

 PDF
PDF Citation
Citation Print
Print



Introduction
The lung is a complex organ that consists of a tree-like arrangement of branched airways connecting the large airway (trachea) and millions of terminal gas-exchange units (alveoli) (1). Along this complex architecture, the airways and alveoli are surrounded by different mesenchymal cells, which support the maintainence of lung homeostasis (2). However, the use of classical cell line model systems in pulmonary biomedical research has certain shortcomings. It is cultured on plastic and has different cellular heterogeniety and physiological differences found in vivo. Animal models can be used for this reason; however, extrapolating results from animals to humans is risky considering the different genetic backgrounds. Furthermore, the cost is high and moral issues must be addressed in animal experiments (Fig. 1).

Lung organoids and lung-on-a-chip are reasonable solutions to these issues. Organoids are three-dimensional (3D) self-organized construct made up of multiple cell types that reflect in vivo cellular composition. They grow within gels of an extracellular matrix (ECM) protein mixture with a cocktail of growth factors, which can provide equivalent environmental cues found in vivo (3). They can be generated from either adult stem cells (ASCs) or pluripotent stem cells (PSCs) and share a structural similarity to in vivo (4). Additionally, the organoid system is fast, easy, and robust research system that can be rapidly established, easily manipulated, and personalized (5).
In contrast, lung-on-a-chip is a biomimetic microsystem that resembles the functional alveolar-capillary interface of the human lung (6). It comprises an air channel lined with epithelial cells, a blood channel coated with endothelial cells, and a membrane to mediate two compartments. On-a-chip models support customized modification to apply microfluidic systems to resemble the blood flow for blood channels or vacuum to mimic human breathing. Therefore, lung organoids and the on-a-chip system is getting more attention from researchers (Fig. 2).
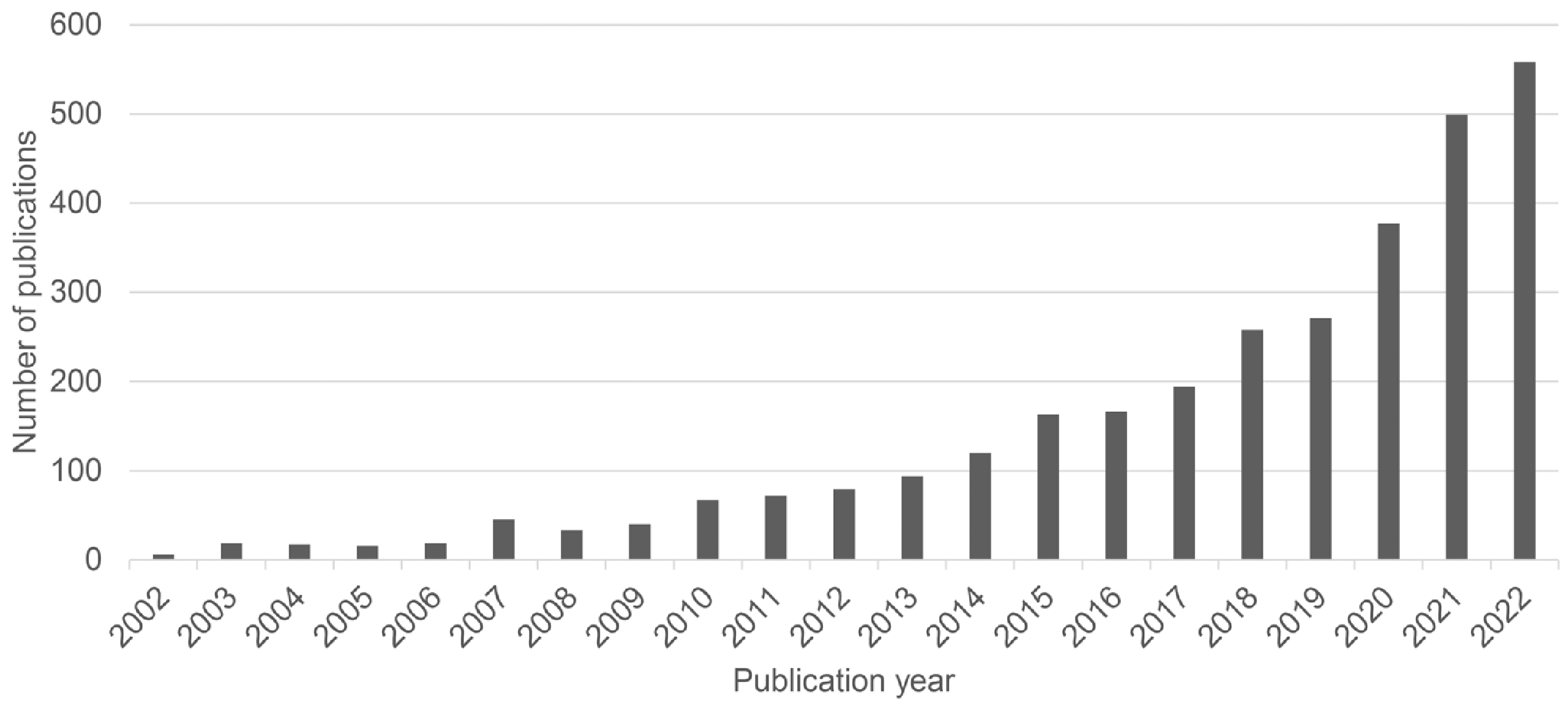
This review summarizes the core components and previous biomedical studies using lung organoids and lung-on-a-chip models and introduces possible future applications. We would like to highlight the importance of these models, hoping that this will lead biomedical researchers to consider using these platforms to understand pulmonary diseases, such as emerging viral infections, progressive fibrotic pulmonary diseases, or primary or metastatic lung cancer.
