

 PDF
PDF Citation
Citation Print
Print



Introduction
Induced pluripotent stem cell (iPSC) technology has sparked a revolutionary transformation across the fields of stem cell research, developmental biology, disease modeling, and personalized medicine. IPSCs can be generated through the reprogramming of somatic cells, often achieved by overexpressing four transcription factors (i.e. Oct4, Sox2, Klf4, c-Myc) that orchestrate a pluripotency auto-regulatory circuit (1, 2). While bypassing ethical concerns associated with embryonic stem cells, iPSCs share attributes similar to embryonic stem cells, exhibiting an indefinite self-renewal capacity and remarkable potential to differentiate into the three germ layers. This facilitates studies on human development, patient-specific disease modeling, and drug testing without ethical concerns. Additionally, iPSCs represent a promising cell source for regenerative medicine as they can be used to improve the function of damaged tissues without eliciting an immune response.
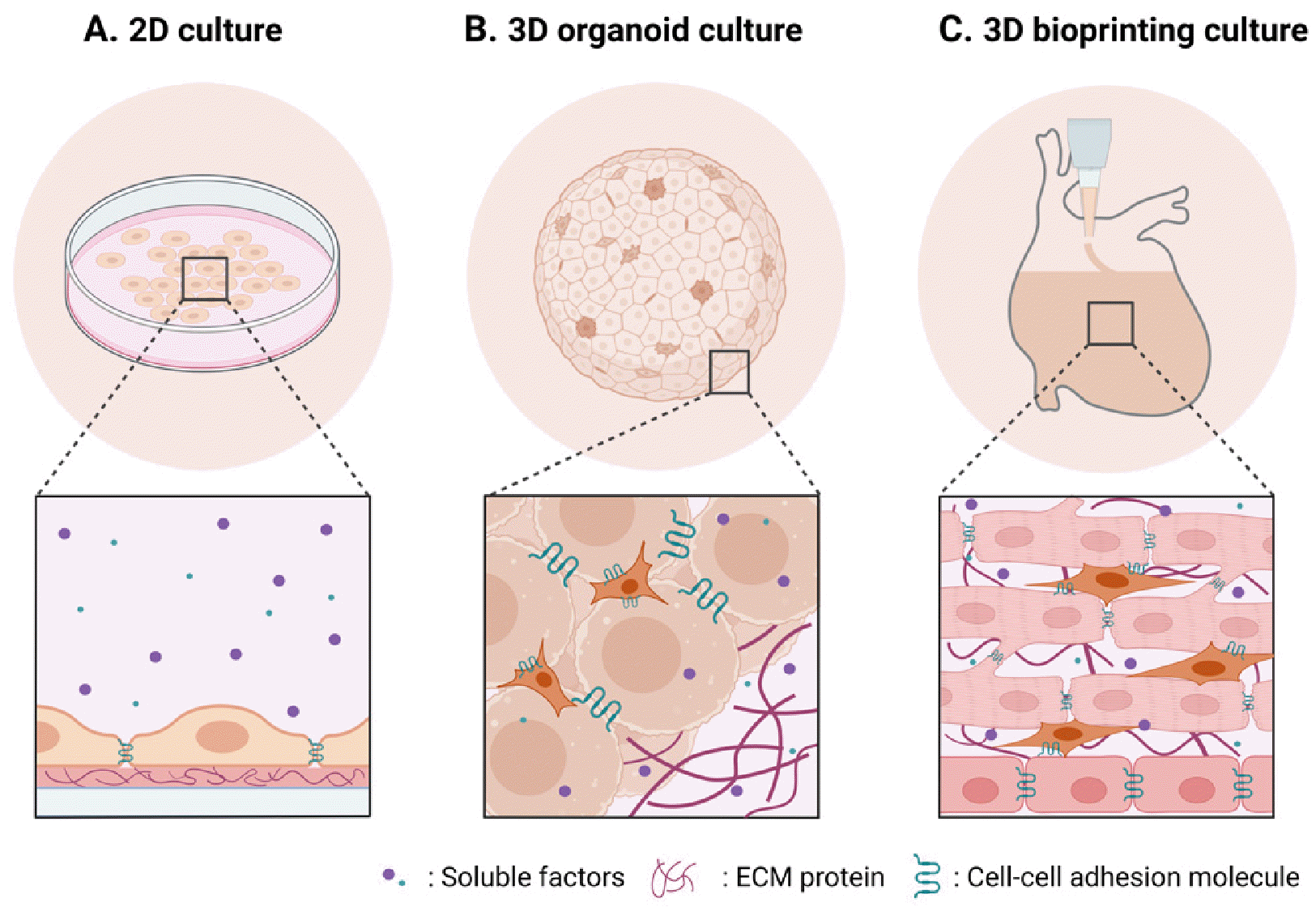
To fully harness the capability of iPSCs, development of in vitro culture models that can support the maintenance of pluripotency of iPSCs is of great importance. Over the decades, iPSC culture condition has evolved from two-dimensional (2D) models using poorly defined, animal-derived products to the human-derived, fully defined models, to more physiologically relevant three-dimensional (3D) models (Fig. 1, Table 1) (3-7). In the beginning, iPSCs were maintained in animal-derived serum containing media on either animal-derived feeder cell layer or animal-derived extracellular matrix (ECM) coating materials such as MatrigelTM (1, 8-10). Then, these animal derived products were replaced by human-derived products, which was still poorly defined (11-13). Further studies identified fully defined conditions, composed of soluble factors such as basic fibroblast growth factor (bFGF) and transforming growth factor-β (TGF-β), in combination with ECM proteins such as collagen IV, fibronectin, laminin, and vitro-nectin, for maintaining pluripotency and genomic stability of iPSCs (14-21). Advantages of utilizing fully defined 2D culture condition include (1) high reproducibility, (2) ease of high throughput analysis, and (3) relatively low variabi-lity (Fig. 1A). However, 2D culture models do not fully recapitulate 3D microenvironment, which hampers studying more physiologically relevant cell-microenvironment interactions. In addition, there is a limitation in scaling-up.
