

 PDF
PDF Citation
Citation Print
Print



Introduction
Human joints like the hip, knee, elbow, and ankle are made up of different types of tissue such as cartilage, bone, and connective tissue. The smooth functioning of all these tissues and the interaction between them is essential for the proper working of these structures (1). The joint also contains nerve endings that innervate the synovium and subchondral bone, helping to regulate processes such as chondrocyte differentiation and bone remodeling (2) On the other hand, pathological stimulation of sensory nerves can lead to chronic pain, which is a component of diseases such as rheumatoid arthritis (RA) and osteoarthritis (OA) (3).
Because of the complex etiology of joint diseases, most studies into their development and treatment are based on animal models, particularly because the complex tissue structure of the joint is difficult to model in vitro (4). However, with the increasing pressure to reduce the use of animals in scientific experiments and due to interspecies differences, making it difficult to translate the obtained results to the clinic, there is a need to develop cellular models. In vitro models allow to use of human cells in studies, precise control of experimental variables, and screening of large numbers of compounds or potential treatments in a relatively short time. Co-culture of the two cell types allows interactions between them to be observed and a more complex biological model to be created, enabling changes in the human body to be better represented. The research in this publication used a co-culture of neurons and synoviocytes, where cells were sequentially cultured at the bottom of a well and a semi-permeable insert. This approach allowed limited physical contact between the two cell types, but it was possible for them to interact via factors secreted into the medium. In addition, in vitro neuronal differentiation would have been impossible in the presence of rapidly proliferating human fibroblast-like synoviocytes (HFLS). Previous studies have used a similar system to co-culture neurons with microglia (5) or astrocytes/endothelial cells building blood-brain barrier (6), however to the authors’ best knowledge, this is the first use of semi-permeable inserts to culture peripheral neurons with cells that build peripheral tissues to unravel the intricate mechanisms involved in joint homeostasis and disease. The ability to study secretome interactions between two cell types in a structure such as a joint, appears to be a significant improvement over standard cultures of a single cell type in a monolayer, because the synovial fluid is a tissue-specific environment and a carrier of signaling molecules (7). The objective of the research described in this manuscript was to derive a model that would allow rapid and simple analysis of the response of neuronal cells to a variety of pro-inflammatory factors secreted by HFLS, as observed in patients suffering from diseases such as OA or RA.
Materials and Methods
Neuroepithelial stem differentiation to peripheral-like neurons
Neuroepithelial stem cells (NESCs) were cultured as previously described (8). Briefly, culture flasks and plates were coated with laminin (#L2020; Sigma-Aldrich) and poly-L-ornithine (#P3655; Sigma-Aldrich) prior to cell seeding. NESCs were maintained at 37℃, 5% CO2 and cultured in DMEM/F-12+GlutaMAX (Gibco) medium supplemented with 1% N2 (Gibco), 0.1% B27 (Gibco), 1% penicillin-streptomycin mixture (Gibco), 10 ng/ml bFGF (#AF-100-18B; PeproTech), and EGF (#AF-100-15; PeproTech). For differentiation, cells were seeded at 40,000 cells/cm2 and in the day-1 differentiation medium was applied. The protocol for differentiation into peripheral-like neurons (PLN) was previously described (9), in detail, neurobasal medium supplemented with 1% N2, 0.1% B27 and penicillin-streptomycin, growth factors such as β-NGF (10 ng/ml, #AF-450-01; PeproTech), NT3 (10 ng/ml, #AF-450-03; PeproTech), BDNF (10 ng/ml, #AF-450-02; PeproTech) and ROCK inhibitor - Y27632 (25 μM, #1293823; Bio-Gems) was used for 4 weeks. The cells were then used for further experiments.
