

 PDF
PDF Citation
Citation Print
Print



Introduction
The endocrinology of type 2 diabetes (T2D) and its predisposing factors have been studied extensively [1] while its skeletal effects have received negligible research [2]. Fractures in T2D are exacerbated by hyperglycemia and disease progression [3, 4]. Proximal humerus fractures (PHF) account for 4% to 10% of all fractures, constituting the 2nd most common fractures of the upper arm [5, 6], and the 3rd commonest fracture site after the proximal femur and distal radius [7]. A study on the humerus of female T2D Goto-Kakizaki rats demonstrated that fracture risk in T2D varies with specific sub regions in bones [8].
The cellular and molecular association between PHF and T2D has not been fully elucidated. However, previous studies suggest that T2D induces osteopenia [5] that compromises bone turnover [9, 10], as well as an imbalance between pro and anti-osteogenic cytokines [11, 12]. Reduced bone turnover is directly linked to altered bone cell numbers and their function [12-14].
Tartrate-resistant acid phosphatase (TRAP) as contained in osteocytes, participates in osteolysis in response to pathologic conditions such as T2D [15, 16]. Additionally, TRAP positive osteocytes, also found in elevated levels in T2D, attract osteoclast precursors to their vicinity, accelerating bone matrix resorption and increasing bone fragility [17, 18]. Osteoblasts secrete osteoid and mineralize the organic matrix under the influence of ALP [19]. Studies on the femur of the Zucker Diabetic Fatty rat found impaired osteoblast differentiation, reduced osteoblast numbers and less matrix mineralization in diabetes [8, 20, 21].
The extracellular bone matrix cytokine transforming growth factor beta 1 (TGFβ1) [22, 23] is responsible for regulating differentiation, proliferation, and migration of osteoprogenitor cells [24, 25]. In contrast, bone morphogenetic protein 3 (BMP3) is antagonistic to the TGFβ1 functions [23]. In addition, T2D facilitates accumulation of extracellular aged glycation end products (AGEs) that adversely crosslink bone matrix collagen, compromising skeletal integrity [26]. This occurs by inhibiting osteoblast differentiation and proliferation and induction of osteoblast apoptosis while increasing osteoclastic and osteocytic bone catabolism [27-30]. Relatively recent studies show that increased skeletal fragility in T2D is exacerbated by bone marrow adiposity due to the differentiation of skeletal stem cells into adipogenic cell lines instead of osteogenic cell lines [31, 32].
We aimed to compare bone cell quantities and immunolabel osteogenic and antiosteogenic cytokines in the Zucker Diabetic Sprague Dawley (ZDSD) and SD rat humerus. In this study, we will start by assessing bone samples for adipocyte accumulation. We will then quantify the bone turnover cells namely, osteocytes and osteoblasts using TRAP and ALP as cell markers respectively. Expression of the pro-osteogenic cytokine TGFβ1 [33], and the anti-osteogenic cytokine BMP3 [23], will be investigated, as they are reported to be altered in T2D [11]. The extent of glycation end-product accumulation will be assessed by quantifying AGEs immunopositive bone cells together with its extracellular expression.
Materials and Methods
Animals
Approval for use of animals was received from the Animal Ethics Screening Committee of the University of the Witwatersrand (clearance number: 2015/07/28C). The study used 23 male rats consisting of 8 SD and 15 ZDSD. The SD rats, which served as controls were sourced from the Central Animal Services (CAS) University of the Witwatersrand. The SD rats were used as the non-diabetic controls because they are the parental strains of the ZDSD rats [26]. All animals were housed one per cage, maintained under pathogen free conditions, in a temperature-controlled environment of 23°C±2°C under a 12-hour light and dark cycle.
Grouping
The ZDSD rats were further divided into moderate diabetes (ZDSD-MOD) (n=9) and severe diabetes (ZDSD-SVD) groups (n=6). The diabetic groups were defined based on oral glucose tolerance test (OGTT) as follows: (1) Moderately diabetic (ZDSD-MOD) animals had >8.0 mmol/L fasting blood glucose (FBG) prior to glucose load and >12.5 mmol/L 2 hours after glucose loading, and more than 50% increase in area under the curve (AUC) after OGTT. (2) Severely diabetic animals (ZDSD-SVD) had >11.2 mmol/L FBG prior to glucose load and >23.0 mmol/L 2 hours after glucose loading, and more than 200% increase in AUC.
