

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Myocardial hypertrophy (MH) is an increase in the volume and mass of the heart as it adapts to various intrinsic and extrinsic stimuli.1) Massive evidence has demonstrated that multiple cardiovascular diseases, such as myocardial infarction, hypertension, are associated with pathological MH.2) Long-term pathological MH contributes to arrhythmias and heart failure, which seriously pose a serious life-threatening risk to patients.3) At present, it is essential to search for and find new effective therapeutic drugs to inhibit MH.
Dronedarone is a multichannel blocker that can simultaneously inhibit Na, K and Ca ion channels, and also has β-receptor antagonism.4) Therefore, dronedarone is an antiarrhythmic drug indicated for patients with paroxysmal or persistent atrial fibrillation (AF). At present, a previous study indicated that dronedarone suppressed myocardial remodeling,5) suggesting that dronedarone might be an effective medicine for the treatment of MH. Therefore, in this study, we will investigate the effects of dronedarone on MH and understand the underlying molecular mechanism.
Sirtuin 1 (SIRT1) is a well-known protein deacetylase that mediates a variety of biological processes by regulating the acetylation of target proteins.6) A growing body of researches have supported that SIRT1 was closely associated with the regulation of MH.7)8) Li and colleagues proposed that fibroblast growth factor 21 through upregulating SIRT1 expression, could alleviate angiotensin II (Ang II)-induced MH.9) Of note, SIRT1 could mediate the deacetylation of forkhead box O3 (FOXO3) to promote FOXO3 expression, thereby regulating various diseases such as hepatocellular carcinoma, chronic obstructive pulmonary disease, premature senescence.10)11)12) Additionally, FOXO3 has been reported to be implicated in MH.13) Based on these evidences, it remains to be determined whether dronedarone targets the SIRT1/FOXO3 axis to affect the progression of MH.
As previously described, protein kinase A (PKA), a multi-subunit cellular kinase, plays a crucial role in a variety of physiological and pathological conditions. There is increasing evidence that PKA signaling was closely associated with cardiac dysfunction including MH.14) For instance, protein kinase inhibitor alpha (PKI, also known as PKIA) peptide suppressed PKA activity to decrease myocyte apoptosis of β-adrenergic receptor (β-AR) agonist-induced adult feline ventricular myocytes.15) PKA inhibitor (KT-5720) could attenuate urotensin II-induced hypertrophy of neonatal rat cardiomyocytes, indicating that inhibition of PKA signaling might affect MH.16) PKIA is encoded for PKIα, which belongs to PKI family members that interact with PKA and suppress its kinase activity, thereby regulating PKA-mediated signaling events.17) A study published in 2020 suggested that angiotensin-(1-9) inhibited the expression of PKIA and activated the PKA pathway to participate in MH.18) In this study, we will further determine the role of PKIA in MH. FOXO3 is a well-known transcription factor that could interact with the promoter region of targeted genes to regulate gene expression.19) JASPAR website predicted a binding site between FOXO3 and PKIA. Therefore, the downstream mechanism of FOXO3 might target PKIA to mediate the PKA signaling pathway, further affecting the progression of MH.
Based on the above backgrounds, we hypothesized that dronedarone suppresses Ang II-induced MH through mediating SIRT1/FOXO3/PKIA/PKA pathway. Here, H9C2 cells were treated with Ang II to establish an in vitro cell model of MH and rats were subjected to transverse aortic constriction (TAC) surgery to establish an animal model of MH. A series of experiments were conducted to testify our hypothesis. We hope that our findings will provide an experimental basis supporting the use of dronedarone in MH.
METHODS
Ethical statement
All animal experiments were permitted by The First Affiliated Hospital, Hengyang Medical School, University of South China (approval No. 2022ll0628003).
Cell culture
H9C2 cells were bought from ATCC (Rockville, MD, USA). All cells were maintained in Dulbecco’s Modified Eagle medium (Thermo Fisher Scientific, Waltham, MA, USA) with 10% fetal bovine serum (Thermo Fisher Scientific) and 1% antibiotics (Beyotime, Shanghai, China) under the condition of 5% CO2 and 37°C.
H9C2 cells were treated with 50, 100, 150 nmol/L Ang II (BIOFOUNT, Beijing, China) for 48 hours to establish a cell model of MH.
In addition, different concentrations of dronedarone (1, 5, 10 μM) (Selleck, Shanghai, China) were used to treat Ang II-induced H9C2 cells for 6 hours.
Cell transfection
The short hairpin RNA targeting SIRT1 (sh-SIRT1), overexpression vector of SIRT1(oe-SIRT1) or FOXO3 (oe-FOXO3) or PKIA (oe-PKIA) as well as their negative control groups (sh-NC, oe-NC) were obtained from GenePharma (Shanghai, China). H9C2 cells were seeded into 6-well plates and incubated overnight. The cells then transfected with the above sequences and plasmids for 48 h using Lipofectamine™ 3000 (Invitrogen, Carlsbad CA, USA) following the instructions.
