

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Inflammation is an immune response in the human body, causing inflammatory pain due to persistent inflammation and continuous noxious stimuli activating neuronal cell bodies [1,2]. The sensitivity of inflammatory pain depends on tissue injury, inflammatory cell infiltration, and the release of inflammatory mediators, which promote peripheral nerve stimulation and pain [3–5]. The most commonly used pain relievers are non-steroidal anti-inflammatory drugs (NSAIDs). Nevertheless, considering the long-term treatment perspective, these drugs can have negative effects on the digestive system, cardiovascular system, liver, and kidney function [6,7]. Therefore, it is necessary to search for and develop novel analgesic drugs that are safer and more effective than NSAIDs, as they have shown vast research potential and promising prospects for application. Recently, natural compounds have gained significant attention. This preference is primarily attributed to their safety and therapeutic efficacy. Alpinia officinarum, a plant-derived remedy, holds remarkable significance in many Asian nations. It serves as a valuable adjunct to conventional pharmacotherapy for a spectrum of conditions, including but not limited to abdominal pain, gastritis, arthritis, dysmenorrhea, diabetes, and various inflammatory and painful disorders [8,9]. However, current knowledge of the analgesic components of highland ginger and the molecular targets that mediate its pharmacological properties remains limited.
Galangin, a flavonoid derived from A. officinarum, exhibits noteworthy anti-inflammatory properties; however, its specific activity and role in the context of alleviating inflammatory pain remain incompletely explored and validated. This experimental study aimed to elucidate the analgesic properties of galangin in a murine model and to evaluate its potential as an alternative to NSAIDs for the treatment of inflammatory pain. The effectiveness of galangin in reducing inflammatory pain using mouse models was investigated. We assessed its binding to key targets through molecular docking, and the impact of galangin was evaluated using rat models, with or without capsazepine, a selective transient receptor potential vanilloid 1 (TRPV1) antagonist. Inflammatory pain-associated factors were measured using enzyme-linked immunosorbent assay (ELISA), Western blot, and real-time quantitative PCR (qPCR) techniques. As shown in Fig. 1, our overall experimental results and conclusions were presented.
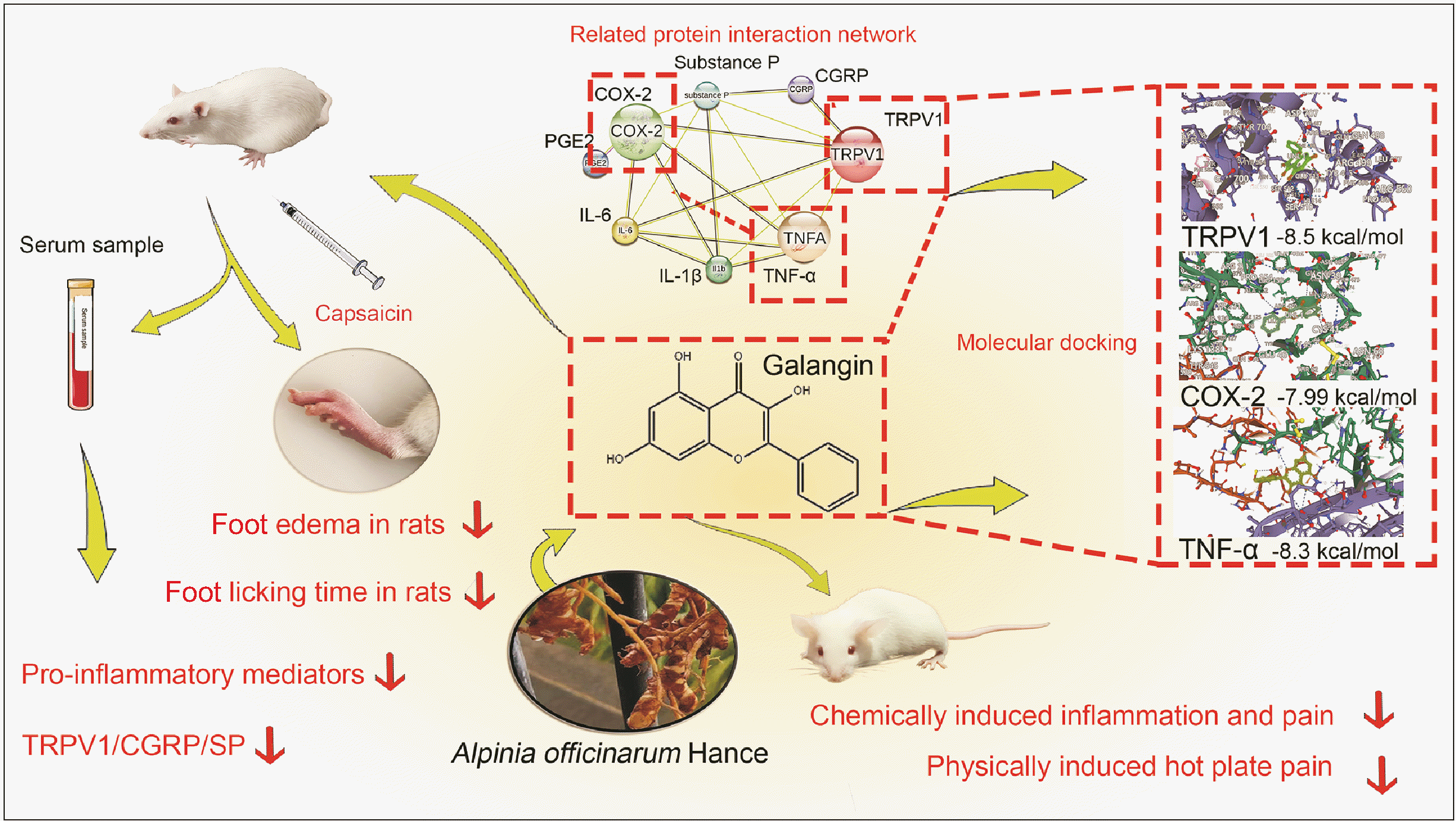
Go to : 
MATERIALS AND METHODS
1. Drugs and chemicals
Galangin (NO: G100561) and Carrageenan (NO: C107615) were purchased from Shanghai Aladdin Biochemical Technology Co., LTD. Formalin reagent (1000337) and glacial acetic acid (1000078) were purchased from Guangxilong Science Co., LTD. Celecoxib capsule (NO: 230802) was purchased from Jiangsu Zhengda Qingjiang Pharmaceutical Co., LTD.