Recent studies demonstrated that 3D iPSC culture methods of forming spherical aggregates of iPSCs or spheroids for suspension culture can successfully support the maintenance of iPSC pluripotency and long-term propagation (22, 23). These iPSC spheroids can be further directed to differentiate into multiple lineages such as brain (24, 25) and heart (26, 27), and can self-organize into organ-mimicking 3D structures, termed as organoids (Fig. 1B). These 3D iPSC culture systems reflect cell-cell and cell-ECM interactions more effectively. In addition, the system enables more scalable culture comparable to their 2D adherent counterparts. However, the current 3D iPSC culture systems often have simple geometry such as sphere, limiting construction of complex tissue structure. When grown beyond diffusion limit (∼200 to 300 μm), 3D iPSC spheroids/organoids develop a necrotic core, thereby restricting their use in long-term studies. 3D iPSC spheroids/organoids are often generated manually and thus can be a labor-intensive process with high batch variability, which limits the scalability and reproducibility.
3D bioprinting is an advanced biotechnology that involves layer-by-layer deposition of biological materials, such as cells, growth factors, and biomaterials, to construct complex 3D tissue structures. The application of 3D bioprinting to iPSCs or iPSC-derived cells can address the limitations of current 2D and 3D iPSC culture systems. Compared to monolayer 2D cultures or simple spherical 3D culture systems, 3D bioprinted iPSCs can be fabricated into complex tissue structures that mimic the architecture and functionality of native tissue at both macro- and micro-levels (Fig. 1C). Moreover, the 3D microenvironment of bioprinted iPSCs can be finely tuned for their specific applications using bioinks. Additionally, the precise and rapid control offered by 3D bioprinting enables high scalability and reproducibility.
In this review, we will discuss the latest advances in iPSC bioprinting, with a specific focus on the research progress on undifferentiated iPSC bioprinting. First, we review the concepts and techniques of 3D bioprinting, followed by the basic principles of design strategy of bioink. We then review and summarize the research advancement of 3D bioprinting especially using undifferentiated iPSCs. Finally, the current limitations and future prospects of bioprinting undifferentiated iPSCs will be discussed.
3D Bioprinting
3D bioprinting is a cutting-edge approach to fabricating biological tissues and organs by precisely depositing cell-laden materials layer by layer, following a computer-designed blueprint. Unlike traditional 3D printing that uses various materials like plastics or metals as an ink, 3D bioprinting uses the “bio-ink” composed of living cells, biomaterials, and other biologically active compounds such as growth factors. 3D bioprinting process involves several key steps: (1) creation of digital 3D model of the desired tissue or organ based on medical images or user-defined models; (2) engineering of bioink mimicking the cellular microenvironment in the tissue of the interest; (3) biopri-nting; and (4) post-printing processes such as crosslinking and maturation.
