retinoic acid-inducible gene I (RIG-I), both of which are ameliorated by restoring NAD+ levels [57]. Moreover, it has been shown that NAD+ supplementation can enhance mitochondrial biogenesis in a sirtuin (SIRT)-dependent manner and that its precursor, nicotinamide riboside, can attenuate IR injury [58]. Mitoquinone, a mitochondria-targeted antioxidant, alleviates IR injury by activating the SIRT3-dependent pathway [59]. These findings are supported by a recent study showing that SIRT3 deficiency can dampen early-stage fibrosis after IR injury [60]. The pregnane X receptor (PXR)/aldo-keto reductase family 1, member B7 (AKR1B7) axis was recently introduced as a novel therapeutic target [61]. Another recent study highlighted that peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), a master regulator of mitochondrial biogenesis, is downregulated in IR injury [62]. That study further demonstrated that inhibiting forkhead box protein O1 (FOXO1) could restore PGC-1α transcription, implicating the FOXO1 inhibitor AS1842856 as a potential therapeutic agent for IR injury [62].
Certain drugs have demonstrated promising results. Lasmiditan, a 5-hydroxytryptamine receptor 1F (5-HT1F) agonist, attenuates IR injury by enhancing mitochondrial biogenesis [63]. The importance of 5-HT1F has also been highlighted in a genetic KO model [64]. The antidiabetic drug saxagliptin attenuates IR injury by activating the NRF2/HO-1 pathway [65]. Eplerenone, a mineralocorticoid antagonist available in clinical practice, can reduce kidney IR injury by modulating inflammation and the SIRT1/SIRT3/PGC-1α pathway [66]. Mefunidone, a novel pyridinone drug, can regulate the mitochondria-related Bax/Bcl-2/cleaved-caspase 3 apoptotic pathway, protect mitochondrial electron transport chain complexes III and V levels, and ameliorate kidney function in both in vivo and in vitro disease models [67]. Additionally, acetate, a well-known short-chain fatty acid, reduces cellular ROS production and the cells positive for specific indicator for mitochondrial superoxide (MitoSox, Thermo Fisher Scientific, Waltham, MA, USA) that are indicative of mitochondrial ROS production. Acetate also reduced mitochondrial fission and alleviated IR injury in a murine model [68]. Notably, the ROS-responsive chitosan-SS31 prodrug, which has antioxidant properties, alleviated ROS levels and improved kidney function in a murine model of IR injury [69]. As a member of the vitamin E family, γ-tocotrienol improved mitochondrial function, promoted tubular regeneration, and ameliorated kidney functions in a murine IR model [70]. Treprostinil, a U.S. Food and Drug Administration-approved prostacyclin (PGI2) analog, can improve mitochondrial function and reduce IR injury by a PGC-1α- and SIRT-dependent mechanism [71].
The aforementioned mPTP opening resulting from mitochondrial dysfunction [10] is also gaining attention as a molecular target for IR injury. A recent study highlighted that mitochondrial ribosomal protein L7/L12 (MRPL12) can bind to adenosine nucleotide translocase 3 (ANT3), thereby stabilizing the mPTP. However, during AKI, MRPL12 expression is reduced in TECs, leading to ANT3 conformational changes and mPTP opening. Overexpression of MRPL12 can protect TECs from apoptosis during hypoxia/reoxygenation challenge [72]. Similarly, Bax inhibitor-1, which conveys anti-apoptotic signals to the mitochondria, is implicated in both IR injury and mitochondrial health [73]. Glutamine administration attenuated kidney damage in vivo during AKI and restored TEC viability in vitro by affecting glutamine gamma glutamyltransferase 2 (Tgm2) and apoptosis signal-regulating kinase (Ask1) [74].
2. Targeting mitochondrial dynamics and mitophagy
Excessive mitochondrial fragmentation has been observed in kidney IR injury [6,9]. There is growing interest in whether strategies to reverse mitochondrial fragmentation can mitigate kidney injury. Recently, it was revealed that the catalytic subunit of DNA-dependent protein kinase (DNA-PKcs), known for its functional relationship with cancer and aging, can directly phosphorylate mitochondrial fission 1 protein (Fis1) to induce mitochondrial fission and fragmentation in TECs upon IR. The authors showed that knock-in mice expressing a nonphosphorylatable mutant exhibited improved kidney function, improved histological features, and reduced mitochondrial fragmentation upon AKI induction [75]. Furthermore, SIRT3 promotes mitofusin 2 ubiquitination and degradation, thus suppressing IR-induced AKI [76]. Empagliflozin, an inhibitor of sodium-glucose cotransporter 2, enhanced mitochondrial fusion by promoting the AMPK-OPA1 pathway in an IR model [77]. Emodin (1,3,8-trihydroxy-6-methylanthraquinone), an anthraquinone derivative, demonstrated protective effects in IR-injured mice by suppressing calcium/calmodulin-dependent protein kinase II (CAMKII)/Drp1-mediated mitochondrial fission [78]. Additionally, SIRT3 has been found to alleviate kidney IR injury by enhancing mitochondrial fusion and activating the OPA1 signaling pathway [79].