Go to : 
Technological Considerations
Cell sources
The standard cell sources for human lung organoids include ASCs and PSCs (Table 1). ASCs are undifferentiated cells that are surrounded by differentiated cells that regulate stem cell homeostasis in a tissue or organ and are characterized by quiescence and plasticity (7). ASCs can be acquired directly from patients but can only differentiate into cells found in the lung (8). When cells are implanted in 3D, lung ASCs can differentiate, generating lung epithelial cells like goblet and ciliated cells that cover the luminal side of the lung spheroid (9). ASCs can be embedded into ECMs to mimic in vivo conditions.
Table 1
Two major types of stem cells used to develop organoids
![]()
On the other hand, PSCs are cells that can self-renew and develop into diverse germ cell layers and eventually all cell types of the human body (10). To form structures such as native fetal lungs, PSCs should be differentiated into endodermal progenitors and anterior foregut endoderm over 9∼10 days. Structures similar to those of adult airways can only appear after in vivo transplantation (11). PSC-derived lung organoids are also embedded in the ECM and niche factors, where they can be directed to form lung organoids (5).
ECM
ECMs are necessary for the formation of lung orga-noids. ECMs include basement membrane extracts, synthetic scaffolds, and a decellularized ECM. Among basement membrane extracts, the most widely used material is MatrigelTM, which is extracted from Engelbreth-Holm Swarm mouse sarcomas (12). MatrigelTM consists of four major proteins (laminin, collagen IV, entactin, and heparin sulfate proteoglycan perlecan), tumor-derived proteins (transforming growth factor and fibroblast growth factor), and enzymes such as matrix metalloproteinase (13). This composition of MatrigelTM provides biological functions in vitro such as cell differentiation, tumor growth, and angiogenesis (12). Despite the fact that MatrigelTM is commonly used, it has issues of variable composition, high batch-to-batch variability, and potential antigenicity (13). Considering these limitations, synthetic scaffolds have been evaluated as reasonable alternatives to Matrigel. One of the most commonly studied and used scaffolds is polyethylene glycol (13). This material is advantageous for cell culture since it is hydrophilic, bioinert, and highly amenable to chemical modifications. Finally, by decellularization, ECMs can be directly derived from humans or animals (14). Decellu-larization removes allogenic or xenogeneic cells from the tissue while leaving behind the ECM scaffold. Such an ECM preserves the protein composition and ultrastructure of native ECM. However, decellularized ECMs can have variable chemical properties, depending on the ECM source and processing method. This can lead to low reliability and reproducibility of results.
Organ-on-a-chip
The 3D cell culture techniques are known to have many advantages over two-dimensional (2D) cell culture systems. They promote responses to stimuli, the expression of genes and proteins, and enhance cell function (15). However, the lung is an essential organ that facilitates constant breathing, with microvessels flowing beneath the surface. Organ-on-a-chip models provide a more lifelike imitation of the in vivo environment than the static trans-well condition, which lacks relevant mechanical properties. Lung-on-a-chip devices can generate mechanical stretching of epithelial cells by negative pressure in the top channel and dynamic fluid flow in the bottom channel. A microfluidic device constructed on a co-culture system of epithelial and endothelial cells demonstrated inflammation-induced thrombosis when exposed to external stimuli, such as lipopoly-saccharide (16). Jain et al. (16) showed that lipopolysa-ccharide endotoxin indirectly stimulates intravascular thrombosis and that lung-on-a-chip models can be a novel methodology for in vitro analysis of potential drug candidates.