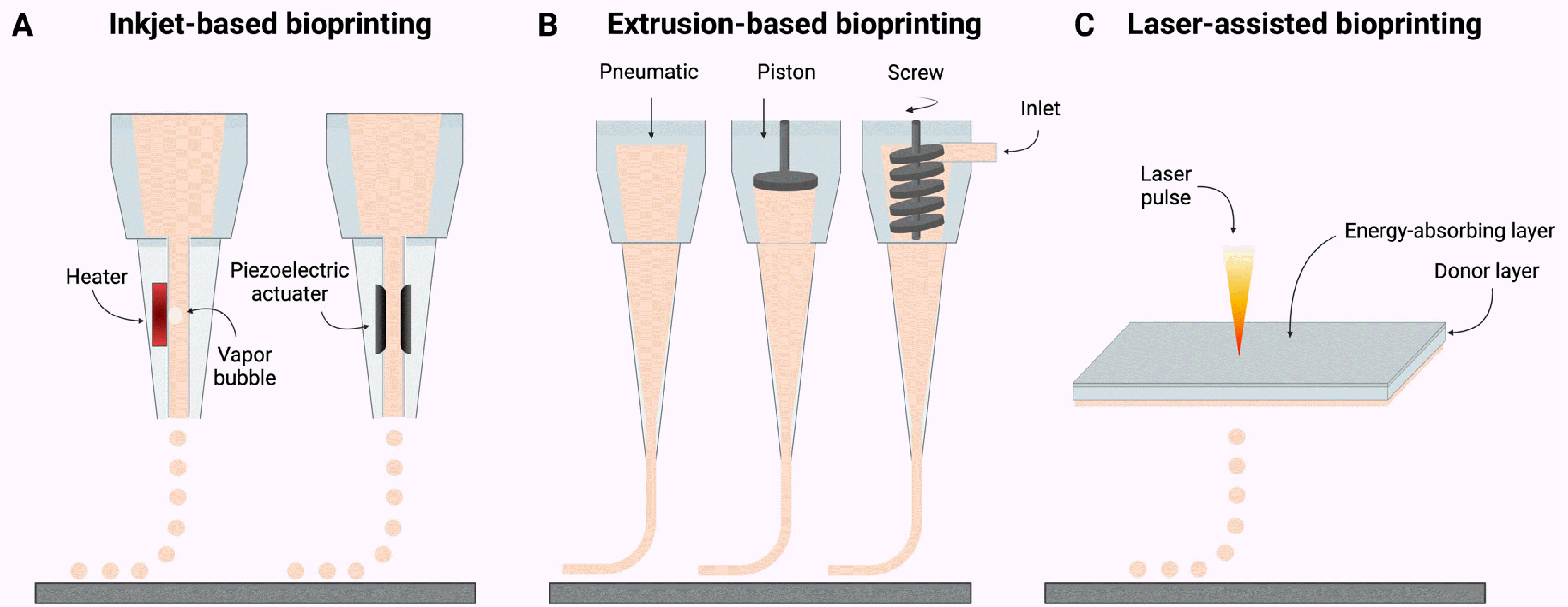
There are three representative 3D bioprinting techniques, namely inkjet-based, extrusion-based, and laser-assisted bioprinting (Fig. 2, Table 2) (28-40). These three 3D bioprinting techniques have been utilized for studies of 3D iPSC bioprinting.
Inkjet-based bioprinting
Similar to the function of a regular inkjet printer, inkjet-based bioprinting employs a print head to eject picoliter-sized droplets of bioink onto a substrate in a specific pattern at a rapid printing speed (up to 10,000 droplets per second) (41). Inkjet bioprinting is recognized for its high resolution (50∼300 μm) and speed (28), making it suitable for creating intricate structures. However, it has limitations in handling viscous or dense bioinks, thereby restricting the use of materials within a narrow viscosity range (3.5∼12 mPa/s) and relatively low cell density (<106 cells/ml) (29, 30, 41, 42).
Extrusion-based bioprinting
In extrusion-based bioprinting, a bioink is extruded through a nozzle or syringe via pneumatic or mechanical pressure. Extrusion bioprinting presents several advantages over other printing techniques. It can handle a wide range of bioink viscosities (30∼6×107 mPa/s) (29) and high cell density (>108 cells/ml) (28), making it versatile for printing various tissue types. In addition, it has been demonstrated for producing human-scale large constructs (43) or large batches of tissues (44), demonstrating its high scalability. Furthermore, extrusion bioprinter is relatively cost-effective and ease of use, making it accessible to researchers (31). There are several limitations associated with extrusion bioprinting. Given the cells in bioink are subject to mechanical stress during extrusion process, cell viability and functionality may be compromised. Bioink should be carefully designed to minimize the mechanical stresses. Also, extrusion printing may have limitations in achieving high-resolution features due to the nozzle size and viscosity of bioinks. Nozzle clogging with viscous ink can also be an issue (32).