Dietary scheme
All animals were fed ad libitum a standard Purina 5008 chow (Lab diet) and given water, ad libitum. At 15 weeks, the ZDSD rats were changed into a high fat diet (5SCA; Test diet), where 47.7% of the kilocalories is derived from fat. These animals were switched back to the standard Purina 5008 chow after 6 weeks. The high fat diet was used to synchronize the onset of hyperglycemia in the ZDSD rats [26].
Blood tests
Fasting blood glucose
Animals were fasted for 12 hours overnight prior to blood glucose measurements. A glucometer (Performa Accutrend; Roche Diagnostics) was used to measure FBG by lancing the tail vein, every 2 weeks.
Oral glucose tolerance tests
Oral glucose tolerance tests (OGTTs) were performed monthly following a 12 hours fast. Fasting glucose levels were recorded at time zero (T0) before receiving an oral glucose load (2 g/kg). Glucose levels were measured from blood collected from the tail vein at 15, 30, 60, and 120 minutes intervals after the glucose load.
Termination and bone harvesting
At the age of 28 weeks, the animals were injected with a lethal intraperitoneal dose of phenobarbital. Humeri were removed, and muscles cleaned out. Individual bones were then stored in 10% buffered formalin for fixation until further processing.
Tissue processing for immunohistochemical techniques
Bone samples were fixed in 10% buffered formalin for a minimum of 14 days, then decalcified in 20% w/v ethylenediaminetetraacetic acid (pH 7.4) for 10 days in an incubator at 45°C with regular gentle shaking. The bones were then rinsed, and their proximal epiphysis severed about 3 mm below the intertubercular groove, then immersed in 10% buffered formalin for 24 hours before processing for histology. An automatic processor (Citadel 2000; Thermo Fischer Scientific) was used to take the samples through ascending grades of ethanol prior to embedding in paraffin wax. During embedding, bone samples were orientated longitudinally to minimize differences in the sectioning angle. Bone sections were cut at 5 μm thickness using a microtome (Leica RM2125 RTS; Leica Biosystems) and mounted on silane-coated glass slides. Three sections per slide were mounted. Section sequences were mounted in order for specific stains, namely; Hematoxylin and Eosin (H&E) to assess humerus head and morphology, and to quantify adipocyte numbers; Immunolocalization of TRAP, ALP, BMP3, TGFβ1, and AGEs for the same region.
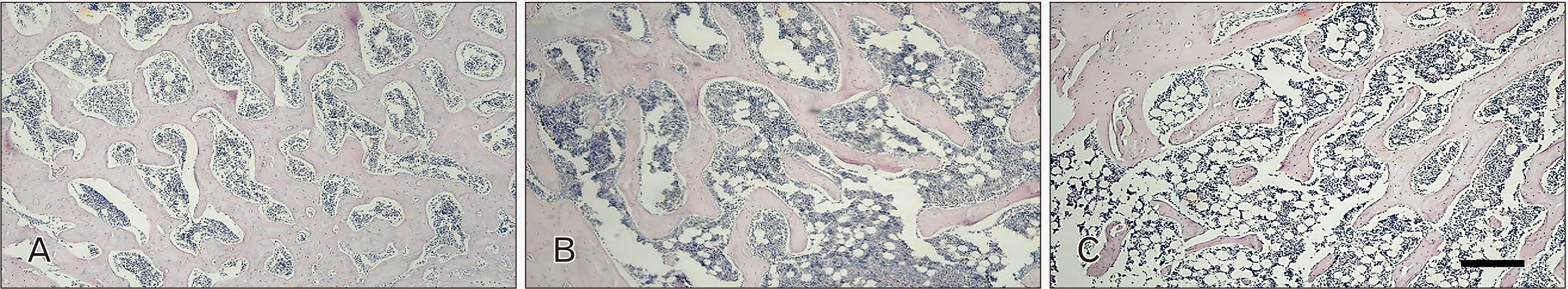
Hematoxylin and Eosin stain
Sections were dewaxed in xylene for 10 minutes and hydrated by passing through 3 changes (1 minute each) of absolute alcohol followed by 90% and 70% alcohol (1 minute each) and eventually into running tap water for 1 minute. The sections were then stained for 10 minutes in haematoxylin, differentiated in periodic acid for 10 seconds, rinsed in tap water for a minute, blued in Scott’s tap water for 2 minutes, and rinsed again in tap water. The cytoplasm was stained with eosin by immersing the sections in 70% alcohol followed by eosin for 2 minutes. The sections were then dehydrated in increasing concentrations of alcohol before mounting in entellen.