Reverse transcription quantitative polymerase chain reaction
Total RNA was acquired from H9C2 cells with the indicated treatments using TRIzol reagent (Beyotime). After cDNA synthesis was performed using Script Reverse Transcription Reagent Kit (TaKaRa, Beijing, China), SYBR Premix Ex Taq II Kit (TaKaRa) was used for the quantitative polymerase chain reaction (qPCR) process (Table 1).
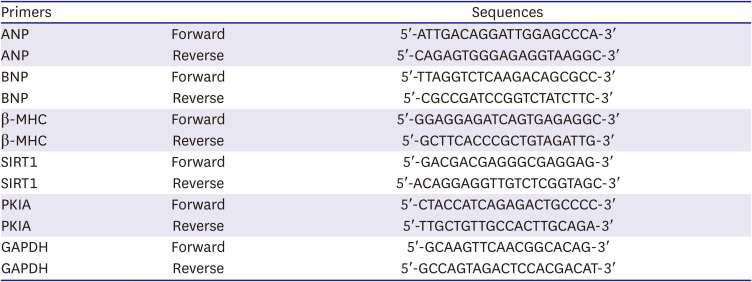
Table 1
Primer sequences
ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; β-MHC = β-myosin heavy chain; SIRT1 = sirtuin 1; PKIA = protein kinase inhibitor alpha; GAPDH = glyceraldehyde 3-phosphate dehydrogenase.
![]()
All data were calculated by using 2−ΔΔCt formula. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) served as reference gene.
Western blot
Total protein was extracted from with H9C2 cells and myocardial tissues from rats using RIPA buffer (Beyotime). After quantification of protein concentration using BCA protein kit (Beyotime), the proteins were separated by sodium dodecyl-sulfate polyacrylamide gel electrophoresis, and then were transferred onto the polyvinylidene difluoride (PVDF) membrane. After blocking with 5% bovine serum albumin for 1 hour, the PVDF membrane was immersed in primary antibodies including SIRT1 (#ab110304; Abcam, Cambridge, UK), atrial natriuretic peptide (ANP, #PA5-72527; Thermo Fisher Scientific), B type natriuretic peptide (BNP, #PA5-96084; Thermo Fisher Scientific), β-myosin heavy chain (β-MHC, #ab37484; Abcam), FOXO3 (#PA5-27145; Thermo Fisher Scientific), Ac-FOXO3 (#9441S; Cell Signaling Technology, Danvers, MA, USA), PKIA (#PAB17373; AmyJet Scientific Inc, Wuhan, China), PKA (#PA5-17626; Thermo Fisher Scientific), tubulin (#ab18207; Abcam), and β-actin (#ab8226; Abcam) overnight at 4°C. Subsequently, horseradish peroxidase-conjugated secondary antibody was then applied to the PVDF membrane for 1 hour. ECL chemiluminescent reagent (Beyotime) was employed to detect protein bands. The densitometry analysis was estimated by Image J (National Institutes of Health, Bethesda, MD, USA).
Determination of cell surface area
H9C2 cells with the indicated treatments were fixed with 10% formaldehyde. The fixed cells were then stained with 1% crystal violet solution. The samples were photographed with a microscope (Nikon Co., Tokyo, Japan) and the size of cells was determined using ImageJ software.
H9C2 cells with the indicated treatments were fixed the cells with 4% paraformaldehyde and permeabilized with 0.5% Triton X-100. Then, rhodamine phalloidin staining (Invitrogen) was used to stain the actin filaments. Besides, the nucleus was stained with 4′,6-diamidino-2-phenylindole (Abcam, Waltham, MA, USA). Cells were imaged under a fluorescence microscopy (Nikon Co.).
Luciferase reporter assay
The JASPAR database (https://jaspar.genereg.net/) was employed to predict the binding site between FOXO3 and PKIA promoter. The wild-type sequences of PKIA promoter were inserted into pGL3 plasmid (GenePharma) to obtain WT-PKIA plasmid and mutated sequences of PKIA promoter were inserted into pGL3 plasmid (GenePharma) to obtain MUT-PKIA plasmid. The plasmids of PKIA promoter were co-transfected with oe-FOXO3 and the luciferase activity was tested using Dual-Glo Luciferase Assay System (Promega, Fitchburg, WI, USA) according to the instruction.
Transverse aortic constriction surgery for rats
The pressure overload-induced MH model in rats was established via TAC surgery. The TAC surgery of rats was conducted following a previous study.20) Fifteen Sprague-Dawley rats, aged 8 weeks and weighing 190–220 g, were purchased from Hunan Silaikejingda Experimental Animal Co., Ltd (Changsha, China). Rats were randomly divided into three groups: Sham, TAC, TAC+dronedarone. All operations were conducted under sterile conditions and anesthetized by intraperitoneal injection of pentobarbital sodium. Under sterile conditions, the 2nd–3rd intercostal space of the left lateral breast was incised to expose the ascending aorta. After exposing the aortic arch, the aortic arch was banded against 26-G needle with 7-0 silk suture. The needle was removed before abdominal closure. The procedures of the sham group were the same as those of the model group, except that the thoracic aorta was not ligated. After TAC surgery, rats in the TAC+dronedarone group received 90 mg/kg of dronedarone daily by intragastric gavage for 8 weeks. The others were given saline solution. Finally, all rats were killed, and corresponding samples were taken for follow-up experiments.