Laser-assisted bioprinting
Laser-assisted bioprinting is a sophisticated technique that employs lasers to precisely deposit cells and biomate-rials in a controlled and non-contact manner. This technique utilizes the composite slide composed of an energy-absorbing layer, a donor layer, and a bioink layer containing cells and biomaterials. A pulsed laser is used to generate a focused energy pulse that creates a rapid localized pressure increase in a donor layer, leading to expelling micro-sized bioink droplets onto a substrate to form a desired pattern (45). Laser-assisted bioprinting offers several advantages including high precision (>0.5∼20 μm) (29, 46), non-contact printing, minimal damages from thermal and mechanical stimuli, which together reduces the risk of cell damage and nozzle clogging and enables printing of intricate tissue structures (28, 29). However, it comes with a set of challenges such as the complexity and high cost associated with the laser system and the potential need for specific laser-absorbing materials for different bioinks (29).
Each of these bioprinting techniques has its own advantages and limitations. The choice of a bioprinting technique should consider factors such as the desired tissue type and structure, and the required resolution. Depending on the selected bioprinting approach, the optimization of bioink is necessary for maximizing the potential of bioprinting and creating intricate and functional tissues.
Bioink
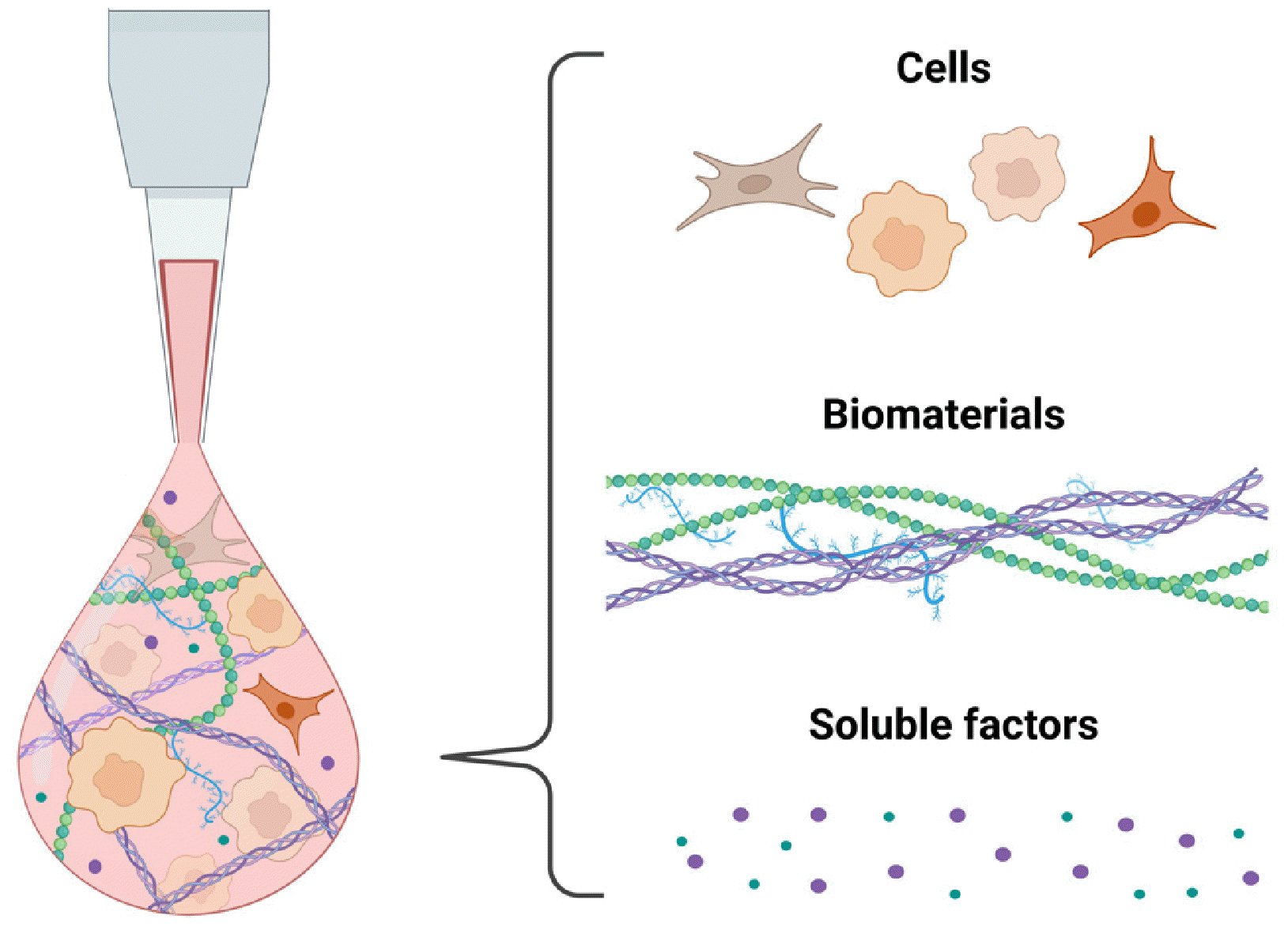
As a key technology in 3D bioprinting, bioink is responsible for carrying and supporting encapsulated cells throughout the printing process with the ultimate goal of creating functional tissues or organs. Based on the principles of tissue engineering, bioinks are carefully formulated to contain (1) the cell types found in the target tissue; (2) biomaterials that support the viability of encapsulated cells during printing process and provide instructive, biomechanical and biochemical cues for tissue formation; and (3) bioactive factors that can guide the behaviors of encapsulated cells for functional tissue formation (Fig. 3).