Immunohistochemistry for TRAP, ALP, BMP3, TGFβ1, AGE in the head of the humerus
Immunohistochemistry was conducted using various antibodies (Table 1). All antibodies were sourced from Abcam. Sections were incubated at 40°C for 30 minutes, deparaffinized and rehydrated in a series of descending grades of alcohols, then immersed in a citrate buffer (pH=6) and placed in a water bath set at 60°C for 24 hours. This was to facilitate antigen retrieval. After 24 hours sections were cooled at room temperature for 20 minutes then rinsed 3×5 minutes times in phosphatebuffered saline (PBS; pH=7.4). Endogenous peroxidase was blocked with 1% hydrogen peroxide (H2O2) and washed in PBS 3×5 minutes. Sections were transferred to a perspex plate and incubated with primary antibodies at appropriate dilutions (Table 1) and placed at 4°C overnight. After 24 hours, sections were acclimatized for 20 minutes until room temperature, before washing in PBS (pH=7.4), 3×5 minutes. Sections were soaked in biotinylated secondary antibody (Biotinylated Goat Anti-Rabbit IgG (H+L) (ab64256) for 10 minutes, again washed in PBS (pH=7.4) 3×5 minutes. They were incubated further in streptavidin HRP for 10 minutes (ab64269) to reduce background immunopositivity and enhance antibody binding. PBS (pH=7.4) rinses were performed (3×5 minutes). Sections were then incubated with 3-3’ di-aminobenzidine working solution while monitoring color development under a light microscope. Once optimal immunopositivity was achieved, sections were, rinsed in running tap water for 5 minutes, then dehydrated through graded alcohol, cleared in xylene and mounted on silane-coated slides and entellen.
Photography, quantification, and immunopositivity intensity of region of interest
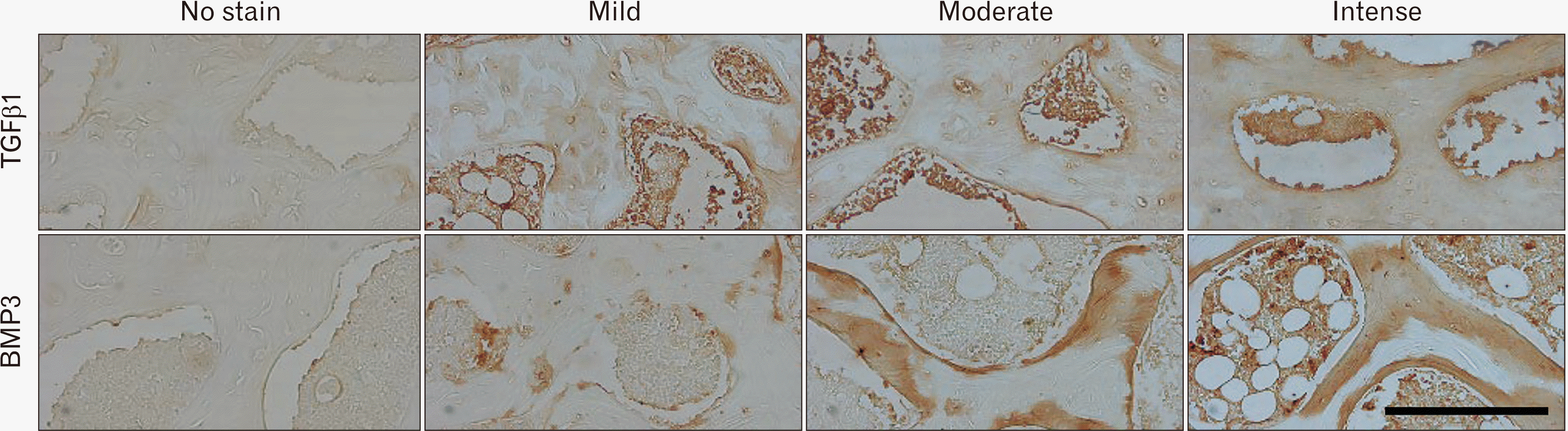
A light microscope (Zeiss Axioscope 2 plus) fitted with an Axiocam HRC digital camera connected to a PC was used to take photomicrographs of the stained slides. Anatomical landmarks were determined for the humerus head, using the H&E stained slides, to create consistency in quantification and immunopositivity analysis. Nonadjacent fields were selected, two above and two below the epiphyseal growth plate (Fig. 1). Photomicrographs were imported into Fiji Image-J software (National Institute of Health USA) for further analysis. TRAP positive osteocytes [15, 34], ALP positive osteoblasts [35, 36], AGEs positive bone cells and adipocytes were identified as specified in previous literature [31]. Cells were counted manually. Immunolocalization for anti-AGEs antibody, anti-BMP3 antibody, and anti-TGFβ1 antibody was performed. These were graded as no stain, mild, moderate, and intense immunopositivity (Fig. 1).