Hematoxylin and eosin staining
After fixation with 4% paraformaldehyde solution and embeddedness with paraffin, myocardial tissues of rats were cut into 5 µm thick serial sections. The sections were stained with hematoxylin and eosin (H&E) (Beyotime). A fluorescence microscopy took pictures for the stained sections.
Echocardiographic assessment of cardiac function
After rats were anesthetized with 1–2% isoflurane, the cardiac function was examined using transthoracic ultrasonography with a VisualSonics 2100 echocardiograph (FUJIFILM VisualSonics, Toronto, ON, Canada). Images of the parasternal long axis and short axis of the left ventricle were acquired using a 30-MHz transducer. The ejection fraction (EF)% and fraction shortening (FS)% were determined and recorded. Then the above parameters were calculated with Vevo Lab 3.1.0 software (FUJIFILM VisualSonics).
Statistical analysis
Data were presented as mean ± standard deviation. GraphPad Prism 9.0 was employed to analyze statistical differences in the data. Student’s t-test was used for comparison between two groups and one-way analysis of variance was used for comparison among multiple groups. All experiments were repeated three times independently. The p value <0.05 was considered as significant difference.
RESULTS
Angiotensin II stimulation successfully established a cell model of myocardial hypertrophy and inhibited sirtuin 1 expression in H9C2 cells
To establish a cell model of MH, different concentrations of Ang II (50, 100, 150 nmol/L) were applied to stimulate H9C2 cells for 48 hours. MTT assay showed that cell viability was gradually decreased along with increasing concentrations of Ang II (Figure 1A). Crystal violet staining and rhodamine phalloidin staining exhibited that cell size was expanded gradually with the increasing Ang II concentrations (Figure 1B and C). In addition, both mRNA and protein levels of hypertrophy-related genes including ANP, BNP, β-MHC were enhanced in an Ang II concentration-dependent manner (Figure 1D and E). Of note, the protein level of SIRT1 was evidently decreased with Ang II induction in H9C2 cells (Figure 1E). Taken together, Ang II stimulation induced hypertrophy in H9C2 cells and resulted in abnormally low expression of SIRT1.
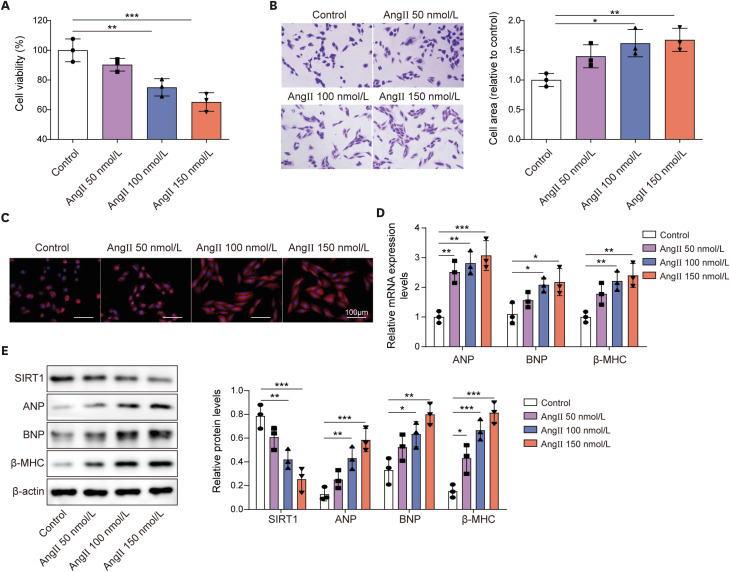
Figure 1
Ang II stimulation successfully established a cell model of MH and inhibited SIRT1 expression in H9C2 cells.
H9C2 cells were treated with diffident concentrations of Ang II (50, 100, 150 nmol/L) for 48 hours. (A) Cell viability was detected using MTT assay. (B, C) Cell size was evaluated using crystal violet staining and rhodamine phalloidin staining. (D) The mRNA expression of ANP, BNP, and β-MHC were examined by RT-qPCR. (E) The protein levels of SIRT1, ANP, BNP, and β-MHC were examined by western blot.
Ang II = angiotensin II; ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; MH = myocardial hypertrophy; RT-qPCR = reverse transcription quantitative polymerase chain reaction; SIRT1 = sirtuin 1; β-MHC = β-myosin heavy chain.
*p<0.05, **p<0.01, ***p<0.001. All data were obtained from 3 independent replications.