Selection of cells
Selecting the right cell types in bioink is critical for achie-ving the desired functions of the resulting bioprinted tissues. Tissues are composed of diverse types of cells, each with unique and specialized functions. For example, in cardiac muscle tissues, cardiomyocytes are the main players responsible for contraction-relaxation function of the tissue. Other cell types, such as cardiac fibroblasts, also play an important role in maintaining the structural integrity and overall function of the cardiac muscle. One strategy is to formulate a single bioink using either a single cell type that is responsible for major tissue function (47) or a mixture of multiple cell types (48). Alternative strategy is to formulate multiple separate bioinks, each containing one cell type to achieve cell-specific distribution in the tissue (49, 50). Progenitor cells or stem cells can be used as a cell source in bioink, where they can differentiate into the cell types needed for creating functional tissues, with appropriate differentiation signals provided in the bioink. Multipotent stem cells such as mesenchymal stem cells or adipose-derived stem cells, as well as pluripotent stem cell-derived cells, have been widely used for 3D bioprinting (51-53). Utilizing stem cells in bioink offers several advantages over using fully differentiated cells for biopri-nting. These advantages include the ability to recapitulate complex cellular diversity due to the differentiation potential of stem cells, along with achieving a more physiologically relevant cellular arrangement due to their capacity for self-organization.
Selection of biomaterials
Hydrogels are most widely used biomaterials for 3D bioink. Hydrogels are a 3D network of hydrophilic polymers that can retain water similar to body water content (∼80% to 90%) with high structural integrity and bio-compatibility. Hydrogels can provide and/or be easily engineered to provide biomimetic cues present in the native cellular microenvironment so that cells can grow, differentiate, and develop into functional tissues in vitro. Hydrogels can be categorized into two types based on their origin: (1) natural polymer-based hydrogels and (2) synthetic polymer-based hydrogels.
Natural polymer-based hydrogels are derived from naturally occurring polymers found in living organisms. Widely used natural polymers in bioink include collagen (54-56), alginate (57-59), and basement membrane proteins such as MatrigelTM (60-62). These hydrogels offer advantages such as biocompatibility, bioactivity, and biodegradability. How-ever, their mechanical properties are often suboptimal and the tunability is limited.
Synthetic polymer-based hydrogels are created by chemically synthesized polymers and thus offer more precise control over hydrogel properties such as mechanical strength and degradation rate. Commonly used synthetic polymer-based hydrogels include polyethylene glycol (63-65) and poly(caprolactone) (66, 67). While these synthetic polymer-based hydrogels can be tailored to match the required characteristics of specific tissues and applications, they often lack bioactivity. To address the limitations, resear-chers have engineered the composite hydrogels by combining natural and synthetic polymers. Examples include gelatin-alginate (68-70), carboxymethyl chitosan (70, 71), and hydroxypropyl chitin (72). Researchers continue to explore novel design strategies to create hydrogels that more closely mimic the native ECM and promote tissue formation.
Selection of bioactive factors
The third component of bioink is bioactive soluble factors. Bioactive soluble factors include growth factors, cytokines, and other signaling molecules that regulate cell behaviors such as proliferation and differentiation. The goal of incorporating these factors in bioink is to create a microenvironment within the printed construct that supports cell viability, proliferation, and differentiation for tissue formation and maturation. Therefore, bioactive soluble factors need to be carefully selected based on the microenvironment of the target tissue.
