

 PDF
PDF Citation
Citation Print
Print



Highlights
· OCM is driven by the folate cycle, the Hcy-methionine cycle, and the transsulfuration pathway.
· Low OCM nutrients result in fewer methyl groups, HHcy, and less antioxidation.
· HHcy and reduced antioxidation lead to oxidative stress and systemic inflammation.
· Low methyl group supply modifies DNA methylation of genes in glycolipid metabolism.
· These changes may cause β-cell dysfunction, IR, glucose intolerance, and DM development.
INTRODUCTION
Diabetes mellitus (DM) affects approximately 9.3% (463 million people) of the population worldwide in 2019 with its prevalence projected to reach 10.2% (578 million) by 2030 and 10.9% (700 million) by 2045 [1]. DM is associated with increased risk of numerous chronic diseases, such as cardiovascular diseases (CVD) and diabetic retinopathy [2,3]. Thus, identifying potentially modifiable risk factors for DM may help develop more effective strategies for its prevention.
Hyperhomocysteinemia (HHcy) has emerged as a risk biomarker for type 2 diabetes mellitus (T2DM) [4]. HHcy was evidenced to promote oxidative stress, β-cell dysfunction, and insulin resistance (IR), which contributes to DM pathology [4-6]. Growing evidence suggests that HHcy can be due to the low status of one-carbon metabolism (OCM) nutrients [7-9]. OCM is a metabolic network that involves biochemical compounds to regulate nucleic acid synthesis and methylation reactions. Homocysteine (Hcy) in this OCM network can be either metabolized into cysteine or recycled into methionine with the aid of a group of OCM nutrients, which act as prerequisite substrate donors (folate, choline, betaine, and methionine) or essential coenzymes (vitamin B2, B6, B12, and zinc) [8,9]. Data directly linking the intake or circulating levels of these OCM nutrients to incident DM are sparse and results remain inconsistent [10-22]. For example, higher choline intake was associated with lower T2DM risk among men in eastern Finland [14], whereas dietary choline or betaine intake was not associated with risk of T2DM among the United States Black or White men [15]. Moreover, the etiology of HHcy may involve genetic variation encoding key enzymes in OCM, which may contribute to DM development [5,6]. A meta-analysis incorporating 68 studies indicated that methylenetetrahydrofolate reductase (MTHFR) 677C>T polymorphism was correlated with T2DM in Asian populations, but not in White and Black populations [8]. Nevertheless, knowledge regarding the role of OCM in DM development remains in its infancy. Further investigations are warranted to better understand the role of OCM nutrients in the etiology of DM prevention and treatment.
The present review aims to provide an overview of the existing literature assessing the relationships between OCM nutrients status as well as related genetic variation and risk of DM in the context of DM pathology. We will focus on understanding the hypothesized mechanism of OCM nutrients action on DM, limitations of the previous studies, and implications for dietetic practice and future research.
ONE-CARBON METABOLISM
OCM is a metabolic network driven by three interrelated metabolic pathways, which include the folate cycle, the Hcy-methionine cycle, and the transsulfuration pathway [2]. The complex set of biochemical reactions of OCM contributes to the generation or utilization of methyl groups (CH3) [2,3].
As shown in Fig. 1, folic acid (FA) from dietary intake can serve as a precursor to dihydrofolate (DHF) that is converted to tetrahydrofolate (THF) via dihydrofolate reductase (DHFR). DHF can also be converted to 5,10-methylenetetrahydrofolate (5,10-MTHF) via thymidylate synthase (TS), which is used for thymidylate synthesis. Conversion to 5,10-MTHF from THF requires serine via serine hydroxymethyltransferase (SHMT) with vitamin B6 as a cofactor. Methylenetetrahydrofolate dehydrogenase (MTHFD) catalyzes 5,10-MTHF to 10-formyltetrahydrofolate (10-formyl THF), which is used for purine synthesis. Conversion from 5,10-MTHF to 5-methyltetrahydrofolate (5-MTHF) requires MTHFR with vitamin B2 as a cofactor. Folate, the natural form from diet, can also contribute to 5-MTHF donating the methyl groups to Hcy to generate methionine and THF via methionine synthase (MS) with vitamin B12 and zinc as a cofactor [2-6].
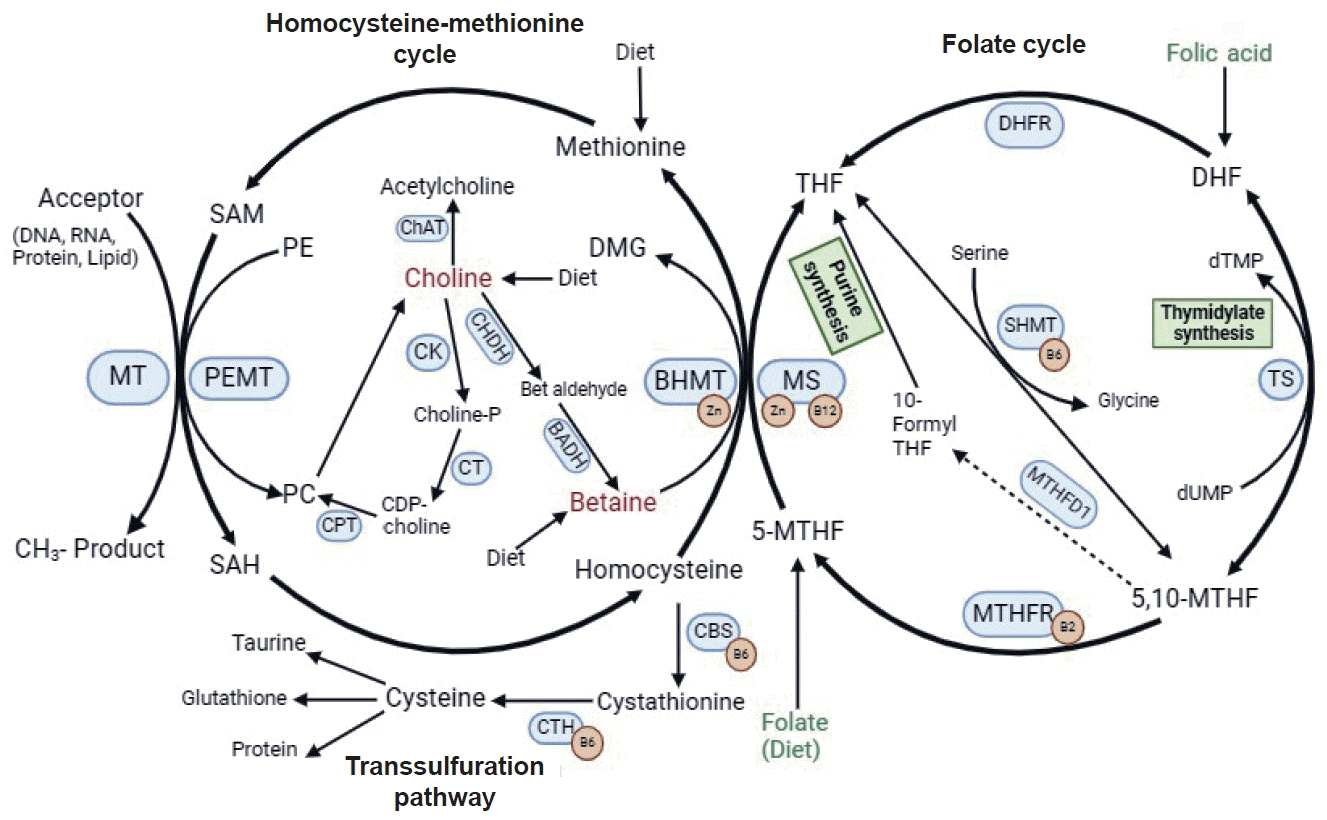
Fig. 1.
