

 PDF
PDF Citation
Citation Print
Print



Introduction
In the majority of cases with spermatogenic abnormalities, the causes of infertility could not be determined [1]. Anatomical malformations, gametogenesis dysfunction, endocrinopathies, immunologic difficulties, ejaculatory failure, environmental exposures, and gene mutations are among the causes of male infertility. Andrologists are particularly concerned with these concerns in relation to the diagnosis of male infertility [2, 3]. Semen parameters were discovered to be only the primary trump card; with them, we can only tell about the count and morphology; nevertheless, the deepest understanding of male infertility will emerge only when semen parameters are associated with a variety of molecular and biochemical parameters [4]. Septin4 (SEPT) are polymerizing GTP-binding proteins that are essential for a variety of cellular processes such as membrane compartmentalization, vesicle trafficking, mitosis, and cytoskeletal remodeling. So far, 14 distinct [5, 6]. Septin genes have been discovered for the past 35 years. Septin4 and its functions are disrupted in many human diseases, including neoplasia, breast cancer, Parkinson’s disease, neurogenerative disease, and male infertility [7, 8]. Infertile men are more likely to have sperm with aberrant expression patterns of Septin4, 7, and 12 in humans [9]. The annulus, a sub membranes ring that separates the middle and peripheral sections of the sperm flagella, is the Septin-rich part of sperm [10]. Septin’s function as an active GTPase or just a guanosine triphosphate binding protein is still debated [11, 12]. Septin provides far more energy, and the ring structure provides the circulatory force that propels sperm forward in the female reproductive system rather than remaining immotile [6, 13]. SEPTIN-4 is found in the annulus, a ring-like structure between the midpiece and the tail region of mature spermatozoa, along with other SEPTs [14]. Septin4 is required for the maintenance of normal mitochondrial architecture and the formation of the annulus during spermiogenesis [15]. SEPT4–/– mice were alive, but infertile in males due to asthenozoospermia and a bent neck [16]. SEPTIN-4 dis-localization from the annulus was also observed in immotile sperm with a faulty annulus [17]. Kwitny et al. [18] recently discovered that domain confinement was absent in SEPT4–/– mice sperm tails. Their findings support the idea that mammalian Septin structure plays a function in establishing a membrane diffusion barrier [9, 19]. Therefore, we investigated expression of Septin4 in the sperm of experimental individuals. In addition, we studied relationship of Septin4 expression with sperm parameters, level of malondialdehyde (MDA), and antioxidant enzymes, including total antioxidant capacity (TAC), catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPX).
Go to : 
Materials and Methods
Semen samples were collected from men attending the Infertility Research Center at the Academic Center for Education, Culture, and Research (ACECR) in Qom, Iran, between August 2019 and January 2020 for this prospective study. Three groups of sperm samples were created: Group I consisted of 20 normozoospermia-positive sperm samples. Twenty semen samples with asthenozoospermia were included in Group II. Twenty semen samples with astheno-teratozoospermia were included in Group III. All subjects were given a thorough medical history and physical assessment. A general medical examination was undertaken to look for abnormalities in the metabolic, endocrine, cardiovascular, respiratory, gastro-intestinal, and neurological systems that could affect fertility. All patients had a local examination of the testis, epididymis, vas deferens, and inguinal region to look for any abnormalities.
Ethical consent
Ethical clearance was done for this work to carry on human semen samples. Informed consent was also obtained from the patients in their own language. This study was approved by the Azad Medical University Ethics Committee for Research Involving Human Subjects (IR.QOM.REC.1399.003).
Assessment of sperm parameters
Fresh semen samples were taken by masturbation from cases after 2–7 days of sexual abstinence and allowed to liquefy for 15–30 minutes. Semen samples were washed by Ham’s F-10 solution. The resulting sperm pellet was divided into several aliquot parts and they were kept frozen at –80ºC for subsequent analyses of RNA and biochemical factor levels. The Computer Aided Sperm Analysis system (LABOMED, SDC313B; LaboMed Gerinnungszentrum) was used to determine sperm motility, which was classified as progressive, non-progressive, or immotile. Papanicolaou staining [20] was used to assess normal morphology, and individuals with fewer than 4% normal sperm morphology were classified as teratozoospermia according to World Health Organization (WHO) standards. Eosin-B staining was used to measure sperm viability (Merck). The dead sperm were stained red, whereas the living sperm were not. 100 sperm cells were counted in each sperm sample, and the percentage of viable sperm was computed [21]. Agnihotri et al. [22] used a technique to detect the mitochondrial membrane potential (MMP) of sperm. A fluorescent microscope (Olympus, DP71; Olympus Inc.) with a camera, a suitable filter, a magnification of 1,000×, 100 sperm counts, and sperms with natural MMP was used to extend a slide.