Bioink design criteria
To ensure the success of the bioprinting process and the viability and functionality of the printed constructs, several design criteria of bioinks should be considered. First, bioinks must be biocompatible to ensure the viability and functionality of encapsulated cells. The components of bioink and their degraded parts should not induce toxicity or inflammation. Bioinks should also have appropriate rheological properties to ensure smooth flow of the ink during printing followed by maintenance of their shape after deposition. Printability of bioinks is influenced by the viscosity, shear-thinning property, and other mechanical properties of the bioink. Furthermore, biomimicry of bioinks is an important design criterion. Cell behaviors are well known to be guided by biomechanical and biochemical properties of the target tissue (73-76). Therefore, it is crucial to match the biomechanical properties of bioink to those of the target tissue, and to mimic the biochemical properties of bioink by incorporating relevant ECM proteins and soluble factors. Lastly, biodegradability of bioink is essential to allow encapsulated cells to form intercellular connection and remodel their surrounding environment with newly produced ECM. In sum, bioink design is a critical aspect of bioprinting to produce viable and functional bioprinted tissues for various applications.
IPSC-Based Bioprinting
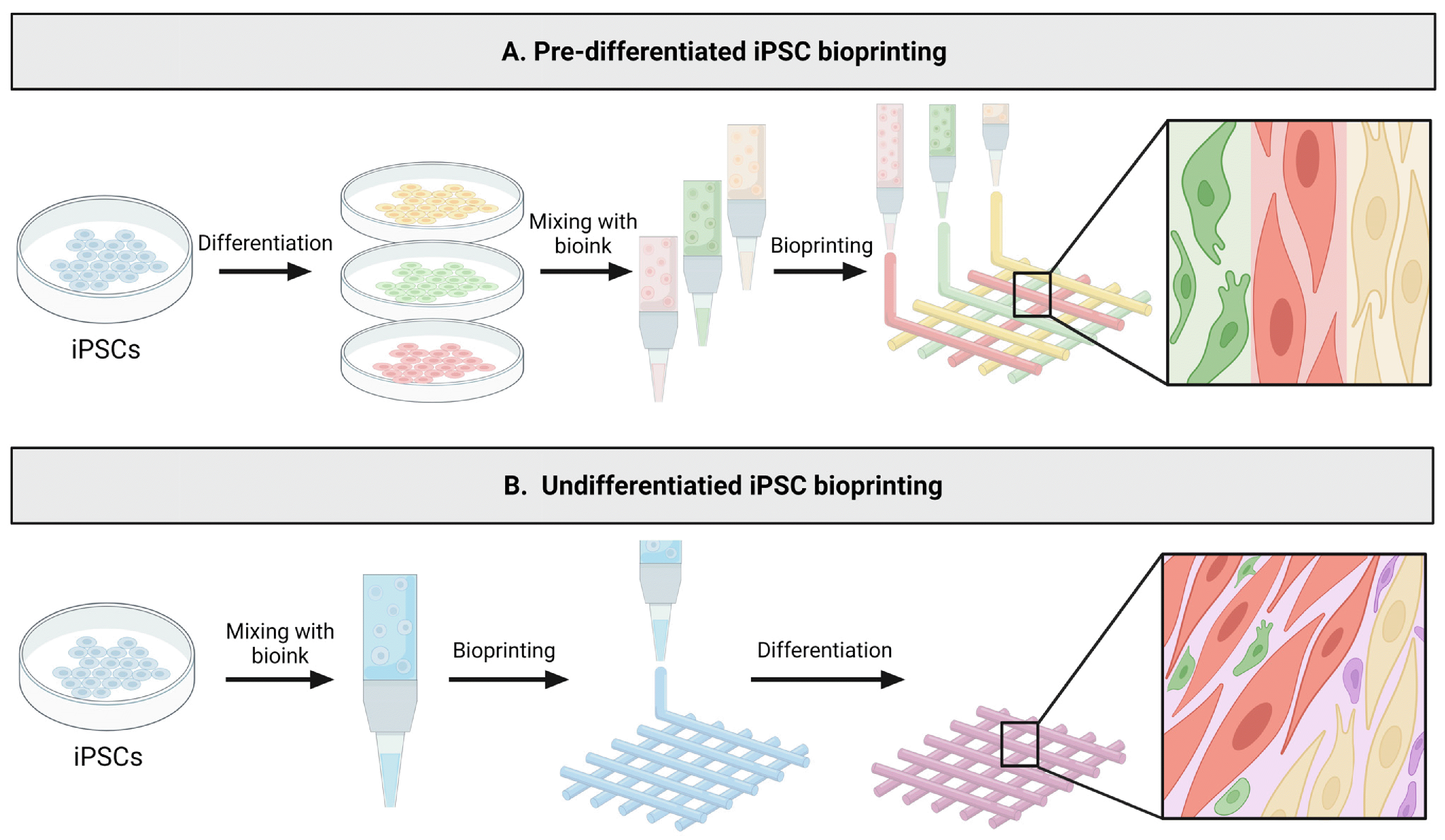
Current iPSC-based bioprinting studies utilize two main strategies: (1) pre-differentiated iPSC bioprinting and (2) undifferentiated iPSC bioprinting. In the case of pre-differentiated iPSC bioprinting, iPSCs are initially differen-tiated into specific cell types in a conventional 2D culture condition. Subsequently, these cells are encapsulated in cell-specific bioink for 3D bioprinting (Fig. 4A). On the other hand, the approach of undifferentiated iPSC-based bioprinting, also known as post-differentiation iPSC-based bioprinting, involves the direct encapsulation and 3D bioprinting of undifferentiated iPSCs. Following printing, these iPSCs are directed to differentiate into specific lineage within the bioprinted tissue construct (Fig. 4B). The selection and application of the iPSC-based bioprinting strategies should consider the respective advantages and disadvantages associated with each method.