Data analysis
The data were managed in Microsoft Excel Office 365 (Microsoft Corporation) and all data were analyzed using SPSS® version 28 (IBM Co.). The data were tested for normality using the Shapiro–Wilks test, the data was found to be normally distributed, and a one-way ANOVA with LSD post-hoc test was applied for multiple group comparisons. Ordinal data for immunopositivity intensity were assigned grading scales of no stain (0), mild (1), moderate (2) and intense (3). Significance level was set at P<0.05.
RESULTS
Body mass
Animals in all the study groups showed a steady body mass gain up to week-17. By week-18, the ZDSD-MOD (530.31±22.50 g) and ZDSD-SVD (519.67±10.45 g) had a lower body mass compared to SD controls (540±12.18 g) (Fig. 2A). As such, significant differences were observed between SD and both ZDSD-MOD and ZDSD-SVD (P<0.001) from week 18 until termination. At 28 weeks the SDs (610.75± 20.66 g) had gained 70.63 g compared to both ZDSD-MOD (521.45±11.23 g) and ZDSD-SVD (490±20.15 g) that had lost 8.86 g and 29.47 g respectively (Fig. 2A).
Fasting blood glucose
The controls exhibited lower values compared to diabetics, with SDs (4.99±0.31 mmol/L) recording lower glucose levels compared to ZDSD-MOD (7.11±0.13 mmol/L) and ZDSD-SVD (13.38±1.61 mmol/L) (P=0.019, P<0.001 respectively) (Fig. 2B).
Oral glucose tolerance tests
The SD (4.70 mmol/L) controls exhibit lower glucose values at T0 compared to ZDSD-MOD (8.25 mmol min/dl) and ZDSD-SVD (11.20 mmol/L) (P=0.014, P<0.001 respectively) (Fig. 2C). This significant trend was maintained 2hrs post administration of the glucose bolus, with SD controls (6.53 mmol/L) showing lower values than ZDSD-MOD (12.97 mmol/L) together with the ZDSD-SVD (23.27 mmol/L) (P<0.001 for both comparisons) (Fig. 2C). The ZDSD-MOD (1,314±136.5 mmol min/dl) and ZDSD-SVD (2,434±320.60 mmol min/dl) showed an increase (71.2% and 217% respectively) in mean AUC when compared to SD (767.50±29.64 mmol min/dl) controls (Fig. 2C).
TGFβ1 and BMP3 expression
Most of the ZDSD-MOD and ZDSD-SVD animals exhibited mild immunopositivity. The remaining ZDSD-MOD and 1 ZDSD-SVD had moderate immunopositivity. The SD controls exhibited mostly mild, moderate (in equal proportions) with 1 animal showing intense immunopositivity (Table 2). This observation between the SD controls, ZDSD-MOD and ZDSD-SVD ZDSDs was statistically similar (P=0.206, P=0.103 respectively) (Table 2, Fig. 6).
All ZDSD-SVD rats exhibit intense BMP3 immunopositivity with majority of ZDSD-MOD exhibiting mild and moderate immunopositivity (Table 2). Most SD controls had mild immunopositivity with only 1 animal exhibiting no immunopositivity. Significant differences were observed between the SD controls and both groups of ZDSDs (P=0.002, P<0.001 respectively) (Table 2, Fig. 6).