Sperm chromatin dispersion (SCD)
Aliquots of 0.2 ml of fresh sample semen diluted in medium to obtain sperm concentrations that ranged were between 5 and 10 million/ml. The suspensions were mixed with 1% low-melting-point aqueous agarose (to obtain a 0.7% final agarose concentration) at 37°C. Aliquots of 50 μl of the mixture were pipetted onto Coverslips were carefully covered, and slides were immediately immersed horizontally in a tray with freshly prepared acid denaturation solution (0.08 N HCL) for 7 minutes. A glass slide precoated with 0.65% standard agarose dried at 80°C, covered with a coverslip (24 by 60 mm), and left to solidify at 4°C for 4 minutes. The agarose matrix allows for work with unfixed sperm on a slide in a suspension like environment. At 22°C in the dark to generate restricted single-stranded DNA motifs from DNA breaks. The denaturation was then stopped, and proteins were removed by a transfer of the slides to a tray with neutralizing and lysing solution (0.4 M Tris, 0.8 M DTT, 1% SDS, 2 M NaCl, 0.05 M Triplex) for 25 minutes at room temperature. Removal of nuclear proteins results in nucleoids with a central core and a peripheral halo of dispersed DNA loop. Slides were thoroughly washed twice in water for 5 minutes, dehydrated in sequential 70%, 90%, and 100% ethanol baths (2 minutes each), and air dried. At the end cells were stained with Wright and PBS (1:1) for 10 minutes. After air dried, the degree of DNA dispersion was assessed by bright field microscopy. A minimum of 200 spermatozoa were evaluated by 2 different observers. In sperm with DNA fragmentation, the nucleus of the sperm is determined with a small halo and without a halo, and in sperms without DNA fragmentation, the sperm nucleus is determined with a large halo and a medium halo. In this study, 200 cells were counted to evaluated DNA fragmentation [23].
Assessment of semen biochemical factors
Seminal plasma was isolated and kept at –80°C until use for the biochemical factors analysis. All plasma samples were tested for TAC and MDA using commercial kits (Zell Bio GmbH). The ELIZA kit was used to measure the levels of SOD, CAT, and GPX (Abnova).
Quantitative reverse transcription-polymerase chain reaction
TRIzol reagent was used to extract total RNA from sperm samples. Sperm cell was mixed with 1 ml TRIzol, vortexed, and kept at room temperature for 5 minutes. It was then centrifuged for 15 minutes at 12,000 g, 4°C. The upper phase was separated into two tubes, one for RNA extraction and the other for protein extraction. Denaturing formaldehyde/MOPS/1% agarose electrophoresis was used to assess the integrity of total RNA. A spectrophotometer was used to verify the purity and concentration of the RNA and protein (Nano Drop Thermo Scientific). To remove genomic DNA, the RNAse-Free DNAse Set was used to perform a DNAase treatment. According to the manufacturer’s instructions, RNA was reverse transcribed using the Whole Transcriptome kit (Fermentase). The negative controls were not treated with reverse transcription (RT) enzyme, and all of the products were examined on a 4% agarose gel. To amplify the samples, a 750 real time polymerase chain reaction (PCR) system (Applied Bio System) was used with SYBR Green PCR master mix to perform quantitative RT-PCR (Q-PCR). The thermal cycling profile was as follows: 90 degrees Celsius for 10 minutes, 40 cycles of 90 degrees Celsius for 10 minutes, 60 degrees Celsius for 20 seconds, and 72 degrees Celsius for 20 seconds. On exons 7 and 8, the Septin4 amplicon was a 120 bp product. All reactions were duplicated three times. Sample Ct values were standardized to the expression of actin beta Ct values after regulating the amplification efficiency of the target genes. For the acquired data, the 2CT was calculated, and REST384- (2006) software was used to compare mean values between groups. The following gene-specific primers were used: Oligo, Macrogen provided us with gene-specific primers. The PCR amplification genes’ primer sets were chosen based on known sequences. The Septin4 gene primer produces a 322 bp fragment as follows: Antisense primer: 5-TGCACTGCTTCATCTCGAACTG-3; sense primer: 5-CCACAGCTATGAACCTGAGTCTGG-3. The glyceraldehyde-3-phosphate dehydrogenase gene has a primer produce a 382 bp fragment as follow: Sense primer: 5’-AATCCCATCACCATCTTCC-3’; Antisense primer: 5’-CATCACGCCACAGTTTCC-3’.