2. Animals
A total of sixty female BABL/c mice (4–6 weeks old, weighing 18–22 g) and forty-eight male and female Sprague–Dawley rats (6–8 weeks old, weighing 180–200 g) were sourced from the Guangdong Medical Laboratory Animal Center in Guangdong, China (License: SCXK (Yue) 2022-0002). The animals were provided with standard rodent feed and water ad libitum and housed under controlled conditions (12 hr light/dark cycle, temperature: 22°C ± 2°C, humidity: 40%–60%). This study followed the guidelines of Hainan Medical University Medical University and received ethical approval from its Ethics Committee (NO. HYLL-2022-266, Haikou, China). All animal handling procedures were carried out in accordance with the regulations for laboratory animal care and utilization set by the National Institutes of Health.
3. Animal experimental design
In this experiment, galangin and celecoxib were dissolved in a 0.5% solution of carboxymethylcellulose sodium. After 2 days of acclimation, the animals were randomly divided into different experimental groups. For the nociceptive behavior tests in the mouse model, the 60 mice were divided into four groups with 15 mice in each group: (1) the control group (mice received 0.5% carboxymethylcellulose sodium), (2) the celecoxib group (mice received celecoxib at a dose of 20 mg/kg), (3) the galangin low-dose group (mice received galangin at a dose of 25 mg/kg), and (4) the galangin high-dose group (mice received galangin at a dose of 50 mg/kg). All rats were treated with the corresponding drugs (different concentrations of galangin and celecoxib) or 0.5% carboxymethylcellulose sodium for seven consecutive days; For the study on pain induced by capsaicin and the protective mechanism of galangin in rats, the 48 rats were divided into six groups with 8 rats in each group: (1) the control group (rats received 0.5% carboxymethylcellulose sodium), (2) the model group (rats received 0.5% carboxymethylcellulose sodium), (3) the capsazepine group (rats received the TRPV1 antagonist capsazepine at a dose of 2 mg/kg, 200, intravenous injection), (4) the celecoxib group (rats received celecoxib at a dose of 20 mg/kg), (5) the galangin low-dose group (rats received galangin at a dose of 25 mg/kg), and (6) the galangin high-dose group (rats received galangin at a dose of 50 mg/kg). The corresponding drugs were administered via continuous gavage or intraperitoneal injection for 7 days. Rats in all groups, except the control group, were then injected with 0.5% capsaicin (TRPV1 agonist).
4. Analgesic and anti-inflammatory effects were assessed by nociceptive behavior tests
1) Hot plate test
To assess pain sensitivity, an intelligent hot plate instrument (model YY92-YLS-6B; Dongfanghua Glass Technology Co., LTD.) was utilized with a precise temperature setting of 55°C ± 0.5°C. The stress response time was defined as the duration between the initiation of jumping or paw licking by the mice. During the experiment, the mice were orally administered either saline, galangin (at doses of 25 and 50 mg/kg), or celecoxib (at a dose of 20 mg/kg) for 7 consecutive days. To evaluate the effects of the treatment, the hot plate test was conducted one hour after the final drug administration on day 7. The latency of pain response was measured at 30, 60, 90, 120, 150, 180, and 210 minutes. It is important to note that a cut-off time of 40 seconds was implemented to prevent potential thermal injury to the mice. The outcomes were expressed as the mean percent maximal effect (%MPE), which was calculated using the following equation (1):
2) Acetic acid-induced abdominal writhing test
One hour after the seventh-day drug administration, the mice were intraperitoneally injected with a 0.6% v/v acetic acid solution (10 mL/kg, intraperitoneal injection). Within a 30-minute timeframe following the acetic acid injection, the number of abdominal twists (including abdominal contractions, torso/pelvis contortions, etc.) was recorded for each animal. The results were expressed as the mean percentage inhibition of writhing (%PIW), which was calculated using the following equation (2):
3) Acetic acid-induced vascular permeability
After the seventh day of drug treatment, each mouse received a single intravenous injection of 1% Evans blue at a dosage of 0.1 mL/10 g body weight. Subsequently, precisely one hour after the drug treatment, each mouse was injected with acetic acid at a concentration of 0.6% and a dosage of 0.1 mL/kg into the abdomen. Thirty minutes post-injection, the mice were euthanized under anesthesia and subjected to three washes with a 5 mL normal saline solution. After centrifugation, the optical density of the supernatant was measured at 540 nm. The degree of inhibition of vascular permeability was determined using the following formula (3):
4) Formalin-induced plantar pain test
One hour after the seventh day of drug treatment, a micro-injector was utilized to withdraw 50 μL of a 5% formalin solution. The solution was then slowly injected into the subcutaneous area of the mouse's right back palm, creating a dermal mound. The mice's spontaneous nociceptive responses, such as foot lifting, foot licking, and foot movement, were observed at 5-minute intervals for a duration of 60 minutes following the formalin injection. The number of pain responses occurring within a 1-minute timeframe was recorded on each occasion.