HFLS culture, pro-inflammatory stimulation and co-culture
HFLS were cultured as previously described (10) in RPMI 1640 Medium (Gibco), supplemented with GlutaMax, 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 10% FBS and 1% penicillin-streptomycin mixture (all reagents from Gibco) at 37℃ in a humidified atmosphere containing 5% CO2. HFLS were seeded at 10,000 cells/cm2 on 6-well plates (TPP) or 0.4 μm pore cell culture inserts (#657641; Greiner Bio-One) for mRNA sequencing and co-culture experiments respectively. When the cells evenly covered the culture surface, they were stimulated with lipopolysaccharide (LPS) or interferon-gamma (IFNγ) (4 or 24 hours for mRNA sequencing or co-culture experiment respectively) and then the insert was placed into the neuronal culture. The dosage of pro-inflammatory factors - LPS and IFNγ (Gibco) was experimentally determined at 10 ng/ml for both compounds (11).
Quantitative polymerase chain reaction
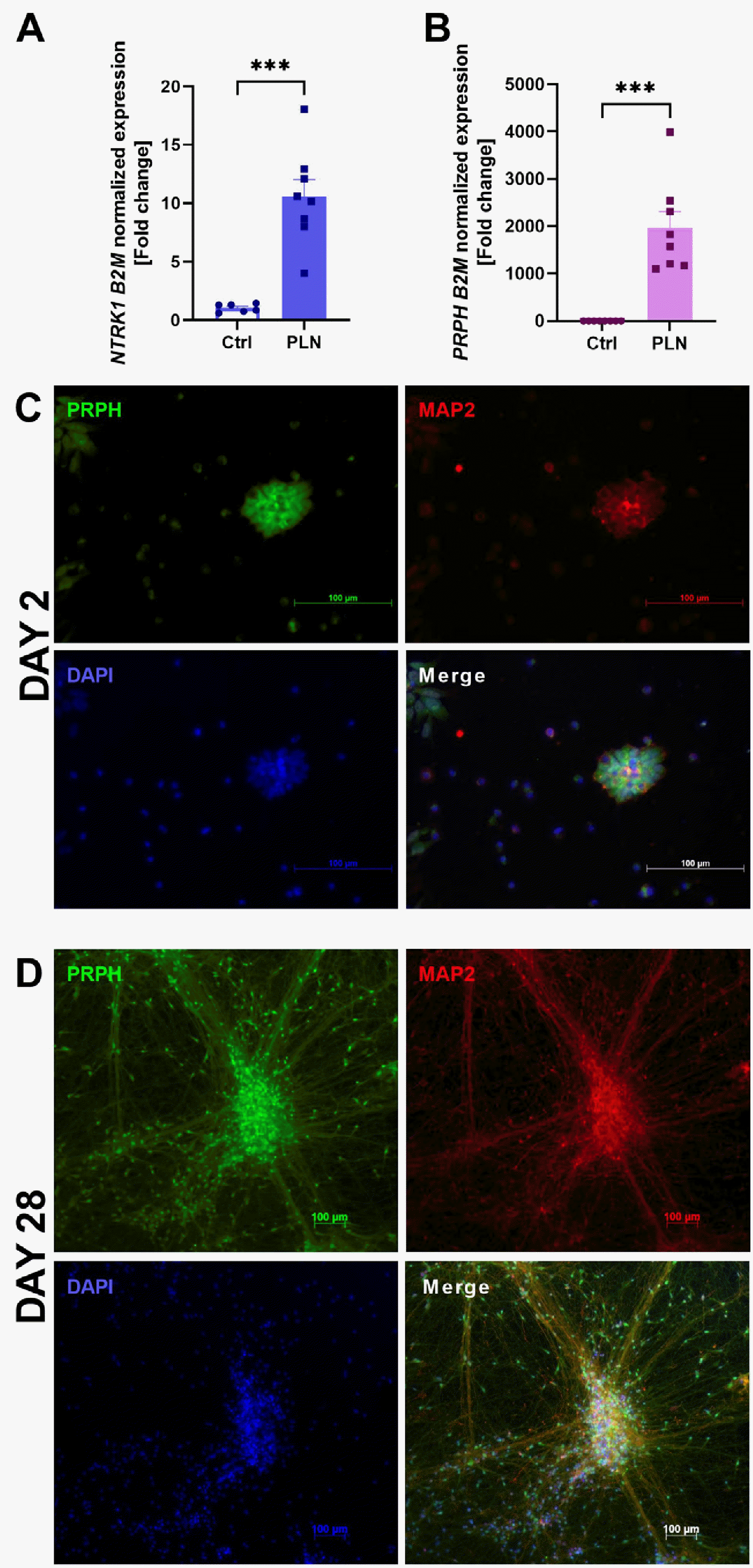
Differentiated PLN were lysed with TRIzol reagent and RNA was isolated according to the supplier’s protocol. The total RNA concentration was measured using an ND-1000 Spectrophotometer (NanoDrop Technologies Inc.) and 500 ng of RNA was used for reverse transcription reaction (EURx). Quantitative polymerase chain reaction was performed in a thermocycler CFX96TM Real-Time system (Bio-Rad) and gene expression was measured with specific Taq-Man probes (PRPH [Hs00196608_m1], B2M [Hs00187842_m1]) and TaqMan Universal PCR Supermix (Bio-Rad). Data were calculated using the ddCt method with normalization to B2M (beta-2-microglobulin) gene expression. The results are presented as fold change of expression level in undifferentiated NESCs.