Statistical analysis
Clinical factors are reported as mean±SD, while Q-PCR results are presented as mean±SEM based on several studies. One-way ANOVA was used to assess statistical differences between groups. SPSS software was used to examine the data (version 19; IBM Co.). The quantitative information was presented as a mean and standard deviation. For the comparison of two groups, the Mann–Whitney test was utilized as a significance test. The qualitative information was displayed as a number and a percentage. To investigate the relationship between variables, the Spearman rank correlation coefficient was used. If the P-values are less than or equal to 0.05, the result is considered significant.
Go to :
Results
Semen parameters of studied groups
The first step was to categorize the semen samples based on the WHO values, this was done by using several semen samples, and each value and its error mean was the final mark. Based on the semen analysis report normozoospermia (n=20), asthenozoospermia (n=20), astheno-teratozoospermia (n=20). The semen parameter values were tabulated in Table 1. In the present study, results showed the sperm total motility (30±9.2 vs. 71±11.2; P=0.001), progressive motility (21±9.1 vs. 54±12.1; P=0.001), mean percentage of non-progressive motility (9±3.4 vs. 17±4.2; P=0.001), and immotile sperm (29±6.1 vs. 70±5.1; P=0.001) different between asthenozoospermia and normozoospermia groups. Astheno-teratozoospermia group the sperm total motility (25±7.1 vs. 71±11.2; P=0.001), progressive motility (18±8.1 vs. 54±12.1; P=0.001) significant decreased. But, the mean percentage of non-progressive motility (7±3.3 vs. 17±4.2; P=0.001), immotile sperm (85±6.3 vs. 29±6.1; P=0.001) and abnormal morphology (98±1.0 vs. 93±3.1; P=0.001) showed a significant increased. Also, there was highly significant decrease in viability (75±6.8 vs. 89±4.1; P=0.001 and 74±7.5 vs. 89±4.1; P=0.001), capacity (30.1±2.1 vs. 39.2±3.3; P=0.001 and 22.3±3.3 vs. 39.2±3.3; P=0.001) and MMP (60.13±2.494 vs. 75.73±3.494; P=0.001 and 55.44±3.09 vs. 75.73±3.494; P=0.001) in asthenozoospermia and astheno-teratozoospermia groups than normozoospermia group. The sperm DNA fragmentation index (DFI) was highly significantly decreased in asthenozoospermia (16±3.2 vs. 4.1±2.55; P=0.001), and astheno-teratozoospermia (26±2.44 vs. 4.1±2.55; P=0.001) compared with the normal group.