One-carbon metabolism. SAM, S-adenosylmethionine; PE, phosphatidylethanolamine; ChAT, choline acetyltransferase; DMG, dimethylglycine; MT, methyltransferase; PEMT, phosphatidylethanolamine N-methyltransferase; CK, choline kinase; CHDH, choline dehydrogenase; BHMT, betaine-homocysteine S-methyltransferase; Zn, zinc; SAH, S-adenosylhomocysteine; PC, phosphatidylcholine; CPT, cholinephosphotransferase; CDP, cytidine diphosphate; Choline-P, phosphocholine; CT, cytidylyltransferase; BADH, betaine aldehyde dehydrogenase; CBS, cystathionine β-synthase; CTH, cystathionine γ-lyase; THF, tetrahydrofolate; DHFR, dihydrofolate reductase; DHF, dihydrofolate; MS, methionine synthase; 10-formyl THF, 10-formyl-tetrahydrofolate; SHMT, serine hydroxymethyltransferase; dTMP, thymidine monophosphate; TS, thymidylate synthase; 5-MTHF, 5-methyl- tetrahydrofolate; MTHFR, methylenetetrahydrofolate reductase; dUMP, deoxyuridine monophosphate; 5,10-MTHF, 5,10-methylenetetrahydrofolate.
![]()
In addition, choline and betaine act as other methyl-donors that can be supplied from diet and endogenously synthesized. Choline as an essential nutrient can be acetylated by choline acetyltransferase to produce acetylcholine, a pivotal neurotransmitter [4]. Also, choline can be phosphorylated by choline kinase to be converted to phosphocholine, and then to cytidine diphosphate-choline and phosphatidylcholine (PC) by cytidylyltransferase (CT) and cholinephosphotransferase, sequentially. PC serves as an essential constituent of cell and mitochondrial membranes as well as a major component of lipoproteins [2-4]. In addition, choline can be endogenously synthesized through the conversion of phosphatidylethanolamine to PC, which is catalyzed by phosphatidylethanolamine N-methyltransferase (PEMT) [3]. Moreover, choline dehydrogenase catalyzes choline to synthesize betaine that can be oxidized by betaine-homocysteine S-methyltransferase with zinc as a cofactor to methylate Hcy to methionine. Then, methionine passes the methyl group to S-adenosylmethionine (SAM), which serves as a universal methyl-donor contributing to methylation modification of DNA, RNA, and protein [2-4]; After that, SAM is converted to S-adenosylhomocysteine and deadenosylated to produce Hcy [20]. If there is abundant methionine, the transsulfuration pathway will become active, by which Hcy reacts with serine to form cystathionine by cystathionine β-synthase with vitamin B6 as a cofactor. Cystathionine is further processed by cystathionine γ-lyase with vitamin B6 as a cofactor to generate cysteine, which is used to produce taurine, glutathione (GSH), and other protein [2-4]. It has been evidenced that deficiency of the OCM nutrients (e.g., folate, choline, betaine, vitamin B6 and B12) and reduced activity of the key enzymes in OCM due to genetic variation (e.g., MS 2756 G>A, MTHFR 677C>T) contribute to HHcy, thus impairing the remethylation and/or transsulfuration pathways [5-9,20].
OCM NUTRIENTS AND RISK OF DM
Observational studies that directly related OCM nutrients to incident DM were limited and results remained contradictory (Table 1). One cross-sectional study reported an inverse relationship between serum folate level and DM prevalence among Chinese adults [20]. Consistently, a cross-sectional study demonstrated that serum choline or betaine was inversely correlated with fasting glucose levels and IR index in Canadian adults [18]. Another cross-sectional study conducted in Chinese adults showed that higher dietary consumptions of vitamin B6 and choline, but not folate, vitamin B12, methionine and betaine, were related to lower incident hyperglycemia [21]. However, a positive correlation between dietary methionine and the rate of DM was observed among Chinese adults [22].
Table 1.