The unique function of the lungs is gas exchange between oxygen and carbon dioxide. This can be monitored within the chip with a sensor system that measures hypoxia in real time along with the lung disease process (17). Alveolar diseases are characterized by the formation of solids or liquids in the alveolus, which is usually filled with air. The “increased shade” findings are observed via chest imaging (18). Suppose that this physiological phenomenon can be maintained under a microscope and the degree of shading can be quantified. More complex in vitro lung modeling is expected to be possible in this case.
Air-liquid interface
When inhaled or aerosolized particles enter the respiratory tract, they first encounter the surfactant film, which is a thin lining layer along the surface of the lung (19). Beneath the surfactant film, the airway is lined with alveolar type I and II cells in close connection with endothelial cells to facilitate gas exchange. This is called the air-liquid interface (ALI), which is a unique structure of the lungs. Considering such physiology, culturing airway and alveolar cells in traditional submerged cell-culture conditions cannot truly mimic pulmonary physiology in vivo (20). Models including 3D cultures under ALI conditions mimic better in vivo environment of the lung which provide more physiological condition (21). ALI is instrumental in conditions involving aerosolized exposure, such as in air pollutant models or inhaled therapeutics. Several types of ALI have been developed utilizing not only BEAS-2B, A549, and NCH-H441 or primary human cells (22), but also lung-on-a-chip technology (23).
A list of the key references for the technological considerations mentioned above can be found in Table 2.
Table 2
The main references for the technological issues with organoids and organ-on-a-chip models
| Categories | Explanation | Reference |
|---|---|---|
| Cell sources | Includes adult stem cells and pluripotent stem cells |
Kim et al., 2020 (5)
Clevers, 2015 (7)
Tian et al., 2021 (8)
Rock et al., 2009 (9)
Hofer and Lutolf, 2021 (10)
Dye et al., 2016 (11)
|
| Extracellular matrix | Sources required for the formation of organoids such as basement extracts, synthesis scaffolds, and a decellularized extracellular matrix |
Orkin et al., 1977 (12)
Aisenbrey et al., 2020 (13)
Lee et al., 2021 (14)
|
| Organ-on-a-chip | Can mimic the breathing motion with mechanical stretch, and blood flow with fluidic system at bottom channels |
Gupta et al., 2016 (15)
Jain et al., 2018 (16)
Ding et al., 2021 (17)
|
| Air-liquid interface | Provides physiological exposure of epithelial cells to air |
Lenz et al., 2013 (20)
Wu et al., 2017 (21)
|
![]()
Go to :
Clinical Applications
Respiratory infection
The coronavirus disease 2019 pandemic accelerated the establishment of respiratory infection models using organoids or on-a-chip platforms. Lamers et al. (24) developed a 2D differentiated small airway model to reproduce severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection in vitro. Because primary cell models have donor-to-donor variation, restricted usability, and a lack of cellular heterogeneity, they used self-renewing organoids that can differentiate into tissue-specific cell types. Lung organoids from two donors were cultured in an ALI set-ting. The organoids differentiated not only into alveolar cells but also into basal cells and rare neuroendocrine cells. When SARS-CoV-2 infected organoids, viral replica-tion was mainly targeted at type II alveolar (ATII) cells, for which the model was mainly composed. A low dose of interferon-λ1 treatment significantly reduced viral replication (24).
Si et al. (25) repurposed an existing antiviral agent against influenza A and SARS-CoV-2 using the human lung bronchial cell-on-a-chip technology. Airway chips were manufactured using a commercialized organ-on-a-chip (Chip-S1TM) from Emulate Inc., made of polydimethylsiloxane, including two parallel channels and porous membranes coated with collagen IV from the human placenta. Under ALI conditions, human lung bronchial epithelial basal stem cells were put into the apical channel. In contrast, primary human microvascular endothelial cells were seeded in the vascular channel and exposed to flowing media. The main characteristic of this chip was that epithelial basal stem cells were able to differentiate into several types of cells, such as ciliated cells and mucus-producing goblet cells, which are highly similar to human airways. The influenza A virus was introduced into the model and an immune response was induced, such as increased barrier permeability and the production of inflammatory cytokines. The inhibitory effects of potential antiviral treatments on pseudo-typed SARS-CoV-2 were tested after introducing an anti-influenza metabolite via the chips (25).