5) Carragenan foot swelling experiments
One hour after the seventh day of drug treatment, the mice were intradermally administered a 0.1 mL carrageenan solution (1% w/v) into the plantar surface to induce foot edema. Subsequently, the foot thickness was measured at 1, 2, 3, and 4 hours after the carrageenan injection using a digital volume analyzer. The rate of foot thickening-induced edema was calculated using the following equation (4):
Where “Va” is the thickness at different time points after injection and “Vb” is the thickness before injection.
6) Capsaicin-induced inflammatory pain test
On the seventh day, one hour after the last treatment, a subcutaneous injection of capsaicin (TRPV1 agonist) at a concentration of 0.5% was administered into the right hind paw. The total time of paw licking within five minutes after the injection was recorded.
5. Analysis of expression of TRPV1 receptor and cyclooxygenase-2 (COX-2), nuclear factor kappa B (NF-κB)/tumor necrosis factor-α (TNF-α) and other related factors
1) Molecular docking between galangin and TRPV1, COX-2, NF-κB and TNF-α
Autodock Vina 1.2.2, a computational software for protein-ligand docking, was used for this study. The molecular structure of galangin was obtained from the PubChem compound database (https://pubchem.ncbi.nlm.nih.gov/). The 3D coordinates of TRPV1 (PDB number: 3J5R, resolution: 4.20 Å), COX-2 (PDB number: 4PH9, resolution: 1.81 Å), NF-κB p65 (PDB number: 1RAM, resolution: 2.7 Å), and TNF-α (PDB number: 2AZ5, resolution: 2.10 Å) were downloaded from the Protein Data Bank (PDB) (http://www.rcsb.org/). The molecular docking studies were visualized using Autodock Vina 1.2.2 (http://autodock.scripps.edu/).
2) Mechanism analysis of galangin in treating inflammatory pain
As described in Section 6 of Chapter 4 in the “MATERIALS AND METHODS”, inflammation-induced pain in rats was induced by administering capsaicin. After appropriate pharmacological treatment, serum samples were obtained from the abdominal aorta under deep isoflurane anesthesia. Additionally, segments L4-5 of the dorsal root ganglion, as well as tissues from the injected hindfoot and plantar areas of the rats, were immediately excised and stored at –80°C for subsequent biochemical analysis.
3) Enzyme-linked immunosorbent assay
The concentrations of TNF-α, interleukin (IL)-6, IL-1β, prostaglandin E2 (PGE2), calcitonin gene-related peptide (CGRP), and substance P (SP), as well as the enzymatic activity of COX-2, were measured using an ELISA kit sourced from Elisa Biotech. All assays were performed according to the manufacturer's recommended protocols. Absorbance readings were acquired using a microplate spectrophotometer manufactured by BioTek Instruments, Inc.
4) Analysis of quantitative real-time PCR
RNA was extracted from the dorsal root ganglion and plantar tissues of mice using the Eastep® Super Total RNA Kit. The extracted RNA was then subjected to reverse transcription using the cDNA Synthesis SuperMix (YEASEN). Subsequently, the resulting cDNA was amplified using the precision Agilent Mx 3005p qPCR system. The obtained data was analyzed using the cycle threshold method. The primer sequences used in the amplification process are provided in Table 1.
Table 1
The mRNA primer sequence list for real-time quantitative PCR (qRT-PCR) in the experiment
![]()
5) Analysis of Western blot
After lysing the dorsal root ganglion tissues of mice using a protein lysate, the proteins were extracted and their concentrations were determined using a bicinchoninic acid protein analysis kit (Beyotime). The proteins were then separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (MilliporeSigma). To block non-specific binding, the membranes were sealed with a 5% bovine serum albumin solution at room temperature for 1 hour. Subsequently, the membranes were incubated overnight at 4°C with the corresponding primary antibody (TRPV1, 1:1,000 dilution, Abcam, ab305299). On the following day, the membranes were incubated with a horseradish peroxidase-labeled secondary antibody for 1.5 hours. Specific bands were detected using a Gel imaging system (Bio-Rad) and a chemiluminescence detection reagent (Beyotime).
6. Statistical analysis
All experimental data were presented as mean ± standard deviation. Statistical analysis was performed using GraphPad Prism 8.0 software (GraphPad Software) with a one-way analysis of variance. A Tukey test was conducted to determine the level of significance between the groups. A P value of < 0.05 was considered statistically significant, indicating a significant difference in the data.
Go to :
RESULTS
1. Hot plate test
The thermal stimulation latency of mice pretreated with galangin and celecoxib was significantly increased compared to untreated mice at 30, 60, 90, 120, 150, 180, and 210 minutes (Fig. 2A). Both doses of galangin (25 and 50 mg/kg) reached the maximum average thermal stimulation latency at 180 minutes, with corresponding average analgesic effect percentages of 43% and 58%, respectively, which were lower than that of celecoxib (61.2%) (Fig. 2B). Additionally, at 180 and 210 minutes, the 50 mg/kg dose of galangin (with maximum effect percentages of 58% and 55.8%, respectively) exhibited an effect comparable to that of the 20 mg/kg dose of celecoxib (with maximum effect percentages of 61.2% and 59.8%, respectively).