Observational studies examining association of one-carbon metabolism nutrients status with risk of DM
| Study | Year | Location | Study population | Design | Exposure | Follow-up | Outcomes/Major results |
|---|---|---|---|---|---|---|---|
| Al-Maskari et al. [10] | 2012 | Oman | 100 Omani adults aged 42.92–59.33 years (n=50 diabetic cases, n=50 healthy controls) | Case-control | Dietary intakes and serum levels of folate and vitamin B12 | NA | Bothe dietary intakes and serum levels of folate and vitamin B12 were lower in patients with T2DM than those in the healthy controls (all P<0.05). |
| Hong et al. [11] | 2017 | South Korea | 7,333 Adults aged ≥40 years | Prospective | Dietary folate intake (not including folate intake from supplements) | 4.06 years | Dietary folate intake was inversely associated with risk of T2DM for women (RR in the highest vs. the lowest quartile was 0.64 [95% CI, 0.43–0.95; Ptrend =0.0244]). |
| Eshak et al. [12] | 2019 | Japan | 19,168 Healthy adults aged 40–79 years | Prospective | Intake of water-soluble vitamins | 5 years | Higher dietary intakes of folate and vitamin B2 were associated with lower risk of T2DM in Japanese women (OR in the highest vs. the lowest quartile of intakes were 0.70 [95% CI, 0.46–0.98; Ptrend =0.03] for folate and 0.56 [95% CI, 0.34–0.93; Ptrend =0.03] for vitamin B2). |
| Zhu et al. [13] | 2020 | USA | 4,704 White and Black adults aged 18–30 years | Prospective | Total intake of folate, vitamin B6, and vitamin B12 | 30 years | Intake of folate, but not vitamin B6, or vitamin B12, was inversely associated with DM incidence (HR in the highest vs. the lowest quartile of folate intake was 0.70 [95% CI, 0.51–0.97; Ptrend =0.02]). |
| Virtanen et al. [14] | 2020 | Finland | 2,332 Men aged 42–60 years | Prospective | Choline intake | 19.3 years | Higher choline intake was associated with lower risk of T2DM among men in eastern Finland (HR in the highest vs. the lowest quartile of choline intake was 0.75 [95% CI, 0.57–0.98; Ptrend =0.02]). |
| Dibaba et al. [15] | 2020 | USA | 13,440 Men and women aged 45–64 years | Prospective | Intake of choline and betaine | 9 years | Overall and among male participants, neither dietary choline nor betaine intake was associated with risk of T2DM. |
| Among women, there was a trend for a modestly higher T2DM risk (HR in the highest vs. the lowest quartile of choline intake was 1.54 [95% CI, 1.06–2.25; Pinteraction for sex=0.07]). | |||||||
| Greenberg et al. [16] | 2021 | USA | 46,263 Postmenopausal women aged 50–79 years | Prospective | Intake of choline and betaine | 13.3 years | Higher choline intake was associated with increase in DM risk (HR in the highest vs. the lowest quartile of choline intake was 1.30 [95% CI, 1.15–1.47; Ptrend <0.0001]). |
| There was a significant linear trend but no significant association between betaine intake and risk of DM (HR in the highest vs. the lowest quartile of betaine intake was 0.90 [95% CI, 0.81–1.002; Ptrend =0.04]). | |||||||
| Lu et al. [17] | 2022 | China | 1,565 Chinese adults aged 40–75 years | Prospective | Serum betaine | 8.9 years | Higher serum betaine was associated with lower risk of T2DM (HR in the highest vs. the lowest quartile was 0.46 [95% CI, 0.31–0.69; Ptrend <0.001]). |
| Gao et al. [18] | 2019 | Canada | 1,081 Canadian adults aged 28–56 years | Cross-sectional | Serum choline and betaine levels | NA | Serum choline level was negatively correlated with serum fasting glucose levels in males (R=–0.121, P=0.006). |
| Serum betaine level was negatively associated with insulin levels (R=−0.081, P=0.043), and HOMA-IR index (R=−0.086, P=0.021) in males. In females, serum betaine level was negatively associated with insulin levels (R=−0.104, P=0.016), and with HOMA-IR index (R=−0.092, P=0.034). | |||||||
| Nie et al. [19] | 2022 | China | 188 Chinese adults aged 57–72 years (n=94 diabetic cases, n=94 healthy controls) | Case-control | Plasma metal levels | NA | Plasma Zn level was positively correlated with elevated DM risk (OR in the highest vs. the lowest quartile of plasma Zn levels was 2.37 [95% CI, 1.47–3.81; P<0.001]). |
| Cheng et al. [20] | 2022 | China | 772 Adults aged 43–82 years (n=370 T2DM cases, n=402 controls) | Cross-sectional | Serum folate | NA | Higher serum folate level was associated with lower risk of T2DM (OR in the highest vs. the lowest quartile of serum folate levels was 0.909 [95% CI, 0.840–0.983; Ptrend =0.0177]). |
| Ding et al. [21] | 2022 | China | 901 Participants (n=417 males, n=484 females) aged 18–75 years | Cross-sectional | Dietary intake of betaine, total choline, methionine, folate, vitamins B6 and B12 | NA | Higher dietary intakes of total choline and vitamin B6 were associated with a lower incidence of hyperglycemia (OR in the highest vs. the lowest quartile were 0.601 [95% CI, 0.365–0.988; Ptrend =0.365–0.988] for total choline; 0.575 [95% CI, 0.346–0.956; Ptrend =0.038] for vitamin B6). |
| There were null associations of dietary intakes of folate, vitamin B12, methionine, and betaine with risk of hyperglycemia. | |||||||
| Sun et al. [22] | 2023 | China | 12,489 Chinese adults aged ≥20 years | Cross-sectional | Methionine intake | NA | Higher intake of dietary methionine was associated with higher risk of DM (OR in the highest vs. the lowest quartile was 1.49 [95% CI, 1.12– 1.98; Ptrend =0.009]). |
![]()
Two case-control studies investigated the relationship between OCM nutrients and DM development. Al-Maskari et al. [10] observed that both dietary intake and serum level of folate and vitamin B12 were lower in Omani adult patients with T2DM, compared to the healthy controls. However, Nie et al. [19] reported a positive correlation between plasma zinc and the odds of DM in Chinese adults.
The inverse associations were reported between dietary folate intake and the rate of DM among Korean women aged ≥40 years with an average 4-year follow-up [11], and in Japanese women aged 40 to 79 years within a 5-year study period [12]. Similarly, our previous study found that higher intake of folate, but not vitamin B6 or vitamin B12, in young adulthood was associated with lower diabetes incidence in midlife among White and Black Americans over 30 years of follow-up [13]. Regarding other B vitamins in OCM, dietary vitamin B2 intake was inversely associated with risk of T2DM in Japanese women aged 40 to 79 years [12]. In addition, higher choline intake was inversely associated with lower T2DM risk among men aged 42 to 60 years in eastern Finland [14], while dietary choline or betaine intake was not significantly associated with risk of T2DM among the United States Black or White male participants aged 45 to 64 years [15]. In contrast, a study reported a positive trend for the association between choline consumption and DM risk among postmenopausal United States women aged 50 to 79 years [15]. As for the biomarkers of OCM nutrients, higher serum betaine was associated with lower T2DM risk in Chinese adults aged 40 to 75 years [17]. Presumably, the inconsistent findings from the aforementioned observational studies are mainly due to heterogeneities in exposure measures, time windows of exposure, and diverse study populations.