Pulmonary fibrosis
Few studies have been done on pulmonary fibrosis using organoids or on-a-chip technologies because fibrotic changes in the lungs need to be seen and studied over a long period of time. Suezawa et al. (26) developed a model of idiopathic pulmonary fibrosis (IPF) model using fibroblast-dependent alveolar organoids (FD-AOs) consisting of human PSCs and primary human fetal lung fibroblasts. Researchers used organoids to establish the model because the primary cells have high variability (24). Researchers have emphasized the importance of interactions between epithelial cells and fibroblasts in IPF models. IPF is initiated by damage to alveolar epithelial cells, particularly ATII cells, and progresses through fibroblast activation. FD-AOs were cultured by encapsulation in MatrigelTM. When FD-AOs were treated with bleomycin, shrinkage of the matrix and contraction of the lung tissue occurred, which is one of the phenotypes of IPF. Additionally, bleomycin treatment induced morphological changes in epithelial cells, increased type II alveolar-type I alveolar (ATII-ATI) intermediate-state cells, and cellular senescence in ATII cells, indicating the pathogenesis of IPF. However, this model has limitations in that bleomycin treatment causes damage to both epithelial cells and fibroblasts. Although the pathogenesis of IPF observed in this model was not stronger than that observed in mouse models, FD-AOs may be helpful in drug screening for IPF (26).
Plebani et al. (27) created a human cystic fibrosis (CF) airway chip model that can overcome the shortcomings of murine models. With the Chip-S1TM from Emulate Inc., epithelial bronchial cells from a patient were planted on top of the porous membrane to establish the ALI. In contrast, primary human lung microvascular endothelial cells were co-cultured below the membrane and exposed to a dynamic fluid flow. In the CF airway chip, thick mucus accumulated with a high ciliary density and beat frequency. Secretion of interleukin-8 and recruitment of polymorphonuclear leukocytes have been observed in CF airway chips, which are related to non-resolving neutrophilic inflammation. Furthermore, Pseudomonas aeruginosa, one of the most common bacterial pathogens, thrives because of its thick mucus layer. Furthermore, cytokines such as interleukin-6, tumor necrosis factor-α, and granulocyte-macrophage colony-stimulating factor were highly expressed in the chips. The limitations of this study were that the endothelial cells in the chips were not patient-derived, and some types of cells in the lung tissue, such as lung fibroblasts and pulmonary macrophages, were missing. However, human CF airway chips are a promising alternative to murine models, having high reproducibility in mimicking the structural and pathological features of human CF airways (27).
Lung cancer
Lung cancer exemplifies a disease for which patient-derived organoids (PDOs) can be used. Long-term in vitro expansion of lung cancer organoids replicates the tissue architecture and genomic alterations of primary lung tumors. Lung cancer organoids have responded to drugs based on genomic alterations, including EGFR mutations to erlotinib and MET amplification to crizotinib (28). Lung cancer organoids closely replicate the genomics and biology of patient tumors and may serve as a testing platform for drugs and biomarkers (29). In such perspective, Hu et al. (30) established a method to shorten the time required for drug sensitivity tests based on PDOs. First, the researchers processed tumor tissues from patients into small pieces to produce enough PDOs within three days and then cultured them with MatrigelTM with growth factors over-night. The PDOs were morphologically and genotypically similar to the original tumor tissues. Next, the PDOs were seeded and cultured on an integrated superhydrophobic microwell array chip (InSMAR-chip) for a three-day drug screening. The drug testing results with PDOs on the InSMAR-chip and the clinical outcomes with the original tumor tissues were concordant (30).
Park et al. (31) developed a 3D vascularized lung cancer-on-a-chip (VLCC) that mimics the tumor microenviro-nment and metastasis. This model was used to examine the function of doxorubicin. The chip was based on decellularized lung ECM, including ECM constituents such as collagen and proangiogenic factors such as growth factors and cytokines. The ECM was mixed with collagen to construct a tissue-mimetic hybrid hydrogel. It incorporates lung cancer-derived tumor spheroids consisting of malignant epithelial cells (A549), human lung fibroblasts, and human umbilical vein endothelial cells. The VLCC also includes two cylindrical macrochannels corresponding to the structure of a large vessel. Along with angiogenic sprouting towards the spheroids, capillary networks were successfully formed between channels due to pro-angiogenic factors in the decellularized ECM. Moreover, the VLCC showed significant differences in cell death depending on the doxoru-bicin concentration, which is known to induce apoptosis.