DISCUSSION
Quantification of adipocytes revealed increased adiposity in the ZDSDs similar to other rat [37], and human studies [38]. TRAP and ALP immunopositive cells quantities showed that T2D promotes osteolysis and inhibits osteoblastogenesis in ZDSD rats. To understand the role of cytokines in the ZDSD rat skeletal elements, we assessed the expression of one osteogenic cytokine (TGFβ1) as well as an antagonist (BMP3). The ZDSDs had more BMP3 expression and relatively less TGFβ1. This shows that T2D affects bone cells though suppression TGFβ1 expression while its antagonist (BMP3) is elevated.
Anti-osteoblastogenic effects of adipocytes
Multiple studies have elucidated how T2D contributes to increased bone marrow adiposity. This adipogenic influence of T2D on mesenchymal stem cell enhances the development of adipocytogenic cell lines while suppressing osteogenic cell lines [31, 37, 39, 40]. These marrow adipocytes secrete the hormone adiponectin that is anti-osteoblastogenic, pro-osteoclastogenic and promotes osteoblast apoptosis [29, 40]. This could partially explain the susceptibility of bones to fracture in diabetes [31, 32]. In the present study, the ZDSDs exhibited higher adipocyte numbers compared to age matched controls. Our observation implies that T2D contribute to increased bone marrow adiposity [37] in the head of the humerus.
Osteoblastogenesis and osteolysis
To evaluate the proposition that osteocytes are active [15, 17, 41], and not dormant as previously thought [27], these cells were immuno-labeled using the anti-TRAP antibody. TRAP is typically used as an osteoclast marker; however, recent studies have shown that osteocytes express the bone matrix catabolic enzyme TRAP as they engage in osteolysis or peri lacuna-canaliculi remodeling [15, 41]. This causes direct resorption and deposition of the bone matrix in osteocytes’ immediate environment in response to pathologic conditions like T2D [15, 16, 34, 41]. In the present study, significantly more TRAP immunopositive osteocytes were observed in the ZDSDs and their age matched controls in alignment with previous research [4]. These TRAP immunopositive cells trigger processes that attract osteoclast precursors to their vicinity, leading to increased bone matrix resorption [17, 18], thus contributing to osteolysis.
Osteoblastic ALP promotes osteoid formation and bone mineralization [19]. It is used as a bone turnover marker in the diagnosis of bone disease [19, 42, 43]. Studies on animal models of T2D have reported impaired osteoblast differentiation, reduced osteoblast numbers and poor matrix mineralization in T2D. This is thought to lead to increased bone fragility [8, 20, 21]. In the present study, ZDSDs had fewer ALP positive osteoblasts. This finding supports the proposition that T2D inhibits osteoblastogenesis. This may further explain the known bone fragility in T2D.
Interaction of AGEs with TGFβ1 and its antagonist BMP3
Type 2 diabetes increases the expression of AGEs both intracellularly and in the extracellular matrix [44]. This AGEs accumulation increases with age and diabetes progression [44, 45]. It adversely affects intracellular signaling molecules that encode for osteoblast, osteoclast, and osteocyte functions [4, 26, 45, 46]. A possibility exists that this high AGEs accumulation could interfere with the TGFβ1 receptors, preventing entry of the extracelullar TGFβ1 into the cells, thereby interfering with the osteoblastogenic role of this cytokine.
The present study refers to the secreted cytokine rather than intracellular TGFβ1. Therefore, the extracellular TGFβ1 amounts may not be directly engaged in the cellular activities such as the well-known transcription for osteoprogenitor cell differentiation and function [22]. A possibility exists that the AGEs accumulation may block the interaction of TGFβ1 with cellular components required for osteoblastogeneis gene encoding [45].
In our study, ZDSDs exhibited either moderate or intense BMP3 immunopositivity, with no immunopositivity in the controls. This observation suggests that T2D reduces the expression of BMP3, and possibly suppresses osteoblast function [23, 47]. In support of this proposition, previous research reports increased osteoblast activity in BMP3 null mice [47]. Notably, intense immunopositivity was observed in the overtly diabetic group, possibly suggesting that BMP-3 over expression may be associated with excessive hyperglycemia.
In conclusion, we found that diabetes affected osteoblastogenesis as measured by ALP positive cells and bone marrow adipocytes. TRAP positive osteocytes numbers were increased in the presence of T2D, suggesting an increased osteolysis. The ZDSD groups had less osteogenic cytokine (TGFβ1) expression, with more of the antagonist (BMP3) expression. We also found that T2D increases the number of AGEs immuno-positive cells, as well as its extracellular expression, thus providing a conducive environment for the interaction of the osteogenic cytokine and its antagonist to suppress osteoblastogenesis. ZDSD groups had higher adipocyte numbers therefore increased marrow adiposity in T2D. We conclude that T2D adversely affects bone turnover cells, leading to reduced osteoblastogenesis and increased osteolysis the ZDSD rat humerus.