Randomized clinical trials (RCT) investigating the effects of OCM nutrients supplementation on incident DM were scanty (Table 2). A double-blind RCT reported that Hcy-lowering intervention by daily B vitamins supplementation (FA [2.5 mg], vitamin B6 [50 mg], and vitamin B12 [1 mg]) for 4.5 years failed to decrease incident T2DM among 5,442 United States female health professionals at high risk for CVD [7]. Likewise, another double-blind RCT reported that daily supplementation of FA (0.8 mg) with enalapril (10 mg) for 7.3 years exerted no significant impact on risk of new-onset diabetes among 20,702 Chinese hypertensive adults [8]. Notably, a double-blind RCT involving 200 Sri Lankan participants with prediabetes had noted that 12-month zinc supplementation (20 mg zinc daily) decreased T2DM incidence [9]. These discrepant results are probably explained by different supplementation formulas and duration, and various health statuses across different study populations. Moreover, the participants in these studies were those who were >40 years old and at high risk for metabolic disorders. Thus, the generalizability of these findings is limited.
Table 2.
Intervention studies examining the effect of one-carbon metabolism nutrients on DM development and glucose metabolism index
| Study | Year | Location | Study population | Design | Intervention | Duration | Outcomes/Major results |
|---|---|---|---|---|---|---|---|
| DM risk | |||||||
| Song et al. [7] | 2009 | USA | 5,442 Female health professionals aged 54–72 years with history of CVD or ≥3 CVD risk factors | Double-blind RCT | Treatment group: daily intake of a combination pill containing 2.5 mg FA, 50 mg vitamin B6, and 1 mg vitamin B12 (n=2,132) | 7.3 years | No significant effect on risk of DM |
| Placebo group: a matching placebo pill daily (n=2,120) | ↓Blood Hcy | ||||||
| Qin et al. [8] | 2016 | China | 20,702 Hypertensive patients aged 52.3–57.5 years | Double-blind RCT | Daily treatment with tablets containing: (1) 10 mg enalapril and 0.8 mg FA (n=10,348); or (2) 10 mg enalapril alone (n=10,354) | 4.5 years | No significant effect on FPG and risk of DM |
| Ranasinghe et al. [9] | 2018 | Sri Lanka | 200 Participants with prediabetes aged 44–59.4 years | Double-blind RCT | Treatment group: capsules containing 20 mg zinc daily (n=100) | 12 months | ↓FPG, ↓2-hour glucose after the OGTT, ↓HOMA-IR, ↓T2DM development |
| Placebo group: capsules containing inactive ingredients (n=100) | ↑HOMA-β | ||||||
| Glucose metabolism index | |||||||
| Aarsand et al. [37] | 1998 | Norway | 28 Diabetic patients with metformin treatment for more than 1 year, aged 53.9–64.1 years | Double-blind RCT | Treatment group: tablets containing 0.25 mg folate+60 mg Fe2+ daily (n=14) | 12 weeks | No significant effect on FSG and HbA1c |
| ↓Serum Hcy | |||||||
| Placebo group: tablets containing 60 mg Fe2+ daily (n=14) | ↑Serum vitamin B12 and folate | ||||||
| Doshi et al. [23] | 2001 | UK | 50 Patients with coronary artery disease, aged 46–65 years | A randomized, double-blind, placebo-controlled crossover trial | Treatment group: 5 mg FA supplement per day | Two 6-week treatments separated by a washout period of 4 months | No significant effect on FPG |
| Placebo group: matched placebo | ↑Plasma folate | ||||||
| ↓Plasma Hcy | |||||||
| Doshi et al. [24] | 2002 | UK | 33 Patients with coronary artery disease, aged 46–65 years | Double-blind RCT | Treatment group: 5 mg FA supplement per day (n=16) | 6 weeks | No significant effect on FPG |
| Placebo group: matched placebo (n=17) | ↑Plasma folate | ||||||
| ↓Plasma Hcy | |||||||
| Setola et al. [38] | 2004 | Italy | 50 Patients with metabolic syndrome+hyperinsulinemia, aged 66.1–68.5 years | Double-blind, parallel, identical placebo-drug RCT | Group 1: treated with diet+placebo for 2 months | 2 months | ↓Insulin levels, ↓HOMA-IR, ↓Hcy levels |
| Group 2: treated with diet+placebo (1 month) then diet+FA (5 mg/day)+vitamin B12 (500 µg/day) for 2nd month | No significant effect on fasting glucose | ||||||
| Kilicdag et al. [48] | 2005 | Turkey | 49 Female patients with PCOS (group 1, 17.22–31.06 years; group 2, 18.27–25.73 years; group 3, 18.27–31.61 years) | RCT | Group 1 received metformin (850 mg twice daily) (n=20) | 12 weeks | No significant effect on HOMA-IR |
| Group 2 received metformin (850 mg twice daily)+B-group vitamins (vitamin B1 250 mg, vitamin B6 250 mg, vitamin B12 1,000 µg twice daily) (n=20) | ↓Serum Hcy (groups 1 & 2) | ||||||
| Group 3 received metformin (850 mg twice daily)+FA (174 µg), vitamin D (1,200 µg) and calcium (666.670 mg) twice daily (n=20) | ↑Plasma FA (group 3), ↑plasma vitamin B12 (group 2) | ||||||
| Mangoni et al. [25] | 2005 | Australia | 26 T2DM patients aged 46–65 years | Double-blind, parallel-group RCT | Treatment group: 5 mg FA supplement per day (n=13) | 4 weeks | No significant effect on plasma glucose, HbA1c, and serum vitamin B12 |
| Placebo group: matched placebo (n=13) | ↓Plasma Hcy, ↑serum folate | ||||||
| Sheu et al. [26] | 2005 | Taiwan | 84 Obese women who were 20% over their ideal weight | Double-blind RCT | Treatment group: 5 mg FA supplement per day (n=36) | 12 weeks | ↓HOMA-IR, ↓fasting plasma insulin, ↓serum Hcy |
| Placebo group: matched placebo (n=38) | No significant effect on FPG and fasting serum vitamin B12 and folate | ||||||
| This program also included caloric restriction and light exercises to promote weight loss. | |||||||
| Villa et al. [27] | 2005 | Italy | 20 Healthy postmenopausal women aged 48–61 years | RCT | Treatment group: 7.5 mg FA supplement per day (n=10) | 8 weeks | No significant effect on fasting glucose and insulin, and vitamin B12, methionine |