![]()
Dronedarone attenuated angiotensin II-induced myocardial hypertrophy
Based on the results of Figure 1, 150 nmol/L Ang II was selected to induce H9C2 cells for subsequent experiments. To explore the functions of dronedarone on Ang II-induced H9C2 cells, different concentrations of dronedarone (1, 5, 10 μM) were used to treat Ang II-induced H9C2 cells for 6 hours. Crystal violet staining and rhodamine phalloidin staining displayed that Ang II-induced expansion of cell size was shrunk with dronedarone treatment, which was in a dronedarone concentration-dependent manner (Figure 2A and B). Moreover, Ang II-induced inhibition of SIRT1 expression and elevation of hypertrophy-related proteins including ANP, BNP and β-MHC were gradually abolished by the increasing dronedarone concentration (Figure 2C). Of note, 10 μM dronedarone was used for subsequent experiments. Amiodarone and dronedarone have similar chemical structure, both are benzofuran derivatives, and both are used for antiarrhythmic therapy.21) There were studies reporting that amiodarone can alleviate MH.22)23) Here, we treated Ang II-induced H9C2 cells with different concentrations of amiodarone (1, 5, 10 μM), and western blot was used to detect the expression of SIRT1, ANP, BNP and β-MHC. It was also found that amiodarone significantly increased SIRT1 expression and decreased the expression of ANP, BNP, and β-MHC when the concentrations of amiodarone were at 5, 10 μM (Supplementary Figure 1). Taken together, we came a conclusion that benzofuran derivatives including dronedarone and amiodarone could suppress Ang II-induced MH.
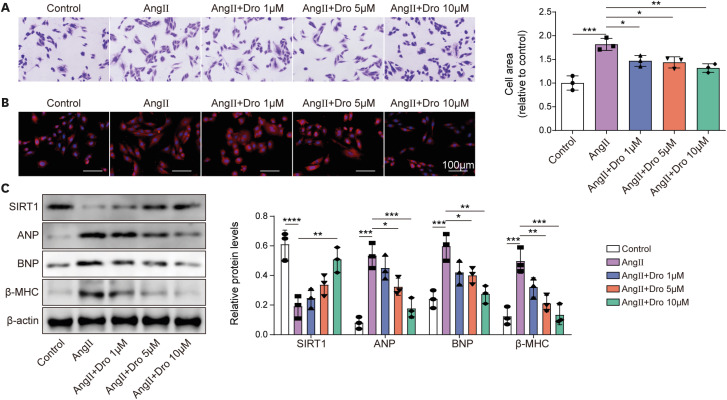
Figure 2
Dronedarone attenuated Ang II-induced MH.
Diffident concentrations of dronedarone (1, 5, 10 μM) were used to treat H9C2 cells upon 150 nmol/L Ang II induction for 6 hours. (A, B) Cell size was evaluated using crystal violet staining and rhodamine phalloidin staining. (C) The protein levels of SIRT1, ANP, BNP, and β-MHC were examined by western blot.
Ang II = angiotensin II; ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; Dro = dronedarone; MH = myocardial hypertrophy; SIRT1 = sirtuin 1; β-MHC = β-myosin heavy chain.
*p<0.05, **p<0.01, ***p<0.001. All data were obtained from 3 independent replications.
![]()
Dronedarone regulated forkhead box O3 acetylation to promote protein kinase inhibitor alpha expression through promoting sirtuin 1 expression, attenuating angiotensin II-induced myocardial hypertrophy
We desired to investigate whether SIRT1 is implicated in dronedarone-mediated suppressive effects on MH and the downstream molecule mechanism of SIRT1. Firstly, SIRT1 expression in H9C2 cells was apparently enhanced by oe-SIRT1 transfection (Figure 3A). Ang II stimulation notably inhibited the protein levels of SIRT1 and FOXO3, but enhanced acetylation of FOXO3, which was attenuated by oe-SIRT1 or dronedarone treatment. As expected, the combined effects of oe-SIRT1 and dronedarone on FOXO3 and Ac-FOXO3 proteins were stronger than the effects of oe-SIRT1or dronedarone alone (Figure 3B). Ang II-induced expansion of cell size was suppressed by SIRT1 upregulation or dronedarone. Under the combined oe-SIRT1 and dronedarone, the decrease in cell size was the most evident, as demonstrated by crystal violet staining and rhodamine phalloidin staining (Figure 3C and D). Through JASPAR prediction, we found that FOXO3 might interact with the PKIA promoter. Then, FOXO3 expression in H9C2 cells was upregulated by oe-FOXO3 transfection. Luciferase activity assay exhibited that oe-FOXO3 observably boosted the luciferase activity in WT-PKIA group, validating the interaction between FOXO3 and PKIA promoter (Figure 3E). In addition, oe-SIRT1 or dronedarone elevated PKIA level while inhibited the ANP, BNP, and β-MHC levels, which these influences were strengthened by SIRT1 upregulation in combination with dronedarone (Figure 3F). Collectively, dronedarone elevated SIRT1 expression to increase FOXO3 expression via mediating deacetylation of FOXO3 and to enhance PKIA expression, eventually alleviating Ang II-induced MH.