Immunocytochemistry
For immunocytochemistry, cells were cultured for 2 or 28 days on coverslips coated with laminin and poly-L-ornithine. After differentiation, cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature and then washed with PBS. For antigen retrieval, cells were heated at 95℃ for 10 minutes in buffer (100 mM Tris, 5% urea, pH 9.5). The cell membrane was then permeabilized with PBS containing 0.1% Triton-X-100 and samples were blocked in 5% normal donkey serum (NDS) (017-000-121; Jackson ImmunoResearch) in PBS and incubated overnight at 4℃ with appropriate primary antibodies (anti-peripherin [ab269861; Abcam], anti-MAP2 antibody [ab254264, Abcam], both diluted 1:100) diluted in 1% NDS solution in the SignalBoostTM Immunoreaction Enhancer Kit (407207; Merck Millipore). After three washes in PBS, double immunofluorescence was detected by incubation for 2 hours with the appropriate fluorochrome-conjugated secondary antibodies: donkey anti-mouse Alexa Fluor 488 (A-21202; Invitrogen) and donkey anti-rabbit Alexa Fluor 594 (A-21207; Invitrogen) diluted 1:500 in 5% NDS. Coverslips were sealed with Fluoromount-GTM mounting medium containing DAPI (Thermo Fisher Scientific). Samples were evaluated using an inverted fluore-scence microscope (Axio Observer; ZEISS).