| Placebo group: commercial calcium capsule (n=10) | ↓Hcy levels | ||||||
| Moat et al. [28] | 2006 | USA | 128 Patients with angiographically proven CAD aged 53–68 years | Double-blind, parallel, RCT | 84 Patients were randomly divided into 3 groups for FA study (placebo, n=29; low-dose FA 400 µg/day, n=30; and high-dose FA 5 mg/day, n=25). Parallelly 44 patients were randomly divided into 2 groups for betaine study (Placebo, n=23 or betaine, 100 mg/kg/day, n=21). | 6 weeks | ↓Total plasma Hcy levels, ↑plasma folate, and no significant difference in blood glucose and serum vitamin B12 for FA study |
| ↓Plasma folate but no significant difference in blood glucose, serum vitamin B12, or plasma Hcy for betaine study | |||||||
| Solini et al. [29] | 2006 | Italy | 60 Healthy overweight adults aged 29–61 years | Unmasked randomized, placebo-controlled trial | All patients were put on a hypocaloric diet and were randomly assigned to either a placebo or FA (2.5 mg/day) group. | 12 weeks | ↓Fasting plasma insulin, ↓HOMA-IR, ↑serum folate |
| No significant difference in serum vitamin B12, Hcy, or FPG | |||||||
| Title et al. [30] | 2006 | Canada | 19 T2DM patients aged 35–65 years | Randomized, double-blind, placebo-controlled, crossover trial | Patients were randomly assigned to receive either oral FA (10 mg/day) or a matching placebo for 2 weeks. This was followed by 8 weeks of washout period and then patients were crossed over to receive alternate treatment for another 2 weeks. | 2 weeks+8 weeks washout+2 weeks | No significant effect on plasma Hcy and plasma glucose |
| ↑Serum folate | |||||||
| Moens et al. [31] | 2007 | Belgium | 40 Patients with acute myocardial infarction, aged 42–70 years | Randomized, double-blind, placebo-controlled crossover trial | Patients were randomly divided into 2 groups: Group A (n=20) received FA (10 mg/day) for initial 6 weeks then a placebo for another 6 weeks. For group B (n=20), the order was reversed. There was a washout of 2 weeks between the treatments. | 6 weeks+2 weeks washout+6 weeks | No significant difference in plasma vitamin B12, and FPG |
| ↑Plasma and RBC folate, ↓plasma Hcy | |||||||
| Mao et al. [50] | 2008 | China | 443 Patients with mild to moderate hypertension, aged 27–75 years | Double-blind RCT | Participants were randomly assigned to 3 groups: control (10 mg of enalapril, n=149); low-FA group (10 mg enalapril+0.4 mg of FA, n=146); high-FA group (10 mg enalapril+0.8 mg of FA, n=148). | 8 weeks | ↓FPG in low-FA and high-FA groups |
| ↑Serum folate | |||||||
| Mashavi et al. [42] | 2008 | Israel | 57 T2DM patients aged 54.1–66.1 years | Double-blind RCT | Group 1: 1,500 mg metformin+folate (1,000 µg), vitamin B12 (400 µg), and vitamin B6 (10 mg) daily (n=28) | 4 months | No significant difference in HOMA-IR and FPG |
| ↑Serum FA, ↑serum vitamin B12 | |||||||
| Group 2: 1,500 mg metformin+placebo daily (n=29) | ↓Serum Hcy | ||||||
| Potter et al. [39] | 2008 | Australia | 162 Patients with history of stroke, aged 52–80 years | Double-blind RCT | Treatment group: a single daily tablet containing FA (2 mg), vitamin B6 (25 mg), and vitamin B12 (500 µg) (n=83) | 104 weeks | No significant difference in FGB and HbA1c |
| ↓Blood Hcy | |||||||
| Placebo group: matched placebo (n=79) | ↑Serum B6, ↑serum vitamin B12, ↑RBC folate | ||||||
| Cagnacci et al. [32] | 2009 | Italy | 30 Healthy White postmenopausal women aged 48–58 years | Double-blind RCT | Treatment group: 15 mg/day 5-methyltetrahy-drofolate (n=15) | 3 weeks | ↓HOMA-IR, ↓blood insulin, ↓blood Hcy |
| Placebo group: matched placebo (n=15) | No significant effect on FBP | ||||||
| Palomba et al. [41] | 2010 | Italy | 47 Females with PCOS, aged 23.6–30 years | Non-randomized placebo-controlled double-blind trial | Experimental group: 1,700 mg metformin+400 µg FA daily (n=25) | 25 weeks | ↓Fasting serum insulin, ↓HOMA-IR |
| Control group: 1,700 mg metformin+a placebo daily (n=25) | ↑Serum Hcy | ||||||
| No significant difference in serum folate, vitamin B12, or FBG | |||||||
| Kurt et al. [40] | 2010 | Turkey | 44 Adults aged >65 years with vitamin B12 deficiency | Double-blind RCT | Treatment group: FA (5 mg)+vitamin B12 (500 µg) daily (n=24) | 8 weeks | ↓HOMA-IR, ↓serum Hcy |
| Placebo group: matched placebo (n=20) | ↑Serum folate, ↑serum vitamin B12 | ||||||
| No significant difference in FPG | |||||||
| Gargari et al. [33] | 2011 | Iran | 48 Overweight and obese men with T2DM and under metformin treatment, aged 46.9–67.7 years | Double-blind RCT | Treatment group: FA (5 mg) supplementation daily (n=24) | 8 weeks | ↓HOMA-IR, ↓serum HbA1c, ↓serum insulin |
| Placebo group: matched placebo (n=24) | ↑Serum folate, ↑serum vitamin B12 | ||||||
| No significant difference in FBG | |||||||
| Grigoletti et al. [34] | 2013 | Brazil | 30 HIV-infected individuals aged 43–47 years | Double-blind RCT | Treatment group: FA (5 mg) supplementation daily (n=15) | 4 weeks | No significant difference in FSG |
| Placebo group: matched placebo (n=15) | ↑Serum vitamin B12, ↑serum folate | ||||||
| ↓Plasma Hcy | |||||||
| Asemi et al. [35] | 2014 | Iran | 81 Overweight or obese women with PCOS aged 18–40 years | Double-blind RCT | Group 1: 1 mg FA daily (n=27) | 8 weeks | ↓Serum insulin, ↓HOMA-IR, ↓plasma Hcy |
| Group 2: 5 mg FA daily (n=27) | No significant difference in FPG | ||||||
| Group 3: matched placebo (n=27) | |||||||
| Karamali et al. [45] | 2015 | Iran | 58 Females with GDM aged 18–40 years | Double-blind RCT | Treatment group: 233 mg zinc gluconate daily (n=29) | 6 weeks | ↓FPG, ↓serum insulin, ↓HOMA-IR, ↓HOMA-β |