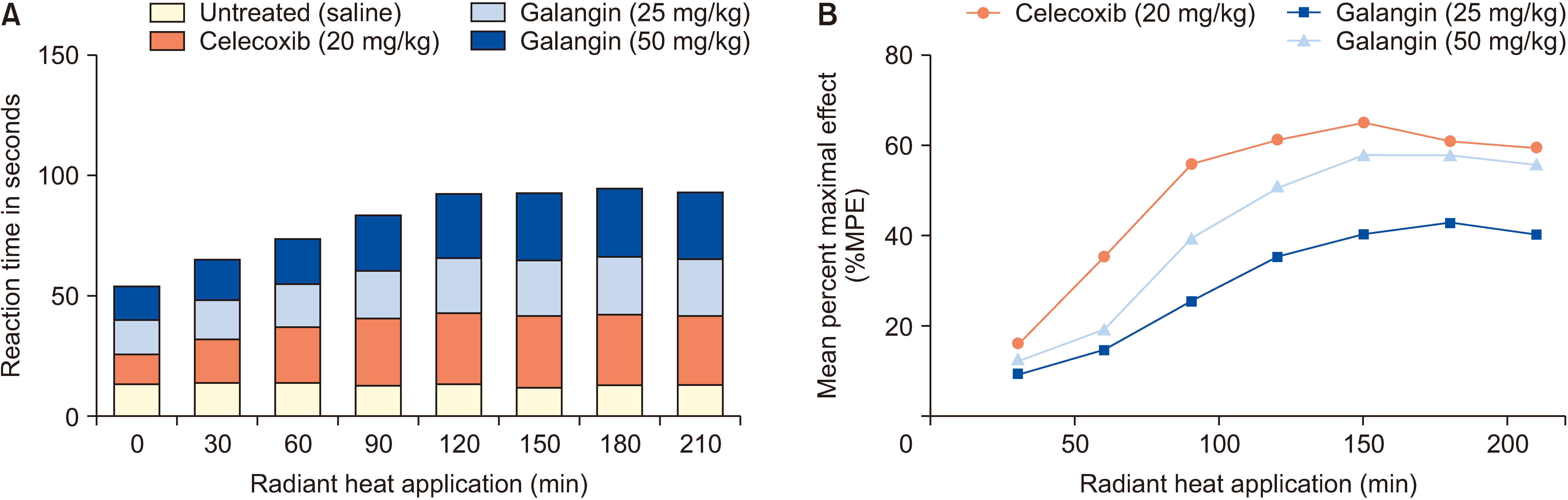 | Fig. 2Effect of galangin on hot plate experiment. (A) During different time intervals within 210 minutes, the effects of galangin and celecoxib on the latency period of the hot plate in mice. (B) Average percentage of maximum effect of the drug (%MPE). Data were expressed as means ± standard error of the mean (n = 6).
|
2. Acetic acid-induced abdominal writhing test
The analgesic properties of galangin are illustrated in Fig. 3A, B. Abdominal writhing experiments revealed a dose-dependent antinociceptive effect, indicating a reduction in body twisting. Administration of galangin at a dosage of 25 mg/kg resulted in a total of 29.5 twists with an average twist inhibition rate of 28.8%. Importantly, both galangin at a dosage of 50 mg/kg and celecoxib exhibited similar antinociceptive effects, leading to 20.5 and 20.1 instances of twisting, respectively, accompanied by average twist inhibition rates of 50.2% and 50.9%, respectively.
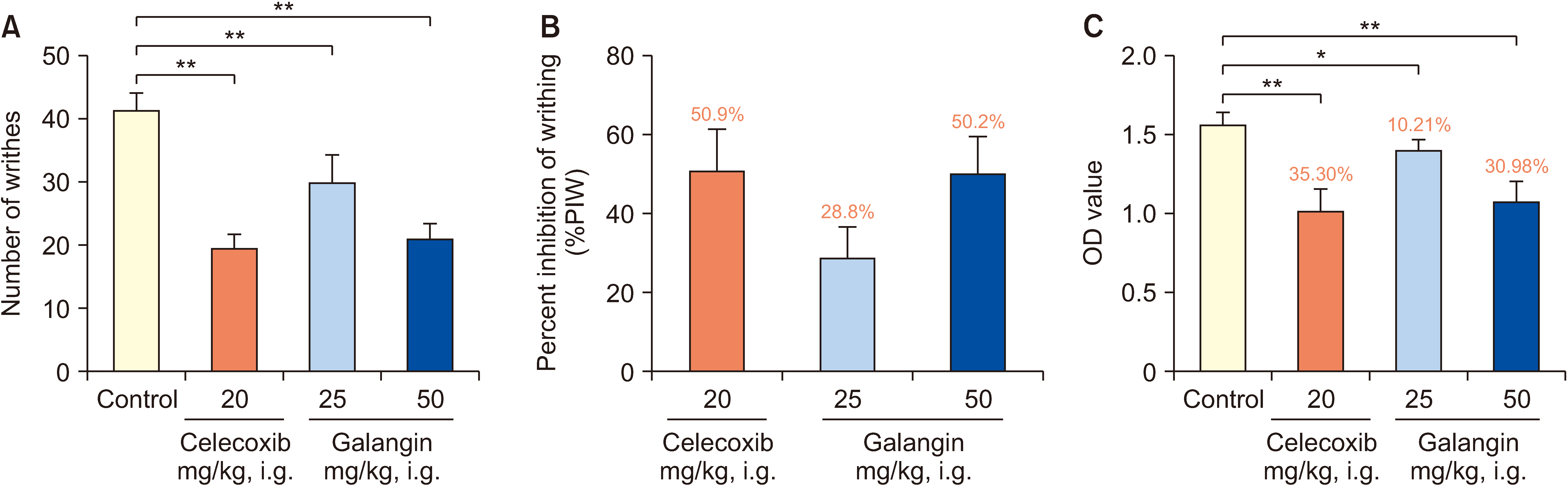 | Fig. 3Effect of galangin on acetic acid induction test. (A) Effect of galangin on twisting times of mice. (B) Effect of drugs on torsion inhibition rate. (C) Effects of drugs on vascular permeability. Compared with control group, *P < 0.05 and **P < 0.01 (n = 6). OD: optical density, i.g.: intragastrical administration.
|
3. Acetic acid-induced vascular permeability
Fig. 3C depicts the effects of galangin on acetic acid-induced peritoneal capillary permeability in mice. Galangin demonstrated a dose-dependent decrease in vascular permeability compared to the model group. The inhibition rates for the dosages of 25 mg/kg and 50 mg/kg of galangin were 10.21% and 30.98%, respectively. Notably, the 50 mg/kg galangin group exhibited an inhibition rate comparable to that of the celecoxib group (35.30%).