Table 1
Semen parameters of studied groups
| Parameters | Normozoospermia | Asthenozoospermia | Astheno-teratozoospermia | P-value |
|---|---|---|---|---|
| Concentration (million/ml) | 85.73±40.2 | 79±22.4 | 70.87±10.0 | NS |
| Total motility (%) | 71±11.2 | 30±9.2a) | 25±7.1a,b) | 0.001 |
| Progressive motility (%) | 54±12.1 | 21±9.1a) | 18±8.1a,b) | 0.001 |
| Non-progressive motility (%) | 17±4.2 | 9±3.4a) | 7±3.3a,b) | 0.001 |
| Immotile (%) | 29±6.1 | 70±5.1a) | 85±6.3a,b) | 0.001 |
| Abnormal morphology (%) | 93±3.1 | 95±2.1 | 98±1.0a) | 0.001 |
| Viability (%) | 89±4.1 | 75±6.8a) | 74±7.5a,b) | 0.001 |
| Capacity (%) | 39.2±3.3 | 30.1±2.1a) | 22.3±3.3a,b) | 0.001 |
| MMP (%) | 75.73±3.494 | 60.13±2.494a) | 55.44±3.09a,b) | 0.001 |
| DFI (%) | 4.1±2.55 | 16±3.2a) | 26±2.44a,b) | 0.001 |
![]()
Biochemical factors of studied group
A lower level of TAC on seminal plasma was observed in asthenozoospermia (1.82±0.11 vs. 2.51±0.13; P=0.01), and astheno-teratozoospermia (1.01±0.33 vs. 2.51±0.13; P=0.01) groups than normozoospermia group. In addition, the results demonstrated that CAT (14±1.2 vs. 18.04±1.79; P=0.01 and 11.3±1.7 vs. 18.04±1.79; P=0.01), GPX (331±8.741 vs. 378±13.25; P=0.01 and 303±9.145 vs. 378±13.25; P=0.01) and SOD (0.14±0.008 vs. 0.18±0.006; P=0.01 and 0.11±0.004 vs. 0.18±0.006; P=0.01) levels were significantly decreased in asthenozoospermia, and astheno-teratozoospermia groups compared with normal group. Moreover, the level of MDA on seminal plasma was significantly higher in asthenozoospermia group (2.21±0.01 vs. 1.97±0.09; P=0.01), astheno-teratozoospermia group (2.85±0.05 vs. 1.97±0.09; P=0.01) compared with the normal group (Table 2).
Table 2
Biochemical factors of studied group
| Biochemical factors | Normozoo-spermia | Asthenozoo-spermia | Astheno-teratozoospermia | P-value |
|---|---|---|---|---|
| TAC (μM) | 2.51±0.13 | 1.82±0.11a) | 1.01±0.33a,b) | 0.01 |
| MDA (μM) | 1.97±0.09 | 2.21±0.01a) | 2.85±0.05a,b) | 0.01 |
| CAT (U/ml) | 18.04±1.79 | 14±1.2a) | 11.3±1.7a,b) | 0.01 |
| SOD (U/ml) | 0.18±0.006 | 0.14±0.008a) | 0.11±0.004a,b) | 0.01 |
| GPX (U/ml) | 378±13.25 | 331±8.741a) | 303±9.145a,b) | 0.01 |
Values are presented as mean±SD. TAC, total antioxidant capacity; MDA, malondialdehyde; CAT, catalase; SOD, superoxide dismutase; GPX, glutathione peroxidase. a)Significance between norm group and asthenozoospermia, astheno-teratozoospermia; b)significance between asthenozoospermia group and astheno-teratozoospermia.