| Placebo group: matched placebo (n=29) | ↑QUICKI, ↑serum zinc | ||||||
| Asemi et al. [49] | 2016 | Iran | 58 Females with cervical intraepithelial neoplasia grade 1, aged 18–55 years | Double-blinded RCT | Treatment group: 5 mg FA daily (n=29) | 6 weeks | ↓Serum insulin levels, ↓HOMA-β, ↓plasma Hcy |
| Placebo group: matched placebo (n=29) | |||||||
| Hashemi et al. [36] | 2016 | Iran | 79 Pregnant women with preeclampsia, aged 21–41 years | Randomized, triple-blind, clinical trial | Treatment group: 5 mg FA daily | 8 weeks | No significant difference in FBG |
| Placebo group: matched placebo | |||||||
| Talari et al. [43] | 2016 | Iran | 60 Patients with metabolic syndrome, aged 40–85 years | Double-blind RCT | Treatment group: tablets containing 5 mg FA daily (n=30) | 12 weeks | ↓FPG, ↓serum insulin, ↓HOMA-IR, ↓plasma Hcy |
| Placebo group: daily placebo tablets (n=30) | No significant difference in HOMA-β | ||||||
| Bahmani et al. [44] | 2018 | Iran | 60 Women with endometrial hyperplasia | Double-blind RCT | Treatment group: 5 mg FA daily (n=30) | 12 weeks | ↓FPG, ↓serum insulin, ↓HOMA-IR, ↑QUICKI |
| Placebo group: matched placebo (n=30) | |||||||
| Attia et al. [46] | 2022 | Australia | 98 Prediabetic participants aged 40–70 years | Double-blind RCT | Treatment group: a daily capsule containing 30 mg elemental zinc gluconate (n=48) | 12 months | No significant difference in FBG, HbA1c, and HOMA-β |
| Placebo group: a daily capsule containing cellulose (n=50) | |||||||
DM, diabetes mellitus; CVD, cardiovascular diseases; RCT, a randomized placebo-controlled trial; FA, folic acid; Hcy, homocysteine; FBG, fasting blood glucose; FPG, fasting plasma glucose; OGTT, oral glucose tolerance test; HOMA-IR, homeostasis model assessment of insulin resistance; T2DM, type 2 diabetes mellitus; HOMA-β, homeostatic model assessment of β-cell function; FSG, fasting serum glucose; HbA1c, glycosylated hemoglobin; PCOS, polycystic ovarian syndrome; CAD, coronary artery disease; RBC, red blood cell; FGB, fibrinogen beta chain; FBP, folate-binding protein; HIV, human immunodeficiency virus; GDM, gestational diabetes mellitus; QUICKI, quantitative insulin sensitivity check index.
![]()
EFFECT OF OCM NUTRIENTS SUPPLEMENTATION ON GLUCOSE METABOLISM INDICES
Although it is insufficient to draw a firm conclusion as to whether OCM nutrients can prevent incident DM, a growing body of interventional trials have demonstrated beneficial effects of OCM nutrients consumption on glucose metabolism indices (Table 2).
Fasting blood glucose (FBG) is a common glucose metabolism index and the easiest way to monitor blood glucose levels for DM diagnosis. The present review identified 26 previous studies that explored the effects of OCM nutrients supplementation on FBG and the results remain elusive (Table 2). Single administration of FA (400 µg/day to 15 mg/day), betaine (100 mg/kg/day), and zinc (30 mg/day) failed to result in significant changes in FBG values [23-36]. In addition, no remarkable alterations in FBG values were reported following the joint supplementation of FA with Fe2+ [37], FA with other B vitamins [38-40], FA with metformin [41], B vitamins complex with metformin [42]. However, FA (5 mg/day) administration alone or joint supplementation of FA (0.4 or 0.8 mg/day) and enalapril (10 mg/day) decreased fasting plasma glucose (FPG) among Iranian patients with metabolic syndrome (MetS) [43] and Iranian women with endometrial hyperplasia [44]. Likewise, administration of zinc (20 or 233 mg/day) reduced FPG among Sri Lankan participants with prediabetes [9] and Iranian women with gestational diabetes [45].
Glycosylated hemoglobin (HbA1c) is commonly assayed to indicate average blood glucose level over the past 3 months, which is also used for DM diagnosis [25,37,39,46]. Among five studies identified in the present review, only one reported that FA (5 mg/day) supplementation led to serum HbA1c reduction in overweight and obese Iranian men with T2DM [33]. No significant changes in HbA1c values were observed following administration of FA in T2DM patients [25], supplementation of zinc in prediabetic patients [46], the joint supplementation of FA with Fe2+ [37] in diabetic patients, or FA administration with other B vitamins in patients with history of stroke [39].
The homeostasis model assessment of insulin resistance (HOMA-IR) value is calculated by an equation derived from FBG and insulin levels. Higher HOMA-IR was independently associated with an increased DM risk [47]. Single FA administrations at high doses (ranging from 2.5 to 15 mg/day) were inversely associated with HOMA-IR among obese women in Taiwan [26] and overweight adults [29] as well as postmenopausal women in Italy [32]. Similar inverse associations were reported in overweight and obese men with T2DM [33], overweight or obese women with polycystic ovarian syndrome (PCOS) [35], men and women with MetS [43], and women with endometrial hyperplasia in Iran [44]. Likewise, a single administration of zinc (20 mg/day or 233 mg zinc gluconate/ day) reduced HOMA-IR in prediabetic patients [9] and women with gestational diabetes mellitus (GDM) [45]. In addition, concomitant administration of FA (5 mg/day) with vitamin B12 (500 µg/day) or FA (400 µg/day) with metformin (1,700 mg/day) decreased HOMA-IR in patients with MetS and hyperinsulinemia [38], and elderly adults with vitamin B12 deficiency [40]. However, two other studies reported no effect of joint supplementation of B vitamins with metformin on HOMA-IR among Turkish women with PCOS [48] and Israeli patients with T2DM [42].