Table 3 provides examples of disease modeling using lung organoids and the lung-on-a-chip models mentioned above.
Table 3
Examples of lung organoids and lung-on-a-chip models used in disease modeling
| Disease | Modality | Details | Reference |
|---|---|---|---|
| Respiratory infection | Organoid | Organoid derived bronchioalveolar model for SARS-CoV-2 infection | Lamers et al., 2021 (24) |
| Lung-on-a-chip | Influenza A virus infection model using epithelial basal stem cells cultured on lung-on-a-chip (Emulate Inc./Chip-S1TM) | Si et al., 2021 (25) | |
| Pulmonary fibrosis | Organoid | Idiopathic pulmonary fibrosis model using fibroblast-dependent alveolar organoids | Suezawa et al., 2021 (26) |
| Lung-on-a-chip | Cystic fibrosis airway chip modeling using patient-derived epithelial bronchial cells cultured on lung-on-a-chip (Emulate Inc./Chip-S1TM) | Plebani et al., 2022 (27) | |
| Lung cancer | Organoid | Drug screening using patient-derived tumor organoids | Kim et al., 2019 (28) |
| Lung-on-a-chip | Three-dimensional vascularized lung cancer-on-a-chip model mimicking tumor microenvironment and metastasis | Park et al., 2021 (31) |
![]()
Go to :
Ensemble of Lung Organoids and Organs-On-A-Chip
Organ-on-a-chip technology relies on our knowledge of human organs to engineer manufactured chip constructs in which cells and their microenvironments are precisely con-trolled. Organoids, on the other hand, grow grom self-organizing stem cells and follow their own development programs to make structures and functions that are very similar to their in vivo counterparts. The combination of organ-on-a-chip technology and organoids can more realistically mimic the human body characteristics. Researchers are now exploring the possibility of combining the best features of each approach to develop a more potent in vitro technology.
Given the spheroid shape of a typical lung organoid, it is challenging to create a structure that is physiologically similar to the lung, such as an ALI (32). But it is important to set up a co-culture system to mimic how the human lungs function within the alveolus-interstitium-vessel boundary (33). This can be achieved thanks to on-a-chip technology; organoids-on-a-chip has several strengths compared to the simple organoid of organ-on-a-chip technology. First, vascular systems can be introduced. With cancer cell-derived organoids, the spheroid structure surrounded by microvessels had a similar shape and cellular density as in vivo models (34). The vascular system can be mimicked by creating an endothelial barrier-trapping ECM (35). Second, the interstitial space can be imitated. The interstitial space of the lung is mainly composed of the ECM and fibroblasts, which should be used to create a meticulous lung model (36). When a disease occurs in the lung, various immune cells are mobilized from outside the lung and cause different immune responses at the alveolar interstitium-vascular boundary (37). Third, a composite mixture of epithelial cells can be generated from the organoids. In most lung-on-a-chip models, only one type of cell line is cultured on a biometric membrane (38). However, human alveoli are composed of a mixture of 90% ATI cells and 10% ATII cells (39). Constructing such a ratio of epithelial cells is more likely to be achieved through organoid culture than through 2D culture. ATI and ATII cells are present in alveolar organoids created from normal lungs (40). Fourth, the physical factors can be controlled for micromechanical or microfluidic processes. To better mimic the actual organ, it is necessary to resemble respiratory movement, also called breathing. It is well known that lung epithelial cells show a biological response in response to a physical reaction. Most diseases in the alveoli are caused by changes in airflow. SARS-CoV-2 and fine dust are representative examples of stimulants that can cause diseases of the alveoli through breathing movements (Fig. 3).