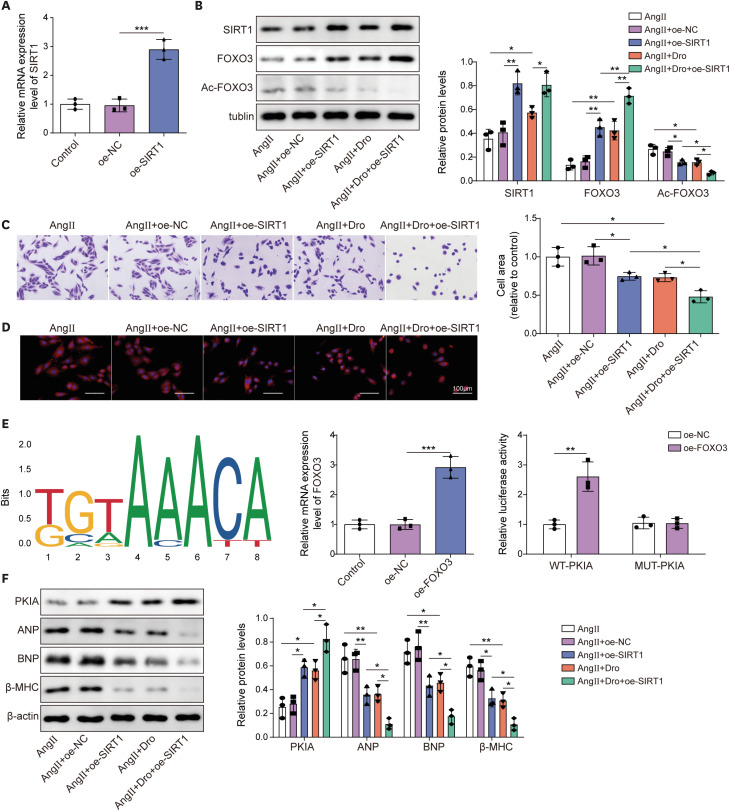
Figure 3
Dronedarone regulated FOXO3 acetylation to promote PKIA expression through promoting SIRT1 expression, attenuating Ang II-induced MH.
(A) The overexpression efficiency of SIRT1 was measured using RT-qPCR. Oe-SIRT1 H9C2 cells were subjected to Ang II induction and dronedarone treatment. (B) The protein levels of SIRT1, FOXO3 and Ac-FOXO3 were examined by western blot. (C, D) Cell size was evaluated using crystal violet staining and rhodamine phalloidin staining. (E) JASPAR predicted the binding site between FOXO3 and PKIA promoter. RT-qPCR was used to detect the overexpression efficiency of FOXO3. Luciferase activity assay validated the interaction between FOXO3 and PKIA promoter. (F) The protein levels of PKIA, ANP, BNP and β-MHC were examined by western blot.
Ang II = angiotensin II; ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; Dro = dronedarone; FOXO3 = forkhead box O3; MH = myocardial hypertrophy; NC = negative control; oe- = overexpression; PKIA = protein kinase inhibitor α; RT-qPCR = reverse transcription quantitative polymerase chain reaction; SIRT1 = sirtuin 1; β-MHC = β-myosin heavy chain.
*p<0.05, **p<0.01, ***p<0.001. All data were obtained from 3 independent replications.
![]()
Dronedarone suppressed PKA pathway to restrain angiotensin II-induced myocardial hypertrophy through upregulating protein kinase inhibitor alpha expression
To clarify whether PKIA is involved in dronedarone-mediated suppressive effects on MH, oe-PKIA was transfected into H9C2 cells to upregulate PKIA expression, which was determined by reverse transcription-qPCR (Figure 4A). Ang II-induced increase of cell size was reversed by oe-PKIA or dronedarone treatment. Notable, the combination of oe-PKIA and dronedarone treatment generated most obvious in suppressing cell size (Figure 4B and C). In addition, oe-PKIA or dronedarone treatment could enhance PKIA level whereas decrease PKA, ANP, BNP, and β-MHC levels, which were reinforced by oe-PKIA together with dronedarone treatment (Figure 4D). In total, dronedarone inactivated PKA pathway through elevating PKIA expression, thereby suppressing Ang II-induced MH.

Figure 4
Dronedarone suppressed PKA pathway to restrain Ang II-induced MH through upregulating PKIA expression.
(A) The overexpression efficiency of PKIA in H9C2 cells was measured using RT-qPCR. PKIA-overexpressed H9C2 cells were subjected to Ang II induction or together with dronedarone treatment. (B, C) Cell size was evaluated using crystal violet staining and rhodamine phalloidin staining. (D) The protein levels of PKIA, PKA, ANP, BNP, and β-MHC were examined by western blot.