The quantitative insulin sensitivity check index (QUICKI) is another surrogate biomarker of IR, the higher of which reflects a lower degree of IR [44,45]. In the study by Karamali et al. [45], 6-week zinc supplementation (233 mg/day zinc gluconate) increased QUICKI in women with GDM. Likewise, Bahmani et al. [44] demonstrated that 6-week supplemental FA at 5 mg/day augmented QUICKI in women with endometrial hyperplasia.
Homeostatic model assessment of β-cell function (HOMA-β), derived from fasting plasma insulin and glucose levels, is applied as an index of the insulin secretory function of pancreatic β-cells [47]. Lower HOMA-β was independently associated with an increased DM risk [47]. Increasement of HOMA-β were observed under administration of zinc (20 mg/day) in prediabetic patients [9], whereas zinc (233 mg zinc gluconate/day) or FA (5 mg/day) supplementation decreased HOMA-β among women with GDM [45] or women with cervical intraepithelial neoplasia [49]. However, no significant effect on HOMA-β was found under single supplementation of FA (5 mg/day) in patients with MetS or zinc (30 mg zinc gluconate/day) in prediabetic patients [46].
The heterogeneous effects of either the single or the combined supplementation of OCM nutrients on the above glucose metabolism indices are due to the various dosages and combinations of the OCM nutrients with various durations [23-46,48-50]. In addition, the medication (e.g., enalapril, metformin) that was co-ingested with the OCM nutrients may also counterbalance the beneficial impact of the OCM nutrients on the glucose metabolism indices [41,42,50]. Moreover, most of the studies were conducted among patients with various health problems which contributed to the disputed results [23-46,48-50]. Whether the OCM nutrients intake prevents the public at an early age from developing prediabetic status or incident DM later in life merits further investigation. Furthermore, interventions aimed at optimizing balanced OCM nutrients status and preventing HHcy may help mitigate IR and improve insulin signaling and glucose homeostasis. While further research is warranted, incorporating OCM nutrients into holistic lifestyle strategies may provide a valuable avenue for DM management and prevention.
GENETIC VARIATION ENCODING KEY ENZYMES OF OCM IN RELATION TO RISK OF DM
Emerging evidence indicates that the etiology of HHcy may also involve genetic variation encoding key enzymes of OCM, which may contribute to diabetes development [6,13]. The transformation of the methyl group from these OCM nutrients to Hcy is catalyzed by an array of key enzymes including MTHFR, methionine synthase reductase (MTRR), and MS [51]. Genetic single nucleotide polymorphisms (SNPs) may alter these key enzymes activities, thus changing the enzymes catalytic efficiencies of metabolizing OCM nutrients after dietary intake. SNPs encoding these enzymes such as MS 2756A>G, MTRR 66A>G, MTHFR 677C>T, and 1298A>C, have been evidenced to alter blood Hcy level [51-53]. Therefore, these genetic variants may also be potential genetic markers for incident diabetes development. A meta-analysis incorporating 68 studies indicated that MTHFR 677C>T polymorphism was correlated with T2DM in Asian populations, but not in White and Black populations [54]. Additionally, MTRR 66A>G polymorphism was related to T2DM risk in overweight and obese Chinese individuals [55]. Another study showed that only those Chinese adults who carry MTHFR 1793 GA+AA genotype or MTHFR 1298 AC+CC genotype appeared to have lower T2DM risk [17]. In addition, in the middle-age Han Chinese, those with the genotype CC of MTHFR 1470 A>C had a significantly higher likelihood of T2DM, whereas those with the genotype AA of MTHFD 1958G>A or carrying CT+TT of PEMT (rs4646356) had a significantly lower likelihood of T2DM [53]. Moreover, MTHFR CTCCGA haplotype (rs12121543-rs13306553-rs9651118- rs1801133-rs2274976-rs1801131) was found to be related to decreased risk of T2DM in a Chinese population, compared with CTTTGA haplotype [56]. Nevertheless, the frequencies of these SNPs appear different among people with diverse ethnicities, which may partially explain the inconsistent results from previous studies that a direct relationship between these SNPs and diabetes remains controversial among different study populations. However, previous studies investigating the interplay between OCM nutrients status and genetic variation encoding key enzymes of OCM on DM risk were limited. Lu et al. [17] reported the joint effects of higher serum betaine levels (>47.82 µmol/L) and heterozygous or homozygous variants of MTHFR (G1793A, A1298C) could be found influencing risk of T2DM among Chinese adults aged 40 to 75 years. The biological relevance of these OCM nutrients and genetic variation to the efficiency of the OCM pathway and Hcy homeostasis makes it necessary to consider interrelationships of the OCM nutrients intakes/circulation levels and SNPs with DM risk.