Enzyme-linked immunosorbent assay
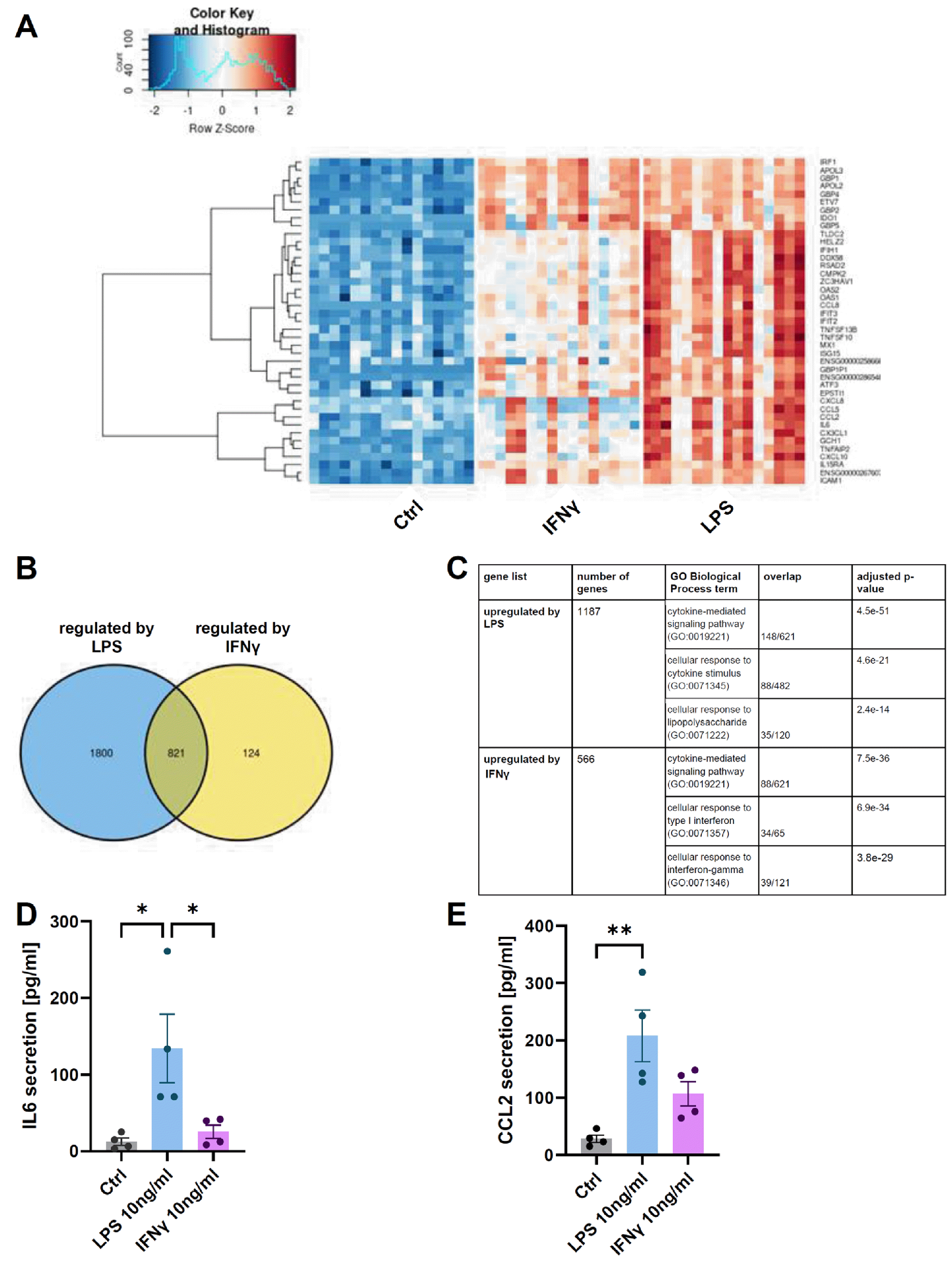
Media for enzyme-linked immunosorbent assay (ELISA) were collected from HFLS cultured on inserts, 24 hours after stimulation with pro-inflammatory factors (10 ng/ml of LPS or IFNγ). Secreted IL6 or CCL2 proteins were measured with an ELISA kit (88-7066 and 88-7399 respe-ctively, Thermo Fisher Scientific) according to the manufacturer’s manual.
The luminescent adenozyno-5’-trifosforan assay
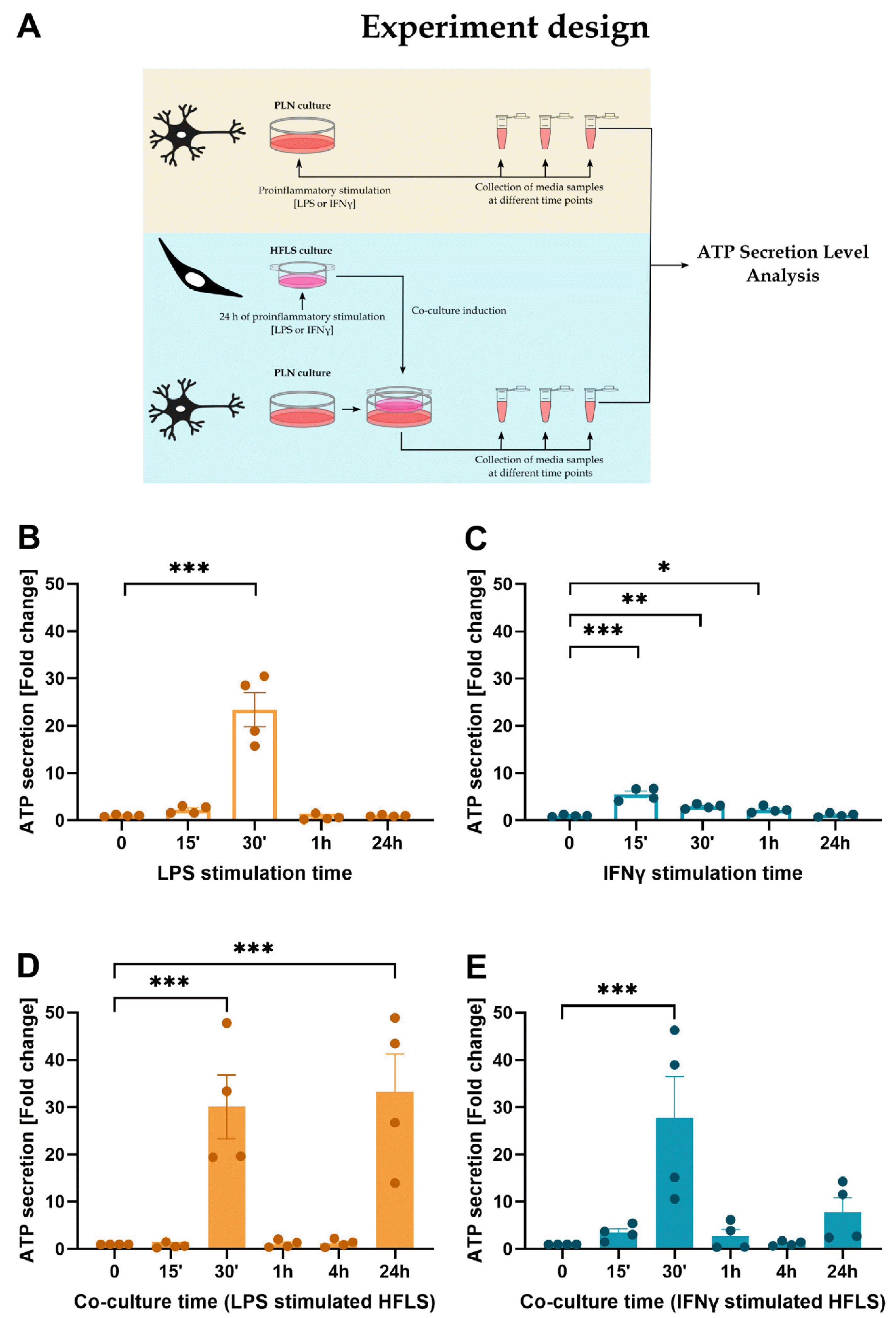
Neuronal activation was measured by adenozyno-5’-trifosforan (ATP) secretion into culture media using an ATP Determination Kit (A22066; Thermo Fisher Scientific). The experiment was performed in two regimes. First, the effect of pro-inflammatory stimulation with LPS or IFNγ on the activity of PLN was tested. Secondly, an insert with control or pro-inflammatory activated synoviocytes (24 hours incubation with the LPS or IFNγ) was added to the neuronal culture and neuronal activity was measured under these conditions. Media were collected at time points after stimulation with pro-inflammatory factors - LPS or IFNγ (10 ng/ml) (15’, 30’, 1 hour, 24 hours) or after addition of inserts to the neuronal culture (15’, 30’, 1 hour, 4 hours, 24 hours). ATP levels were measured according to the manufacturer’s protocol. Briefly, ATP standards (0.01∼1 μM) and measured samples were mixed with a reaction solution (containing D-luciferin and firefly luciferase). Bio-luminescence was measured immediately using GloMaxⓇ Navigator (Promega) and data were calculated as fold change of ATP level in co-culture media from unstimulated synoviocyte cultures or media from unstimulated neurons.
RNA-seq analysis
RNA isolation, library preparation and sequencing have been described previously (8). For this study, cells from OA patients (the control group and cells stimulated with LPS and IFNγ) were selected from the full dataset. All statistical analyses of RNA-seq data were performed with R software 4.2. Statistical significance was tested using two-way RM ANOVA (between-subject factor, treatment; within-subject factor, patient) on log2(1+FPKM) values with a false discovery rate adjustment. False discovery rate (FDR) threshold of 0.01 was used as the significance cutoff. T-tests were used as post-hoc tests and the p-value<0.01 threshold of significance was applied to the results. Enrichment analysis was conducted with the Enrichr tool (12). Within the tool, the Gene Ontology (GO) Biological Process database was used.