4. Formalin-induced plantar pain test
The efficacy of galangin in reducing the duration of formalin-induced neuropathic (0–15 minutes, stage 1) and inflammatory (20–30 minutes, stage 2) pain in mice was demonstrated through treatment with doses of 25 and 50 mg/kg (Fig. 4). In the initial stage of the formalin test (Fig. 4A), mice administered 0.5% carboxymethylcellulose sodium exhibited a licking time of 75.66 ± 5.18. However, mice treated with galangin at doses of 25 and 50 mg/kg displayed durations of 42.6 ± 1.63 (reduction rate of 43.6%) and 33.7 ± 4.72 (reduction rate of 55.5%), respectively. During the second stage (Fig. 4B), the 0.5% carboxymethylcellulose sodium group had a licking time of 150.5 ± 7.13, while galangin-treated mice recorded durations of 70 ± 4.69 (reduction rate of 53.5%) and 57.7 ± 4.27 (reduction rate of 61.7%). Furthermore, the positive control group treated with celecoxib at 20 mg/kg exhibited a licking time of 49 ± 1.41 (reduction rate of 35.2%) during the first stage and 48 ± 3.68 (reduction rate of 68.1%) during the second stage.
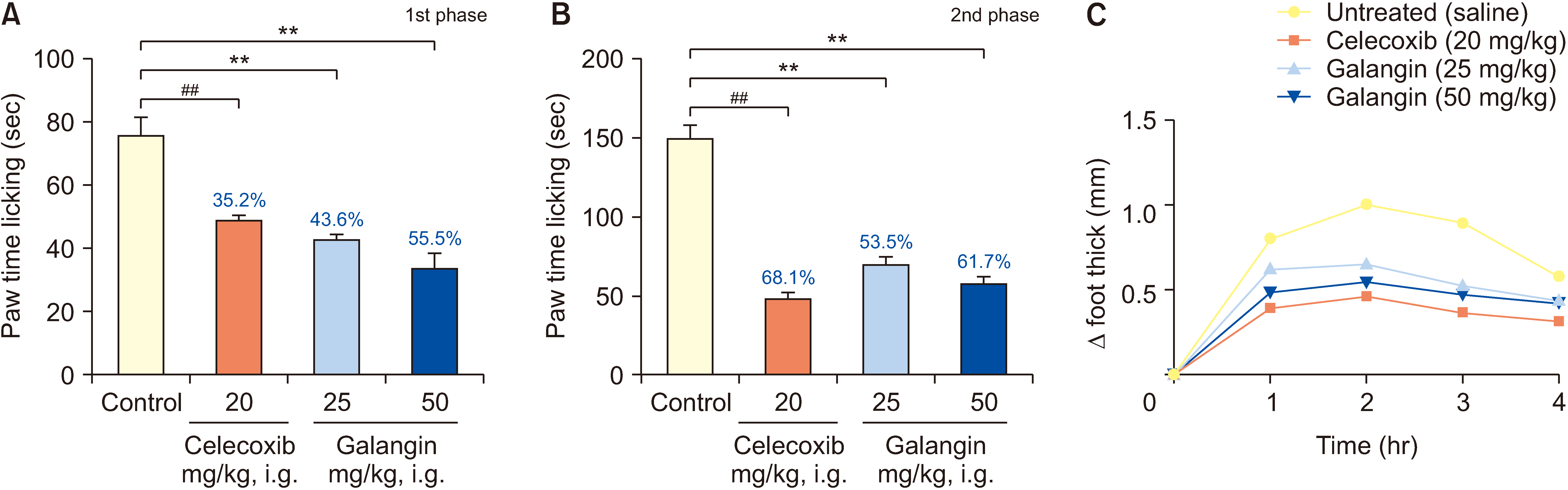 | Fig. 4The experimental effect of galangin on formalin pain and carrageenan foot swelling. (A) The effect of drugs on first-phase pain caused by formalin. (B) The effect of drugs on the pain caused by formalin in the second phase. (C) The effect of drugs on the inhibition rate of carrageenan induced foot swelling in mice. Compared to the control group, **P < 0.01; ##P < 0.01, (n = 6). i.g.: intragastrical administration.
|
5. Carragenan foot swelling experiments
The administration of plantar carrageenan injection in mice (model group) resulted in a significant and time-dependent increase in hindfoot thickness, as depicted in Fig. 4C. Notably, mice treated with galangin showed a significant reduction in foot thickness at 1–4 hours. The peak of foot inflammation occurred in the second hour after carrageenan infusion. Subsequently, the groups treated with galangin (25 mg/kg and 50 mg/kg) and celecoxib (20 mg/kg) exhibited comparable suppressive effects on hindfoot edema in mice.