Pre-differentiated iPSC bioprinting
Previous studies have extensively utilized pre-differentiated iPSC bioprinting to fabricate a range of tissues, including cartilage, heart, nerve, liver, and skin. This topic has been thoroughly reviewed in numerous review articles, and thus it is not within the scope of this review (please refer to (77).
Utilizing pre-differentiated iPSCs provides several advan-tages. First, it provides better control over bioink formulation due to a more controlled differentiation of iPSCs in a 2D setting. Additionally, the pre-differentiated iPSCs can be encapsulated in biomaterials customized to guide specific cellular behaviors in a cell-specific manner. By utilizing multiple cell-type-specific bioinks, it is possible to recapitulate the diversity and distribution of cells present in tissue through spatial patterning of various cell types. For example, Noor et al. (50) used two cell-specific bioinks, namely a cardiomyocyte-specific, and endothelial cell-specific bioinks to create structurally complex and functional cardiac tissues. However, pre-differentiated iPSC bioprinting has its disadvantages. The resolution of current bioprinting technology limits the recreation of micro-structures present in tissues. Also, cell-cell integration is restricted, particularly between cells in different bioinks (49, 78-80).
Undifferentiated iPSC bioprinting
Compared to pre-differentiated iPSC bioprinting, the bioprinting of undifferentiated iPSCs has received relatively less attention. One of the advantages of undifferentiated iPSC bioprinting is the versatility of the printed tissue constructs as they can subsequently undergo differentiation into various tissues. In addition, since iPSCs, during differentiation, give rise to diverse cell types found in tissues and these cells can self-organize, post-differentiation of the bioprinted iPSC constructs closely emulate the process of organoid formation, which can lead to better recapitulating the native tissue structure and function. Moreover, thanks to their remarkable proliferative capacity, the formation of stable and well-connected tissue structures can be more readily achievable (81). In the following section, we will summarize the research progress on the bioink laden with undifferentiated iPSCs.
Bioink development for undifferentiated iPSC bioprinting
Previous studies have reported the development of undifferentiated iPSC-laden bioink that supports high viability post-printing and maintains pluripotency (Table 3) (49, 59, 72, 81-85). Earlier research adopted two distinct strategies for creating bioinks containing undifferentiated iPSCs: one without scaffolding materials and the other incorporating scaffolding materials. The use of undifferentiated iPSC-laden bioink without scaffolding materials facilitates cellular proliferation and the secretion of ECMs, resulting in the formation of tissue structures resembling the process of organoid development. Due to its simplicity, this strategy was employed to generate a high-throughput array of spheroids or organoids (82). Nonetheless, bioprinting iPSCs without scaffolding materials poses limitations on fabricating intricate 3D structures. Hence, the incorporation of scaffolding materials in bioink is imperative for achieving more complex 3D structure.
In order to bioprint complex 3D structures using undifferentiated iPSCs, researchers have investigated the utilization of diverse combinations of biomaterials and bioactive factors to formulate undifferentiated iPSC-laden bioinks. One of the commonly used biomaterials is alginate (49, 83, 84). Alginate is a polysaccharide extracted from brown algae, that has been widely used as a tissue engineering scaffold due to its biocompatibility, non-immunogenicity, low toxicity, and hydrophilicity (86). Alginate, a negatively charged polymer, can rapidly be crosslinked under the presence of calcium ions and its viscosity can be easily controlled, which makes it an attractive material for bioink. Since alginate lacks bioactivity, it is essential to add ECM proteins and bioactive factors to support the viability and maintenance of pluripotency in encapsulated iPSCs. Particularly, a mixture of basement membrane proteins derived from mouse sarcoma tissue, such as MatrigelTM or GeltrexTM, has been used to enhance the bioactivity (72, 82, 84, 85). It was reported that increasing MatrigelTM concentration leads to an increase in survival and aggregation of the iPSCs (72). Employing these bioinks, previous studies have demonstrated that the bioprinted iPSCs can retain their pluripotency, along with the capability to undergo spontaneous differentiation into the three germ layers and even be guided towards cardiomyocyte differentiation. (82, 85).