Statistics
Statistical analyses were performed using Statistica 13.3 software (StatSoft) using ANOVA or t-student tests depending on the experiment. All biochemical experiments were carried out under the same conditions for all samples, regardless of the type of treatment. All data were obtained in independent experiments and are presented as the mean±SEM.
Results
Neuroepithelial stem differentiation into PLN
After 28 days of cell differentiation, increased expression of peripheral neuron marker genes such as NTRK1 (Fig. 1A) and PRPH (Fig. 1B), which encode peripherin and TrkA proteins, respectively. Furthermore, immunocytochemical staining showed that differentiated cells expressed peripherin and MAP2 (another neuronal marker) proteins (Fig. 1C, 1D) (13), and a significant change in cell morphology was observed.
Changes in the HFLS after pro-inflammatory stimulation with LPS or IFNγ
The second type of cell used in the experiment were HFLS. They form the synovial membrane surrounding the joint and play a key role in the production of hyaluronic acid, the principal component of synovial fluid (14). Our studies have shown that synoviocytes are much more responsive to LPS stimulation than to IFNγ. Transcriptome analysis revealed that 3,059 genes were regulated by either LPS or IFNγ stimulation. The expression of example top genes were visualized in the form of a heatmap (Fig. 2A). The majority of genes were regulated by LPS (Fig. 2B) (2,621 genes with post-hoc t-test p-value<0.01) including 1,187 upregulated and 1,434 downregulated. IFNγ regulated 945 genes out of which 566 were upregulated and 379 downregulated. To uncover common processes in which the genes affected by stimulation are involved, two lists of upregulated genes (by LPS and by IFNγ) were used for enrichment analysis with the GO Biological Process database (Fig. 2C). Our results confirm the data on the high secretory potential of HFLS (15), and the pro-inflammatory agents we used led to a robust immune response. Using ELISA, we have shown that synoviocytes cultured on inserts can be stimulated by the semipermeable membrane and also release pro-inflammatory factors into the medium. A 24-hour stimulation of HFLS with LPS and IFNγ resulted in a 10- and 2-fold increase of IL6 (Fig. 2D) and 8- and 4-fold increase of CCL2 (Fig. 2E) secretion into the culture medium. This means that the use of inserts allows the interaction between the two cell types to be studied without physical contact.
The impact of the pro-inflammatory stimulated HFLS on PLN
In response to stimuli, activated neurons release the neurotransmitter ATP (16). The secretion of ATP can activate purinergic receptors on nearby neurons and glial cells, leading to a variety of cellular responses, including synaptic transmission, neural plasticity and neuroinflammation. The stimulation of the neurons with pro-inflammatory agents (Fig. 3A) showed a significant increase in the level of ATP in the medium after 30 minutes of LPS (Fig. 3B) and 15∼60 minutes of IFNγ (Fig. 3C) stimulation, compared to the control group. Similar to the synoviocyte studies, LPS induced a more potent neuronal activation than IFNγ, but the effect of the second pro-inflammatory agent was observed for longer. Introducing pro-inflammatory stimulated synoviocytes into neuronal cultures induced an approximately 25-fold increase in ATP secretion into the medium after 30 minutes of co-culture (Fig. 3D, 3E), compared to the control group using unstimulated HFLS. ATP levels then returned to control levels, and after 24 hours an increase in secretion was observed, which was significantly higher only in co-culture with LPS-stimulated synoviocytes compared to control group (Fig. 3D). In the second test group after 24 hours of co-culture, activation levels were increased (Fig. 3E), but the result was not statistically significant.
Discussion
The experiments used NESCs differentiated from induced pluripotent stem cells (iPSCs) derived by the Falk group, according to the protocol which has been published previously (17). The next step was to use the neural progenitors differentiation protocol described by Alshawaf et al. (9) and after 28 days of differentiation, an increase in the expression of sensory neuron markers (peripherin and TrkA) and neuronal marker (MAP2) was observed, as in the work from which the protocol was used. Peripherin is a type III intermediate filament protein that is predominantly expressed in peripheral neurons where it forms part of the cytoskeleton (18). It is a useful marker for peripheral sensory and autonomic neurons, as well as their axons and dendrites. TrkA is a protein marker for neurons, particularly sensory neurons. It is a receptor tyrosine kinase activated by the neurotrophin nerve growth factor (NGF) and it is expressed in neurons that respond to NGF, including sensory neurons that transmit informa-tion about pain, temperature, and touch (19). The data obtained allows us to conclude that the phenotype of the differentiated cells is similar to peripheral/sensory neurons.