Ang II = angiotensin II; ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; Dro = dronedarone; MH = myocardial hypertrophy; NC = negative control; oe- = overexpression; PKA = protein kinase A; PKIA = protein kinase inhibitor alpha; RT-qPCR = reverse transcription quantitative polymerase chain reaction; β-MHC = β-myosin heavy chain.
*p<0.05, **p<0.01, ***p<0.001. All data were obtained from 3 independent replications.
![]()
Dronedarone attenuated angiotensin II-induced myocardial hypertrophy through regulating sirtuin 1/forkhead box O3/protein kinase inhibitor alpha pathway
Here, the role of SIRT1/FOXO3/PKIA axis in dronedarone-mediated influences of Ang II-induced H9C2 cells was investigated. Firstly, SIRT1 and PKIA expression in H9C2 cells was evidently decreased by sh-SIRT1 transfection. When H9C2 cells were transfected with sh-SIRT1 and oe-PKIA simultaneously, SIRT1 expression did not change while PKIA expression was elevated compared with sh-SIRT1 group (Figure 5A). In dronedarone-mediated influences of Ang II-induced H9C2 cells, western blot assay demonstrated that sh-SIRT1 transfection successfully downregulated the protein level of SIRT1 and PKIA, and oe-PKIA reversed the inhibitory effect of SIRT1 downregulation on PKIA (Figure 5B). For the changes of cell size, SIRT1 inhibition attenuated dronedarone-mediated cell size suppression effect in Ang II-induced H9C2 cells, whereas oe-PKIA abolished the influence of SIRT1 knockdown (Figure 5C and D). Furthermore, SIRT1 inhibition weakened dronedarone-mediated enhancement of FOXO3 and inhibition of ANP, BNP, β-MHC, and PKA expression in Ang II-induced H9C2 cells. However, oe-PKIA reversed the inhibitory effect of sh-SIRT1 on the expression of PKA, ANP, BNP, and β-MHC (Figure 5E). In total, dronedarone suppressed Ang II-induced MH through modulating SIRT1/FOXO3/PKIA axis.
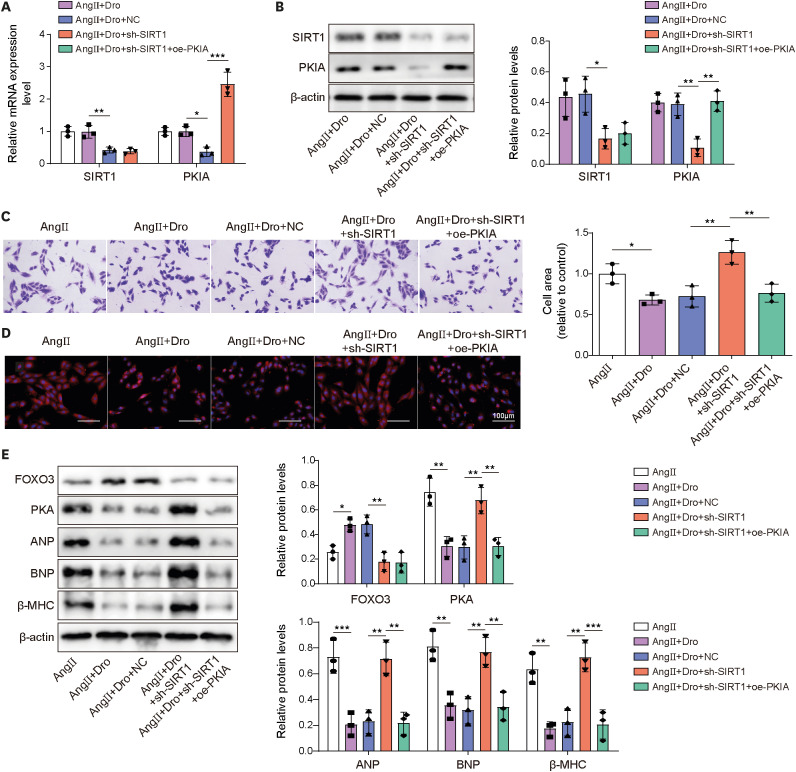
Figure 5
Dronedarone attenuated Ang II-induced MH through regulating SIRT1/FOXO3/PKIA pathway.
H9C2 cells were transfected with sh-SIRT1 or together with oe-PKIA. (A, B) The mRNA and protein expression of SIRT1 and PKIA were evaluated using RT-qPCR and western blot. H9C2 cells were transfected with sh-SIRT1 or together with oe-PKIA and then followed Ang II induction and dronedarone treatment. (C, D) Cell size was evaluated using crystal violet staining and rhodamine phalloidin staining. (E) The protein levels of FOXO3, PKA, ANP, BNP, and β-MHC were examined by western blot.