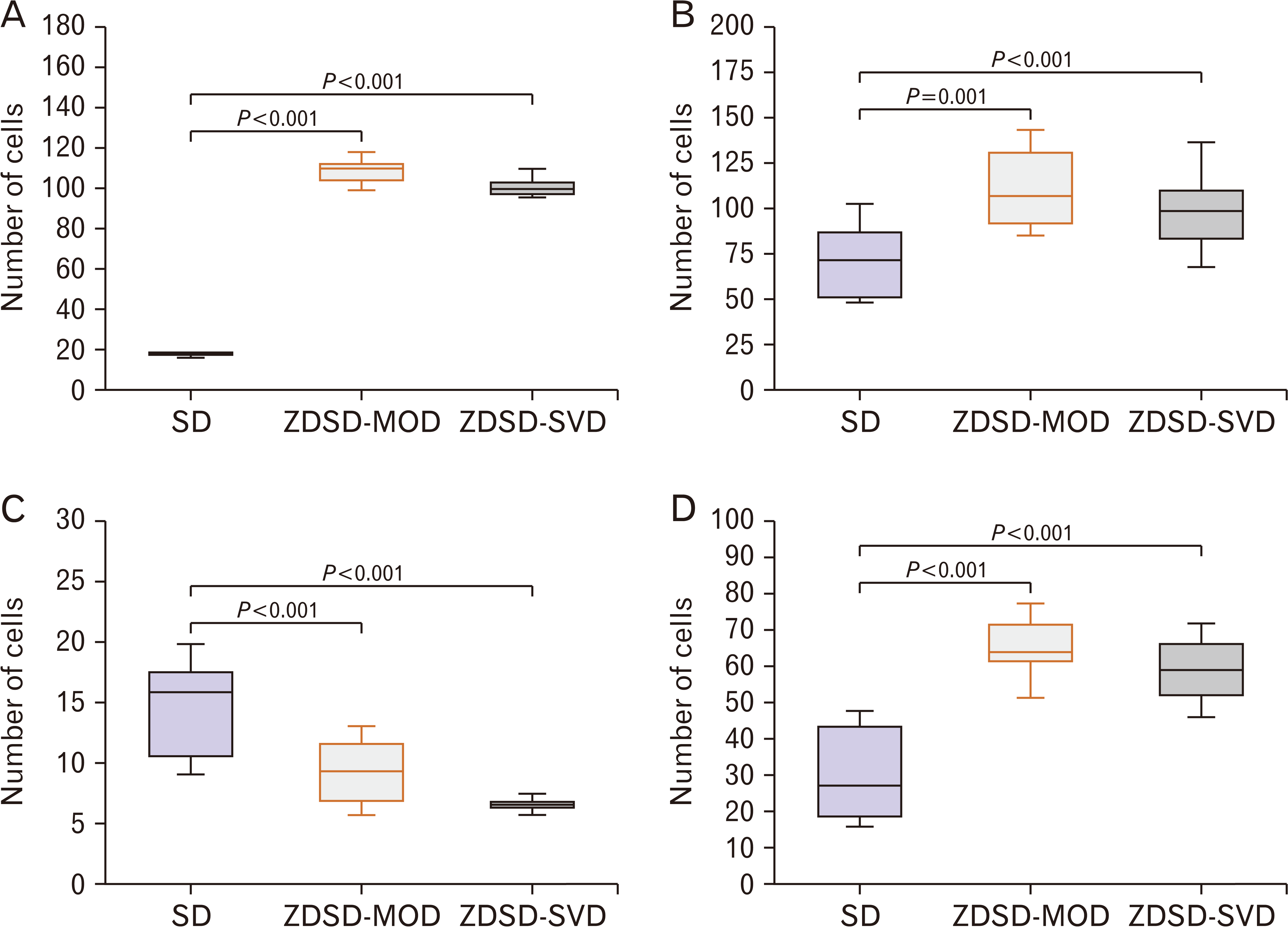
 XML Download
XML Download