HYPOTHESIZED MECHANISM OF ACTION
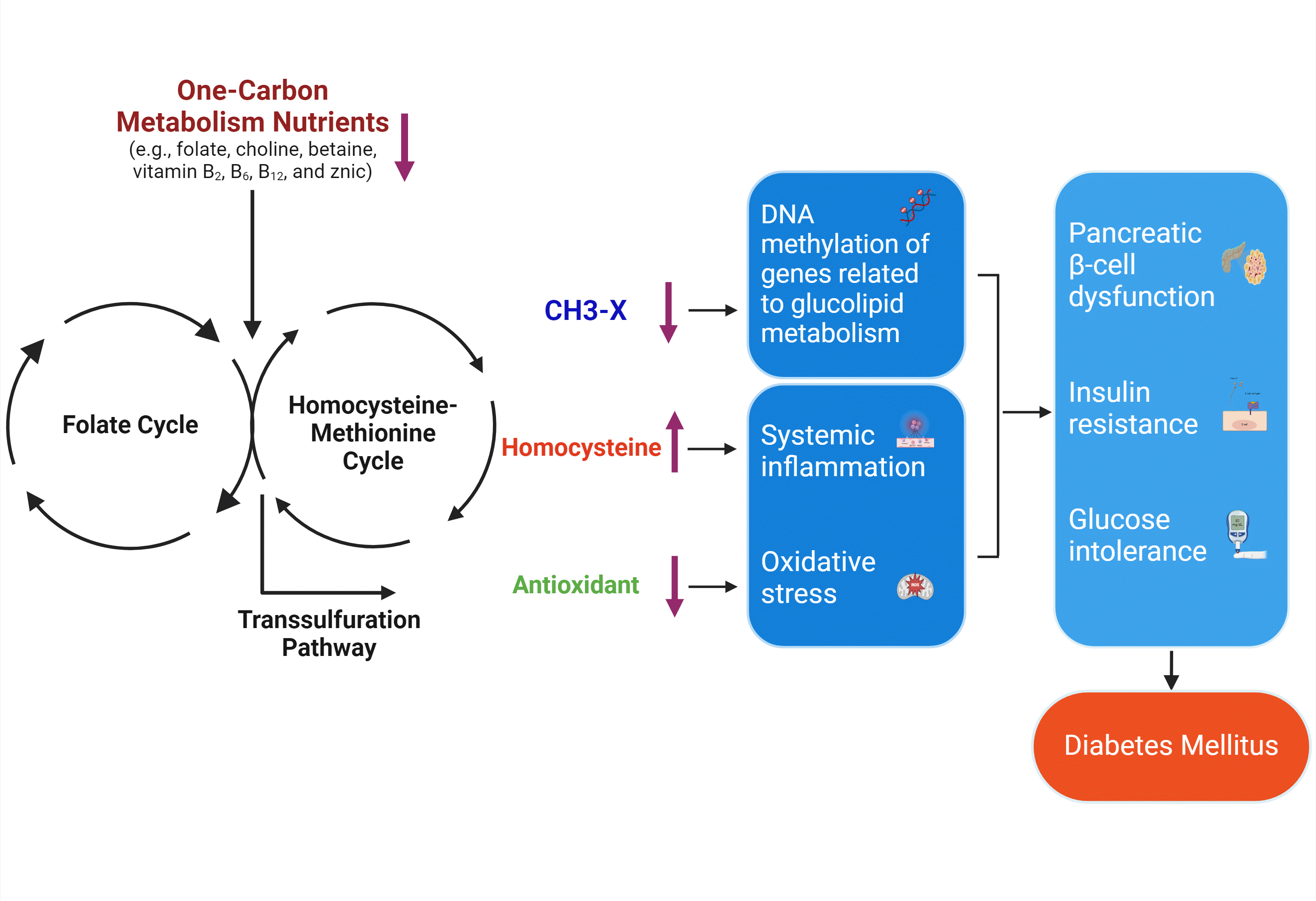
Emerging evidence from in vivo and in vitro studies suggests that OCM nutrients are essential for facilitating energy and glucose metabolism through multifactorial mechanisms. Low status of the OCM nutrients (e.g., folate, choline, or vitamin B12) has been evidenced to induce HHcy [4,10,57], which has been implicated in the pathogenesis of DM [5,13]. HHcy increases reactive oxygen species (ROS) and C-reactive protein (CRP) levels to promote oxidative stress and systemic inflammation, which have been reported to activate various stress-sensitive signaling pathways and eventually lead to pancreatic β-cells dysfunction [5,10], glucose intolerance [58], and IR [21,59,60]. OCM nutrients can directly scavenge ROS, decrease CRP, and promote production of GSH, the major intracellular antioxidant [10,21,61-64], which can counteract the disturbance in glucose metabolism by HHcy. Moreover, OCM nutrients (e.g., folate, choline, betaine) provide the methyl group to the universal methyl-donor, SAM, the change of which can modify methylation status of genetic loci signals involved in insulin signaling and glucose homeostasis [2,6,10,13,65,66]. These alterations in DNA methylation patterns may generate different gene expression profiles that facilitate the development or progression of DM [2,65,66]. However, the molecular mechanisms by which OCM nutrients contribute to DM pathology are only partially understood. The plausible mechanisms remain to be elucidated in human studies. Thus, more future studies exploring the underlying mechanisms are warranted.
IMPLICATION FOR PRACTICE
Based on the existing literature, it is premature to infer the causal relationship between OCM nutrients intake and incident DM. However, the beneficial effect of OCM nutrients on the major glucose metabolism indices indicates that consumption of natural food rich in OCM nutrients should be stimulated for maintaining optimal glucose homeostasis and preventing DM development. Co-ingestion of OCM nutrients-enriched foods, such as green leafy vegetables, legumes, fruit, nuts, whole-grain products, eggs, less-processed dairy products, and deep-sea fish, may exert a synergistic beneficial effect on better glucose control and insulin sensitivity. Common foods rich in OCM nutrients are listed in Supplementary Tables 1-7 [67-69]. In addition, there is no sufficient evidence to establish personalized OCM nutrient recommendations for DM prevention, based on the genetic variation information. Lower blood concentrations of these OCM nutrients may be indicators of a higher risk of DM. Therefore, it is worth monitoring these OCM nutrients biomarkers (e.g., plasma/serum levels of OCM nutrients) regularly to adjust their intake for optimal health.
CONCLUSIONS
The present review summarizes the existing evidence of whether OCM nutrients status influences the occurrence of DM. Currently, our knowledge of how OCM nutrients play a role in protesting against DM development in humans is in its early stages. Although limited RCTs using treatment with single/ multiple OCM nutrient(s) reported different onsets on DM, the majority of observational studies manifested that intakes or blood biomarkers of OCM nutrients, particularly folate and betaine, were inversely associated with abnormal glucose metabolism indices and affect the progression of DM. In addition, association studies suggest that several SNPs in OCM-related genes may influence the metabolic handling of methyl-donors and presumably the risk of DM. While OCM nutrients interventions displayed promise, current human studies were mainly conducted among participants with different underlying medical conditions or middle-aged and elderly populations who may have already had disease onset. Future well-designed RCTs are warranted to examine whether balanced OCM nutrients intakes at an early age prevent DM later in life among the general population. Moreover, it is essential to examine whether the OCM nutrients intakes/circulating levels interplay with genetic risk factors to influence DM development in multi-ethnic populations, which will inform the personalized recommendations for OCM nutrients intakes in terms of DM prevention and management.
 XML Download
XML Download