Ang II = angiotensin II; ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; Dro = dronedarone; FOXO3 = forkhead box O3; NC = negative control; oe- = overexpression; PKIA = protein kinase inhibitor alpha; RT-qPCR = reverse transcription quantitative polymerase chain reaction; sh- = short hairpin RNA targeting; SIRT1 = sirtuin 1; β-MHC = β-myosin heavy chain.
*p<0.05, **p<0.01, ***p<0.001. All data were obtained from 3 independent replications.
![]()
Dronedarone alleviated transverse aortic constriction surgery-induced myocardial hypertrophy and impairment of cardiac function of rats
To investigate whether dronedarone treatment alleviated MH and improved cardiac function, rats received TAC surgery to establish an animal model of MH and 90 mg/kg of dronedarone was given daily by gavage for 8 weeks. As shown in Figure 6A, compared with sham group, the TAC group had enlarged hearts, while the dronedarone effectively reduced sizes of hearts in TAC surgery-treated rats. Besides, H&E staining showed that cardiomyocytes in the sham group were compact (Figure 6B). Cardiomyocytes in the TAC group were hypertrophied and had increased cell space, while those in the dronedarone group were improved (Figure 6B). In addition, TAC surgery resulted in decrease of EF, FS, and heart weight to body weight ratio (HW/BW), which were partially reversed by dronedarone treatment (Figure 6C). Furthermore, TAC surgery led to elevation of ANP and BNP expression and reduction of SIRT1, FOXO3, and PKIA, whereas dronedarone treatment compromised the influences of TAC surgery on those proteins (Figure 6D). Taken together, dronedarone alleviated MH and improved cardiac function of rats via mediating SIRT1/FOXO3/PKIA axis.
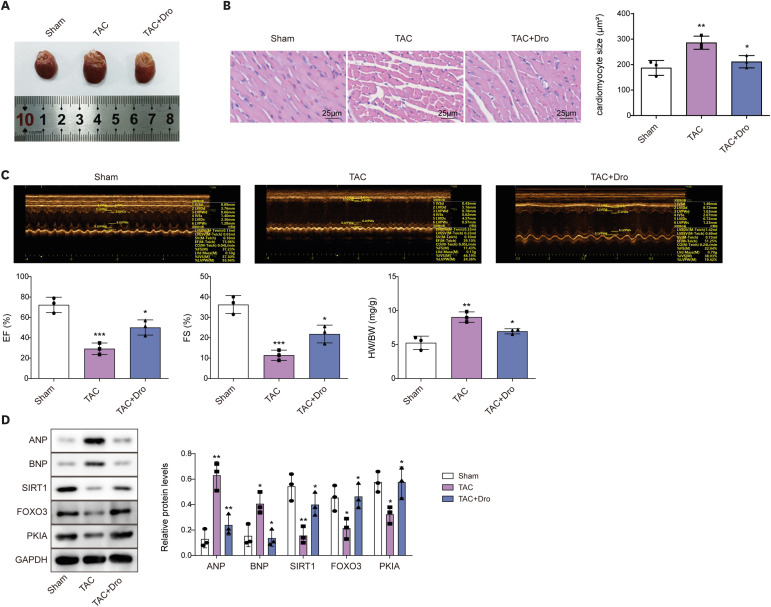
Figure 6
Dronedarone alleviated TAC surgery-induced MH and impairment of cardiac function of rats through regulating SIRT1/FOXO3/PKIA axis.
Rats received TAC surgery to establish an animal model of MH and 90 mg/kg of dronedarone was given daily by gavage for 8 weeks. (A) The images of hearts of rats. (B) Myocardial histomorphology was detected using H&E staining. (C) Representative images of M-mode echocardiography in different groups, EF, FS, and HW/BW were detected. (D) The protein levels of FOXO3, PKIA, ANP, BNP, and SIRT1 were examined by western blot.
ANP = atrial natriuretic peptide; BNP = B type natriuretic peptide; Dro = dronedarone; EF = ejection fraction; FOXO3 = forkhead box O3; FS = fraction shortening; H&E = hematoxylin and eosin; HW/BW = heart weight to body weight ratio; MH = myocardial hypertrophy; PKIA = protein kinase inhibitor alpha; SIRT1 = sirtuin 1; TAC = transverse aortic constriction.
*p<0.05, **p<0.01, ***p<0.001. All data were obtained from 3 independent replications.