6. Capsaicin-induced inflammatory pain test
Treatment with galangin at both doses (25 and 50 mg/kg) resulted in a significant reduction in the licking time of capsaicin-induced mice, as depicted in Fig. 5. The 0.5% carboxymethylcellulose sodium group had a licking time (s) of 36.55 ± 3.9, whereas the groups treated with galangin at 25 mg/kg and 50 mg/kg exhibited licking times of 21.11 ± 3.21 (30.19% reduction) and 13.92 ± 2.78 (49.58% reduction), respectively. Importantly, galangin exhibited a more pronounced effect compared to the celecoxib-treated group, which had a licking time of 28.01 ± 2.82. The capsazepine group also showed a decrease in licking time by 9.07 ± 1.72 times (a reduction of 30.19%). Additionally, the administration of galangin and capsazepine demonstrated inhibitory effects on capsaicin-induced foot edema in rats.
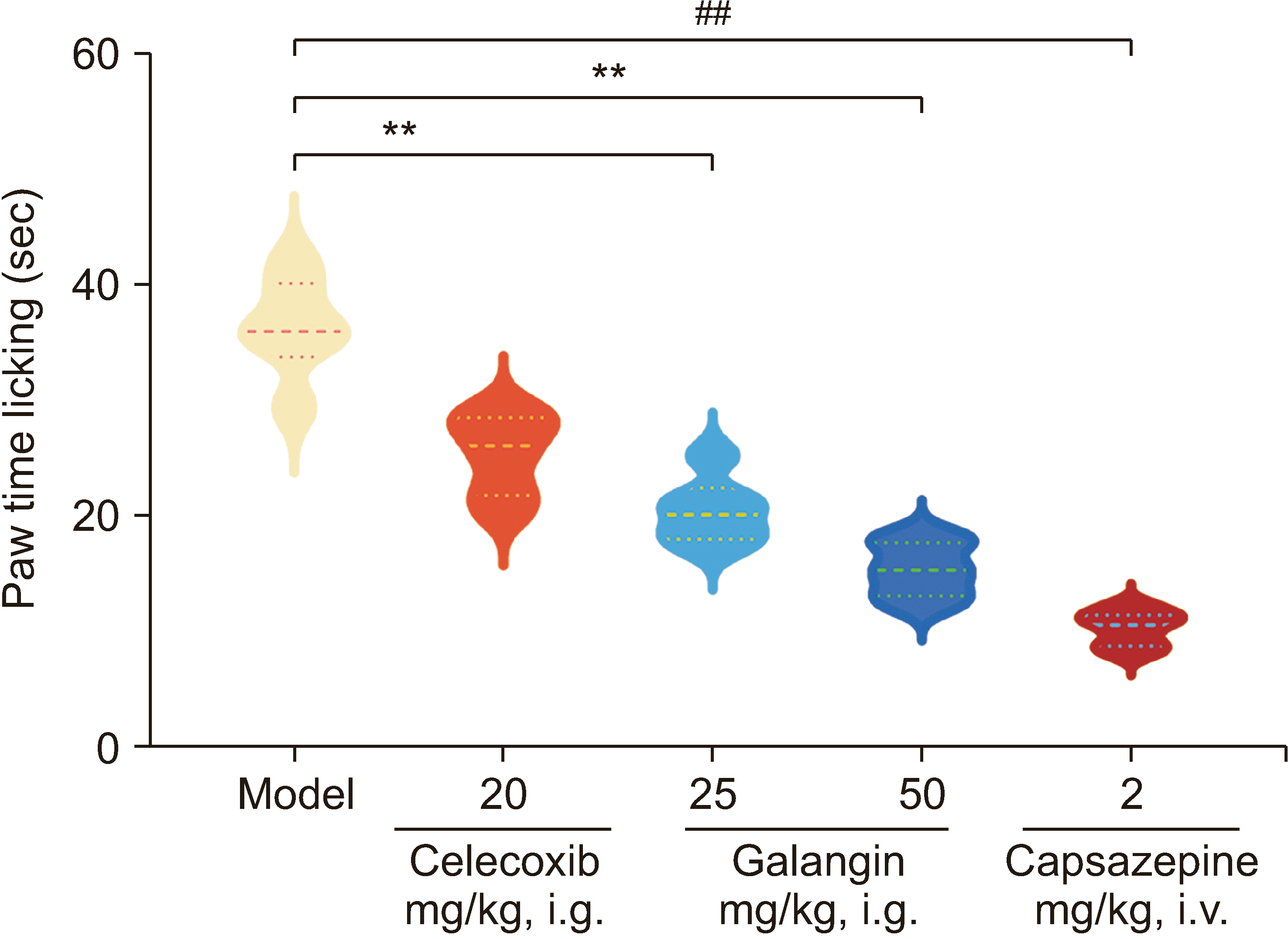
7. Galangin inhibits capsaicin-induced inflammatory factor release
In comparison to the model group, mice treated with galangin exhibited a significant decrease in pro-inflammatory factors, including IL-1β, IL-6, TNF-α, PGE 2, CGRP, and SP levels, as well as COX-2 activity in serum, as illustrated in Fig. 6. Similarly, the capsazepine group demonstrated a substantial effect in reducing inflammatory factors, comparable to that observed in the high-dose group.
 | Fig. 6Galangin inhibits the release of capsaicin-induced related factors. (A–E) The levels of inflammatory factors IL-1β, IL-6, TNF-α, PGE2, and the active concentration of COX-2 in serum. (F) Effect of galangin on CGRP and SP levels in serum. ##P < 0.01 compared with control group; **P < 0.01 compared with model group, (n = 6). IL: interleukin, TNF-α: tumor necrosis factor-α, PGE2: prostaglandin E2, COX-2: cyclooxygenase-2, CGRP: calcitonin gene-related peptide, SP: substance P, i.g.: intragastrical administration, i.v.: intravenous injection.