Regarding mitophagy, overexpression of BNIP3 is sufficient to overcome kidney IR injury [44,80]. Mesenchymal stem cell-derived extracellular vesicles containing miR-223-3p can activate mitophagy and improve kidney IR injury by inhibiting NLRP3 [81]. Collectively, these findings highlight the potential of mitophagy as a viable target for the treatment of IR injury. Further studies are required to validate the role of small molecules and other compounds that target mitophagy.
3. Targeting metabolic reprogramming
Succinate accumulation during ischemia leads to excessive ROS generation and has been identified as a critical factor in kidney IR injury [82]. Recently, in silico analysis revealed that mitochondrial FAO, peroxisomal lipid metabolism, fatty acid metabolism, and glycolysis are downregulated, whereas the pentose phosphate pathway is upregulated in ischemic kidney tissues [83]. However, the causal relationship between this metabolic reprogramming and its effects requires further investigation. As previously mentioned, PDH activation can diminish succinate accumulation, partly through the restoration of mitochondrial function and attenuation of the malate-aspartate shuttle [9]. Enhanced FAO mediated by carnitine palmitoyltransferase 1A (CPT1A) and delivery of hypoxic mesenchymal stem cell-derived extracellular vesicles can attenuate IR injury, suggesting that metabolic reprogramming is a possible treatment for IR injury [84]. Moreover, AKT serine/threonine kinase 1 (Akt1) and protein kinase B, the main downstream molecules of the insulin-phosphoinositide 3-kinase (PI3K) signaling pathway, are activated during IR injury, dampening tubular apoptosis and kidney injury, which can eventually lead to kidney fibrosis [85,86]. Given that excessive insulin-PI3K-Akt is indicative of insulin resistance, further studies are needed to examine whether improving metabolic conditions, such as insulin sensitization, could ameliorate kidney IR injury.
4. Targeting ferroptosis and inflammation
A recent review provided a comprehensive overview of the role of ferroptosis in kidney IR injury after kidney transplantation [87]. Hence, this section briefly discusses recent therapeutic approaches targeting ferroptosis. Visomitin, a novel mitochondria-targeting antioxidant, can mitigate mitochondrial ROS production, resulting in decreased levels of lipid peroxidation and ferroptosis, thereby protecting against ischemia- or nephrotoxicity-induced AKI [88]. Irisin, an exercise-induced hormone known to improve mitochondrial function and reduce ROS production [89], has been shown to protect against IR injury by upregulating GPX4 in vivo. Importantly, the protective effect of irisin was abrogated by the inhibition of GPX4 [90]. This is supported by an important observation that the ferroptosis suppressor liproxstatin-1 can reduce kidney IR injury and decrease the mortality rate in Gpx4-deficient mice [91].
Another facet of the mitochondrial contribution to the pathogenesis of IR injury is its relationship with inflammation [92]. Pathogen-associated molecular patterns and damage-associated molecular patterns are important inducers of NLRP3 inflammasome formation in immune cells. Because mitochondrial ROS are among the most important inducers of the NLRP3 inflammasome [92], targeting this process is crucial for addressing IR injury [93]. Remdesivir, a well-known antiviral agent approved for treating coronavirus disease 2019, has been shown to alleviate AKI through NLRP3 inflammasome inhibition [94]. Similarly, the dopamine D1 receptor agonist A68930 can attenuate AKI by inhibiting NLRP3 inflammasome activation [95]. Further research is warranted to understand this novel compound, which targets inflammatory immune cells in addition to macrophages and monocytes, and to develop novel compounds targeting the NLRP3 inflammasome.
Conclusion
This review is a comprehensive overview of the interplay between the mitochondria and acute kidney IR injury and provides important insights gleaned from previous studies. Several key points have emerged.
During acute kidney ischemia, the mitochondria exhibit reduced ATP production and aberrant mitochondrial dynamics, leading to energy depletion, which can adversely affect the survival and function of kidney cells. During the reperfusion period, succinate accumulated during ischemia is rapidly oxidized, releasing electrons toward electron complex I in a process known as reverse electron transport. This abnormal phenomenon results in a ROS burst, triggering the mitochondrial death pathway. Various strategies have been explored to protect the mitochondria, including the use of antioxidants, mitochondria-specific drugs, and interventions to improve mitochondrial metabolism and reduce succinate accumulation.
Furthermore, mitochondrial damage can trigger inflammatory responses, exacerbating AKI severity. Additionally, mitochondrial dysfunction and the depletion of mitochondrial GPX4 accompany ferroptosis, which occurs during IR injury. Thus, mitochondria also play crucial roles in the regulation of cell death and injury-related inflammation.
In conclusion, the role of mitochondria in acute kidney IR injury raises several unresolved questions. Therefore, further studies are warranted. The development of novel strategies to effectively safeguard mitochondria and prevent or treat AKI is of paramount importance. Such investigations hold promise for pioneering new directions for the prevention and treatment of acute kidney IR injury.
 XML Download
XML Download