Studies have shown that HFLS may be involved in the development of chronic joint disorders such as RA or OA (20), and the pro-inflammatory response of these cells may increase the tissue degeneration and pain observed in these diseases (21). As previously shown (11), stimulation by pro-inflammatory factors induces chemokine and interleukin secretion by HFLS. In our experiment, we used two pro-inflammatory agents, LPS and IFNγ; the former induces a strong pro-inflammatory response through TLR4 (toll-like receptor 4) activation and is often used to model infla-mmation in vitro (22), and the second factor acts through IFNGR (interferon gamma receptor) through which it induces a cellular response. Moreover, IFNγ is also a naturally occurring cytokine in the human body (23). The use of two stimulants allowed us to obtain more convincing data regarding the response of cells to inflammation and compare the pro-inflammatory effects of a substance widely used for modeling inflammation (LPS) with a more physiological cytokine (IFNγ). Our studies have shown that both pro-inflammatory agents led to a robust immune response and that synoviocytes are much more responsive to LPS stimulation than to IFNγ. Moreover, using semipermeable membrane inserts we have proven, that HLFS can be stimulated and release pro-inflammatory factors into the medium to stimulate other cell type in the co-culture. We used this approach to measure ATP secretion by PLN cells.
ATP secretion by neurons is a highly regulated process involving multiple signaling pathways and cellular mechanisms (16). ATP is stored in neurons in synaptic vesicles and released through exocytosis in response to various stimuli, such as depolarization or Ca2+ influx. ATP secretion by activated neurons is important in many physiological and pathological processes, including sensory processing, pain perception, immune responses and neurode-generative diseases (24). Measurement of released ATP was selected because it is fast and easy to determine in a small amount of medium. We showed, that LPS-stimulated HFLS co-cultured with PLN caused an increase of ATP production after 24 hours, which was not observed in the LPS-stimulated neurons. Additionally, the secretion of ATP in the co-culture with IFNγ-stimulated HFLS was increased in comparison to IFNγ-stimulated PLN monoculture, however ATP secretion after 24 hours of co-culture did not reach statistical significance. The early increase in ATP release is probably due to the introduc-tion of a large number of chemokines/cytokines released by the stimulated synoviocytes and present on the surface of the insert. The second wave of increased activity is pro-bably caused by factors newly synthesized by the synoviocytes and released into the medium.
The in vitro models used to study joint diseases are mainly based on the culture of cells of a single type, in mono-layers. This approach is being refined by using three-dimensional-printed scaffolds (25) or matrigel (26), which allow to obtain a complex cellular structure resembling tissue. The model we use is a simple, fast and inexpensive method to study the biochemical interaction of any two cell types via the secretome, without physical interaction between them. This approach is particularly advantageous for co-culturing rapidly proliferating cells (such as synoviocytes) with cells that require long culture for differentiation (such as neurons). However, our research has some limitations, mainly due to the lack of markers for sensory neurons. Two different markers (TrkA and peripherin) were selected based on the Alshawaf publication (9) and a general neuronal marker (MAP2), therefore we use the general term PLN in this paper. Expanding the panel of markers and defining the cells studied as sensory or nociceptive neurons would be more beneficial from the perspective of studying the pain response observed in the development of joint diseases such as OA and RA. Although the derived system appears to be a suitable tool for studying the factors that promote the development of the disease, it may also find application in the search for potential therapeutic substances and learning about their mechanism of action.
Summary
The model presented here appears to be an interesting tool for investigating both the mechanisms of cell-cell interactions in tissues and the therapeutic potential of drugs such as analgesics. The ability to use different types of cells introduced into culture makes it possible to model various disorders and interactions that occur in different tissues, so it seems reasonable to further develop this approach for the evolution of in vitro studies.
 XML Download
XML Download