|
8. Effect of galangin on capsaicin-induced tissue-associated mRNA expression in mouse dorsal root ganglia
In the qPCR experiment (Fig. 7A), treatment with galangin led to a decrease in the expression levels of TRPV1, CGRP, NF-κB, TNF-α, COX-2, and PGE2, with a more pronounced effect observed in the high-dose treatment group. Importantly, the capsazepine group exhibited inhibition of TRPV1 expression, as well as its associated receptors CGRP and SP. Additionally, this group demonstrated a reduction in the mRNA expression levels of NF-κB, TNF-α, COX-2, and PGE2.
 | Fig. 7Molecular docking and PPI network recognition of key genes; Western blot and qPCR validation experiments. (A) Galangin inhibited mRNA expression induced by capsaicin. (B) Molecular docking of galangin with TRPV1, NF-κB, COX-2 and TNF-α. (C, D) Galangin inhibited the expression of TRPV1 protein in dorsal root ganglion induced by capsaicin. (E) PPI network recognition of key genes. #P < 0.05 and ##P < 0.01 compared with the normal group; *P < 0.05 and **P < 0.01 compared with the model group. Values are expressed as means ± standard deviation (n = 3). PPI: protein-protein interaction network, TRPV1: transient receptor potential vanilloid 1, NF-κB: nuclear factor kappa B, COX-2: cyclooxygenase-2, TNF-α: tumor necrosis factor-α, CGRP: calcitonin gene-related peptide, SP: substance P, PGE2: prostaglandin E2, i.g.: intragastrical administration, i.v.: intravenous injection.
|
9. Effect of galangin on capsaicin-induced TRPV 1 protein in rats dorsal root ganglion tissue
Through docking analysis, it was determined that galangin exhibits molecular affinity towards its target proteins, namely TRPV1, NF-κB, COX-2, and TNF-α. This affinity is evident through the formation of hydrogen bonding interactions and robust electrostatic interactions. The calculated binding energies further support this observation, with galangin displaying high affinity towards TRPV1 (–8.5 kcal/mol), NF-κB (–7.52 kcal/mol), COX-2 (–7.99 kcal/mol), and TNF-α (–8.3 kcal/mol), respectively. These findings indicate a stable binding of galangin to TRPV1, NF-κB, COX-2, and TNF-α, emphasizing the strength of their interactions (Fig. 7B). Furthermore, western blotting analysis revealed that the expression of TRPV1 protein in the dorsal root ganglia of rats was consistent with the effect of capsazepine, showing an inhibition of TRPV1 protein expression (Fig. 7C, D).
Go to :
DISCUSSION
Through extensive research and clinical application, natural botanical remedies have demonstrated distinct efficacy and reduced toxicity in the treatment of various ailments, gaining gradual acceptance among individuals [10,11]. As one of the most active ingredients in A. officinarum, galangin has shown important effects in the prevention and treatment of many diseases, including but not limited to anti-cancer, regulation of apoptosis and inflammation, cardiovascular diseases, and neurodegenerative diseases [12–20]. Given the limited research on the therapeutic effects and potential mechanisms of galangin in anti-inflammatory pain, this study aimed to evaluate its anti-inflammatory and analgesic effects and explore its mechanism pathways. The objective was to ascertain whether galangin, which is safe, effective, and readily available, could serve as a potential alternative pharmaceutical agent for anti-inflammatory and analgesic treatments. Moreover, celecoxib has been demonstrated to effectively inhibit epoxide activity, leading to a reduction in prostaglandin synthesis. Due to its well-established role in treating inflammation and pain, the inclusion of celecoxib in this study would enhance the ability to evaluate the anti-inflammatory and analgesic effects of galangin, as well as investigate any potential similarities in their mechanisms and effects.
In this study, a range of methods were employed to assess the anti-inflammatory and analgesic effects of galangin. The investigation commenced with the utilization of the hot plate method, which is a thermal nociceptive model used to evaluate central analgesic efficacy. It was observed that the incubation period of galangin and celecoxib was longer compared to the control group. Moreover, the mean maximum effect (%MPE) percentage in the hot plate test exhibited a significant dose-dependent increase in the galangin group. These findings suggest that galangin possesses the ability to inhibit central analgesia. Additionally, at 180 and 210 minutes, the administration of a 50 mg/kg dose of galangin resulted in a similar effect to that of a 20 mg/kg dose of celecoxib, with maximum effect percentages at 180 and 210 minutes measured at 58% and 55.8%, respectively, for galangin and 61.2% and 59.8%, respectively, for celecoxib.
Furthermore, the acetic acid-induced abdominal torsion test was employed, which is a nociceptive model used to evaluate the efficacy of peripherally acting analgesics. The findings revealed that galangin exhibited a dose-dependent decrease in the torsion inhibition rate (%PIW) at doses of 25 and 50 mg/kg, consistent with previous studies on the anti-inflammatory and analgesic properties of natural plants [21,22]. It has been demonstrated that acetic acid stimulation can increase the permeability of abdominal capillaries, leading to acute inflammation and pain [23]. Therefore, an acute inflammation and pain model was established by inducing vascular permeability using acetic acid and measured the permeability of abdominal capillaries. The results demonstrated that galangin effectively inhibited the increase in abdominal vascular permeability and alleviated acute inflammation and pain. Based on these studies, it can be inferred that galangin may reduce the synthesis of prostaglandins and inhibit the activation of pain nerve endings by IL-6 and TNF-α, thereby exerting a peripheral analgesic effect [24]. Additionally, the formalin plantar pain test was utilized, which is a model used to induce peripheral nociceptive stimulation [25]. The results demonstrated that galangin exhibited a significant dose-dependent reduction in the duration of licking stress during both Stage I and Stage II of the formalin test. Furthermore, galangin demonstrated superior acute pain inhibition compared to celecoxib during Phase I. In Stage II inflammatory pain, the efficacy of galangin was comparable to that of the standard drug celecoxib, with a 68.1% inhibitory effect. These findings suggest that galangin possesses the ability to effectively inhibit pain during both the early neurogenic and inflammatory stages. Additionally, we employed the carrageenan foot swelling assay, a well-established model for evaluating anti-inflammatory effects by measuring the inhibition of acute inflammatory mediators. In this assay, galangin demonstrated a significant reduction in foot edema volume. Based on the results obtained from both the formalin-induced plantar pain test and the carrageenan foot swelling test, it can be hypothesized that the anti-inflammatory and analgesic activity of galangin may be attributed to its ability to inhibit pro-inflammatory mediators such as histamine and prostaglandins, thereby exerting an inhibitory effect on acute inflammation [26,27].