Since MatrigelTM or GeltrexTM originates from animal sarcoma tissues, and their components are poorly defined, efforts have been made to develop novel bioinks with more defined bioactive components. For instance, when alginate-based bioinks were supplemented with chitosan or fibrinogen, the encapsulated undifferentiated iPSCs showed pluripotency marker expression (OCT4, SOX2, NANOG, SSEA4, TRA-1-60), proliferation, and differentiation into three germ layers (49, 83). Nonetheless, these proteins are not naturally present during embryo development. Additional research is necessary to determine whether recapitulating the microenvironment of epiblasts can further enhance the self-renewal and differentiation capacity of the bioprinted iPSCs.
Precisely guiding the fate of iPSCs into tissues of interest is another important aspect to consider when designing an iPSC-laden bioink. It is well-known that iPSC differentiation can be efficiently guided by mimicking the native cellular microenvironment of the desired tissue (87). To design biomimetic bioinks, the microenvironment of the targeted tissue can be recapitulated by including combinations of ECM components and growth factors abundantly found in the tissue, or using decellularized ECM derived from the tissue (Table 4) (88-101). There are only a few studies on an undifferentiated iPSC-laden bioink that is inductive to differentiation by mimicking the microenvi-ronment of the target tissue. For example, Kupfer et al. (81) demonstrated the differentiation of bioprinted iPSCs into cardiomyocytes and the formation of perfusable two-chambered hearts by developing a novel bioink composed of defined components––collagen methacrylate, laminin-111, and fibronectin––that are known to promote cardiac cell differentiation. Similarly, Nguyen et al. (84) employed a bioink incorporating hyaluronic acid, a key component of the chondrocyte microenvironment, to promote the differentiation of chondrogenic lineages after undifferentiated iPSC bioprinting. In summary, tissue-mimetic bioinks designed for bioprinting undifferentiated iPSCs have been demonstrated to be effective to support viability, preserve pluripotency, and enable both spontaneous and directed differentiations.
Challenges and Future Prospects of Undifferentiated iPSC Bioprinting
Previous studies on undifferentiated iPSC bioprinting have demonstrated high viability, pluripotency retention, and differentiation potential post-printing. Despite these advancements, there are still several limitations that need to be addressed for the broader application of undifferentiated iPSC bioprinting.
First, most previous studies demonstrated bioprinting undifferentiated iPSCs in simple tissue structures such as droplet (82, 85) or grid structures (49, 72, 83, 84). Future studies should explore printing more sophisticated structures that are physiologically relevant. Another bottleneck of undifferentiated iPSC bioprinting technology is poor in
tercellular integration post-printing. Although iPSCs present remarkable proliferative capacity, current bioinks reported seem insufficient to support proliferation, migration, and tissue formation. This can be overcome by several strategies such as using cell aggregates rather than single cells (102, 103) and incorporating inducive ECM proteins and/or bioactive molecules that can promote proliferation and migration. Furthermore, previous studies have only validated three germ layer specification and directed differentiation towards only one or two selected cell types such as neuron, chondrocyte, or cardiomyocytes (81, 83-85). Since iPSCs possess broad differentiation potentials, it would be interesting to explore the versatility of the undifferentiated iPSC-laden bioink. For example, it is worth investigating whether bioprinted undifferentiated iPSCs using one type of bioink can be further differentiated into diverse cell types post-printing. Finally, for producing 3D complex and thick tissues, the creation of intricate vascular network structures is imperative to prevent cellular necrosis within the inner regions of the bioprinted tissues.
Undifferentiated iPSC bioprinting holds promising future prospects for advancing various research fields, including tissue engineering, developmental biology, disease modeling, and drug screening. Specifically, undifferentiated iPSC bioprinting has the potential to revolutionize personalized medicine by enabling the construction of patient-specific tissues that mimic the intricate geometry and structures of native tissues and organs. Integrating undifferentiated iPSC bioprinting with microfluidic systems could lead to the development of novel “organ-on-a-chip” platforms, offering a more precise representation of human physiological system for developmental studies and disease modeling. The capability to bioprint arrays of undifferentiated iPSCs could enhance scalability and reduce variability, and thereby facilitate high-throughput drug screening. This innovative technology offers an alternative to using model organism-based preclinical models, holding the potential to facilitate the drug development process. As such, these prospects highlight the exciting potential of undifferentiated iPSC bioprinting, which converges cutting-edge iPSC technology and 3D bioprinting techniques, accelerating both basic research and the translation of scientific discoveries into therapeutic applications.
 XML Download
XML Download