![]()
DISCUSSION
Dronedarone has similar pharmacological properties to amiodarone and is considered to be an effective and safe drug for the treatment of atrial fibrillation because it does not contain iodine.24) Currently, dronedarone was found to be involved in myocardial remodeling and bioactivities of endothelial progenitor cells,5)25) indicating dronedarone is a promising medicine in cardiovascular diseases in addition to atrial fibrillation. However, there has few studies about dronedarone affecting MH. Therefore, in this study, we focused on functions and the underlying mechanism of dronedarone in MH. It was noting that Ang II has been widely used to induce models of MH.26)27) In this study, Ang II was used to establish cell model of MH. Our results validated that Ang II treatment enlarged cellular morphology, enhanced the expression of hypertrophy-related indicators and decreased cell viability, showing that cell model of MH was successfully established. Furthermore, our findings found that dronedarone resulted in decreased cell size of Ang II-induced H9C2 cells and notably suppressed hypertrophy-related genes including ANP, BNP, β-MHC, and alleviated TAC surgery-induce MH of rats and impairment of cardiac function, suggesting that dronedarone could restrained MH progression. Amiodarone, also is an antiarrhythmic drug and shares similar structure with dronedarone.21) Here, we found that amiodarone decreased the expression of hypertrophy-related genes including ANP, BNP, β-MHC in H9C2 cells, which were similar to previous researches.22)23) Additionally, we observed that the expression of SIRT1 in H9C2 cells was inhibited by Ang II stimulation, however, dronedarone and amiodarone could abolish Ang II-induced reduction of SIRT1 expression in H9C2 cells. Therefore, we that SIRT1 might be a targeted gene of dronedarone in mediating MH suppression.
At present, SIRT1 was investigated in MH progression. For example, Zhu et al.28) reported that SIRT1 was evidently decreased by Ang II induction and lncRNA NBR2 activated LKB1/AMPK/SIRT1 pathway to repress endoplasmic reticulum stress, thereby attenuating Ang II-induced MH. Ginsenoside Rg3 upregulated SIRT1 to suppress Ang II-induced MH through inhibiting NLRP3 inflammasome and oxidative stress.20) Consistent with previous results, Ang II induction resulted in decreased SIRT1 expression and induced MH of H9C2 cells, which were reversed by SIRT1 upregulation. Moreover, we observed that Ang II-induced decreased SIRT1 expression was restored by dronedarone. Further experiments proved that oe-SIRT1 reinforced the inhibiting effects of dronedarone on MH. These evidences indicated that dronedarone might suppress MH progression through upregulation of SIRT3.
Previous researches have proved SIRT1-mediated FOXO3 deacetylation to upregulate FOXO3 expression, thereby affecting various diseases. For instance, TSPYL2 suppressed SIRT1-mediated FOXO3 deacetylation to restrain resistance to gefitinib and DNA damage in colorectal cancer.29) Moreover, Angiotensin-(1-7) elevated FOXO3 to prevent T3-induced MH.30) These studies indicated that FOXO3 might be mediated by SIRT1 affecting FOXO3 deacetylation, thereby playing a suppressive role in MH progression. In this work, oe-SIRT1 or dronedarone led to increased FOXO3 expression and decreased FOXO3 acetylation in Ang II-induced H9C2 cells, suggesting dronedarone activating SIRT1/FOXO3 axis to inhibit MH progression. Of note, our results validated that FOXO3 could interact with PKIA promoter to promote PKIA expression. Furthermore, in Ang II-induced H9C2 cells, the expression of PKIA was elevated by oe-SIRT1 and/or dronedarone. PKIA was reported to be a downstream gene of miR-129-3p and Angiotensin-(1-9) upregulated miR-129-3p, inhibited PKIA and promoted AKT activity to prevent MH.18) Currently, this was the only research to report that PKIA may be associated with MH. Our findings revealed that oe-PKIA could attenuate Ang II-induced cell hypertrophy and its overexpression reinforced the inhibiting influences of dronedarone on cell hypertrophy. Moreover, SIRT1 silencing-mediated promotion of cell hypertrophy in Ang II-induced H9C2 cells upon dronedarone were impaired by oe-PKIA.
Regarding the PKIA-related results, our study was associated with the study by Xiaoying Zhang et al.,15) that was, PKIA could alleviate MH, which contradicted the study of Sotomayor-Flores et al.18) In the study of Sotomayor-Flores et al.,18) in the model of MH, inhibition of PKA did not significantly affect the degree of MH. Interestingly, however, in the presence of Ang-(1-9), inhibition of PKA also affected the degree of MH.18) The experimental methods and content of the 2 studies were not quite the same, so it is difficult for us to find the reason for the difference in results. However, in future studies, we will conduct more MH models and perform relevant sequencing, detect the expression differences of the molecules involved, and obtain more relevant regulatory targets.
It was noting that, our results demonstrated that dronedarone alleviated MH at least in part by regulating the expression of SIRT1. Cell hypertrophy was significantly inhibited in the group treated with dronedarone and oe-SIRT1 at the same time, indicating that dronedarone may alleviate MH by mediating other pathways. To seek out other mechanism of dronedarone on improving MH, in the future, we will sequence the cardiomyocytes or rat myocardial tissues treated with dronedarone to obtain more targets and pathways of dronedarone.
In conclusion, our study firstly elucidated that dronedarone alleviated MH through regulating SIRT1/FOXO3/PKIA axis, which suggested that dronedarone might become promising drug for MH and targeting SIRT1/FOXO3/PKIA axis might inhibit MH progression.
 XML Download
XML Download