Upon discovering the ability of galangin to inhibit the synthesis of pro-inflammatory mediators, specifically histamine and prostaglandins, induced by carrageenan and formalin, as well as its capacity to hinder acute inflammation and prevent the activation of pain-sensing nerve endings, this investigation delved deeper into the serum and local tissue levels of TNF-α, IL-6, IL-1β, IL-10, COX-2, and PGE2 in rats experiencing capsaicin-induced inflammatory pain. These findings revealed that galangin effectively suppresses the production and levels of these pro-inflammatory mediators, which aligns with previous studies on the efficacy of anti-inflammatory analgesic drugs [26,28]. This further suggests that galangin may inhibit cyclooxygenase activity, thus reducing prostaglandin synthesis and suppressing the levels of pro-inflammatory mediators to impede the development of inflammatory pain. Our results demonstrate that galangin exhibits anti-inflammatory and analgesic effects against various stimuli, including chemical and physical induction, and displays notable inhibitory activity against capsaicin-induced neuropathic pain.
TRPV1 plays a crucial role in the transmission and modulation of pain signals in both the peripheral and central nervous system, and it also plays a significant role in the development of inflammation-induced thermal hyperalgesia. Upon stimulation by neurotransmitters, endogenous mediators, and inflammatory mediators such as bradykinin, NF-κB, COX-2, PGE2, and TNF-α, TRPV1 is effectively recruited and activated [29–32]. Building upon our previous research, which demonstrated that A. officinarum flavonoids can regulate TRPV1 receptor expression to protect against gastric mucosal injury [8], it was hypothesized that through its interaction with the TRPV1 receptor, galangin inhibits the production and release of neurotransmitters, endogenous mediators, and inflammatory mediators, thereby regulating the activity of inflammatory signaling pathways. This regulatory effect can reduce nociception conduction and the release of inflammatory mediators during inflammation and exert an anti-inflammatory analgesic effect. To test this, a rigorous inflammatory pain assay was designed using capsaicin, a selective TRPV1 activator. The results obtained were consistent with those previously observed in the complete freund’s adjuvant model.
In comparison to the control group, both galangin and capsazepine significantly reduced the duration of lick stress, indicating that galangin modulates inflammatory signaling pathways by inhibiting the production and release of neurotransmitters, endogenous mediators, and inflammatory mediators through its interaction with the TRPV1 receptor. This regulatory effect reduces nociception conduction and inflammatory mediator release during inflammation, exerting an anti-inflammatory analgesic effect. These findings are consistent with previous studies conducted using capsaicin-induced pain models [33–35].
In a previous study, galangin was found to suppress the levels of inflammatory cytokines in the serum of mice induced by capsaicin, suggesting that galangin may modulate the TRPV1 receptor pathway to alleviate inflammatory pain [36]. Our experimental results support the findings of the previous authors. However, unlike previous studies, our research delved further into the underlying molecular mechanisms of galangin’s anti-inflammatory effects on inflammatory pain.
The molecular affinity of galangin to TRPV1, NF-κB, COX-2, and TNF-α was analyzed using molecular docking. Galangin binds strongly to these targets, reducing pain through TRPV1 binding, and inhibiting NF-κB activation to reduce inflammation and immune responses. It also inhibits COX-2 activity, reducing inflammation and pain transmission. Additionally, it regulates TNF-α-mediated inflammation and immune responses. In addition, an interaction network of TRPV1, COX-2, and TNF-α proteins was constructed using the string database (https://cn.string-db.org/) and it was observed that TRPV1 protein may serve as a central signaling molecule among these related factors (Fig. 7E). Building upon these findings, the expression of associated proteins and mRNA in rat dorsal root ganglion tissues was further verified using qPCR and Western blot analysis. These results demonstrate that galangin possesses the ability to suppress the expression of TRPV1 protein induced by capsaicin. Furthermore, galangin has been shown to inhibit the expression of TRPV1, CGRP, SP, TNF-α, COX-2, and PGE2 mRNA in rat dorsal root ganglion tissues.
In conclusion, our findings suggest that galangin possesses the ability to alleviate inflammation and pain induced by chemical or physical stimuli. Furthermore, it exhibits a potent inhibitory effect on neurogenic pain triggered by capsaicin. Based on these results, it is hypothesized that galangin exerts its anti-inflammatory and analgesic effects by modulating the transmission of inflammatory pain mediated through the TRPV1 receptor. This modulation leads to the suppression of neurotransmitters, NF-κB/TNF-α, COX-2, and other endogenous substances, ultimately resulting in the alleviation of inflammatory pain. These findings offer a unique perspective on the therapeutic potential of galangin in the management of inflammatory pain-related conditions. However, it is important to note that the current evidence only supports galangin's potential as a therapeutic agent targeting TRPV1 modulation for inflammatory pain. The therapeutic role and mechanisms of galangin in other types of pain require further investigation in future studies.
Go to :
 XML Download
XML Download