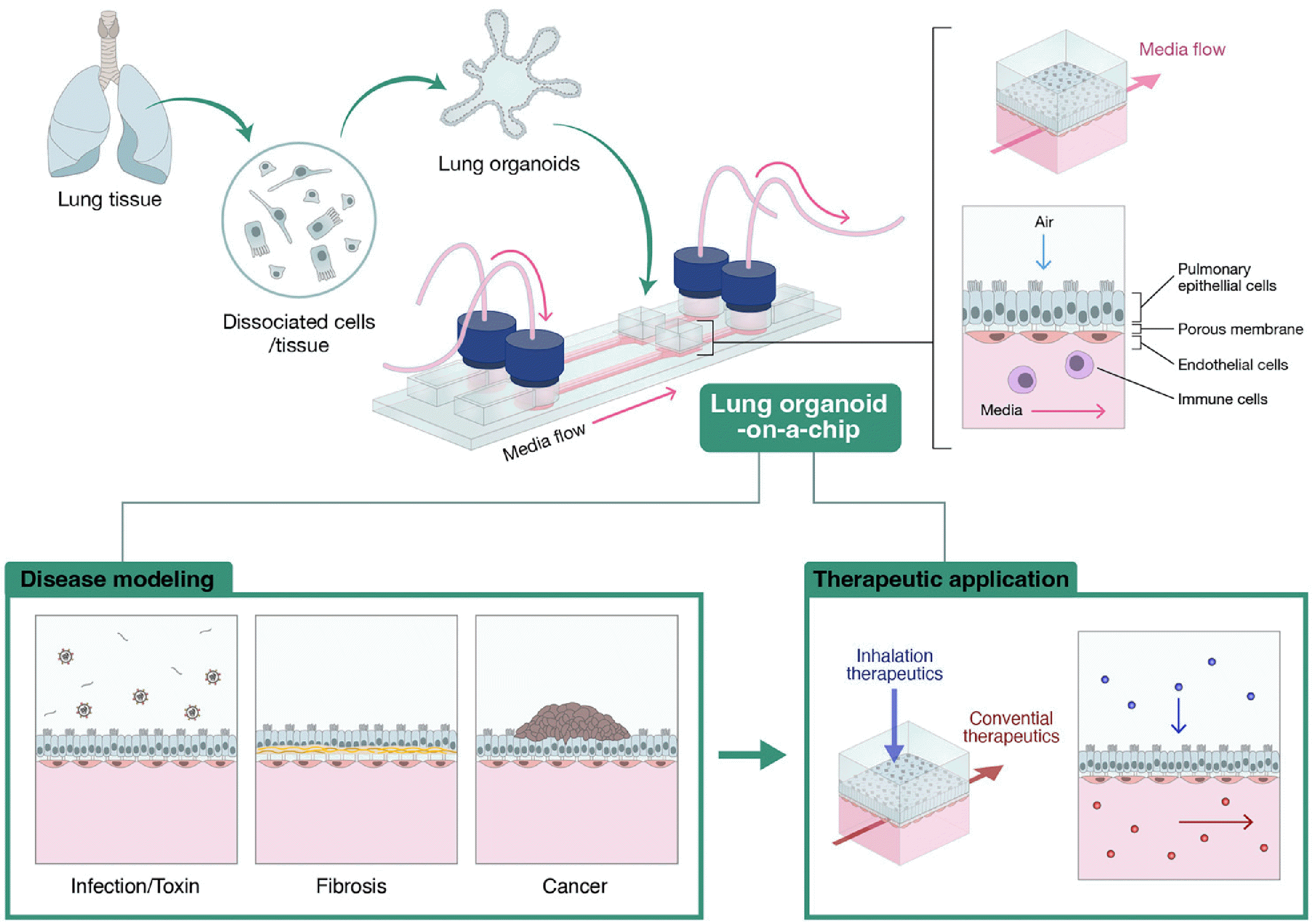 | Fig. 3Ensemble of lung organoids and organs-on-a-chip. Lung organoids derived from the patient can be effectively integrated into the organ-on-a-chip methodology, thereby facilitating disease modeling and screening of the-rapeutic drugs. The distinctive archite-cture of the on-a-chip technique facilitates the flow of media and co-cultiva-tion of diverse cell types.
|
Go to :
Conclusions and Future Perspectives
In this review, we introduced the next-generation in vitro lung organoid and lung-on-a-chip models and their achievements in lung diseases and drug development. Both lung organoids and lung-on-a-chip models have advantages and shortcomings. We highlight the importance of complementing each other through an appropriate combination. Considering the importance of vascular structure for the function of the lung, it is also crucial to develop a co-culture system of lung cells with endothelial cells. Furthermore, there is no complete model for understanding lung diseases that considers the correct inflammatory interactions with other cell types, such as immune cells and fibroblasts, or that considers the biomechanical movements of the lung. Moreover, it is currently impossible to study inter-organ communication; multi-organ-n-a-chip can also be considered to solve such limitation.
Nevertheless, it is undoubtedly true that organoids and lung-on-a-chip models have initiated new opportunities to understand lung physiology, and there is an enormous ongoing effort to make them step forward, which will provide more in-depth insights into lung diseases.
Go to :
 XML Download
XML Download