![]()
Septin4 gene expression
In this study, the transcript levels of Septin4 mRNA in sperm of patients were analyzed by Q-PCR between normozoospermia, asthenozoospermia, astheno-teratozoospermia patients. mRNA gene expression of Septin4 was significantly decreased in asthenozoospermia (P=0.019), and astheno-teratozoospermia (P=0.011) compared with the normal group (Fig. 1).
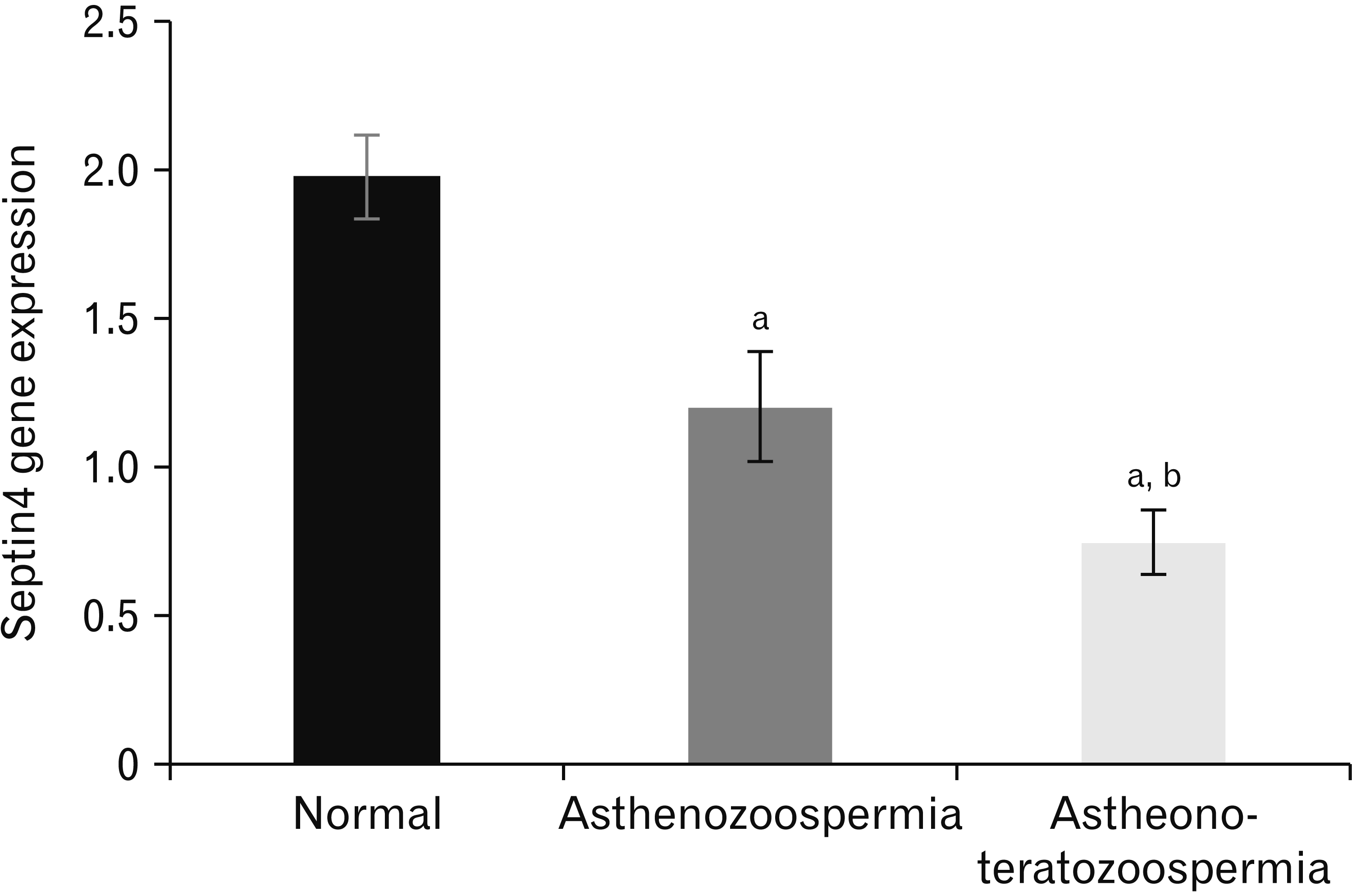 | Fig. 1Expression pattern of Septin4 was analyzed by real-time polymerase chain reaction. Level of mRNA gene expression of Septin4 was highly significantly decreased in asthenozoospermia, and asthenoteratozoospermia compared with the normal group. a, significantly difference with normal group; b, significantly difference with asthenozoospermia group (P<0.05).
|
Correlation of Septin4 mRNA with sperm parameters and biochemical factors
Our results in Table 3 show the correlation of Septin4 mRNA with sperm parameters of all studied groups. In this table, Septin4 mRNA expression shows highly significant (P<0.05) positive correlation with sperm concentration (rho=0.926), total motility (rho=0.698), progressive motility (rho=0.526), viability (rho=0.486), normal morphology (rho=0.633), capacity (rho=0.522), MMP (rho=0.537), and DFI (rho=–0.718). Table 4 show the positive correlation of Septin4 mRNA with TAC (rho=0.614), CAT (rho=0.607), SOD (rho=0.613), and GPX (rho=0.595). However, it shows significant (P<0.05) negative correlation with MDA (rho=–0.552).
Table 3
The correlation of Septin4 mRNA with sperm parameters
![]()
Go to :
Discussion
Septin4 gene expression was studied in human spermatozoa from normal and abnormal seminogram subjects (asthenozoospermia and astheno-teratozoospermia), and it was discovered that infertile groups have lower septin4 gene expression than normal seminogram subjects (fertile normal group). Septin4 deficiency or down regulation of the gene expression was found to cause a total or partial malfunction in the formation of the sperm flagellum, implying that it is important for forward motility [16, 24]. Septin4 was also found to be associated with basic sperm shape and function in our study (motility, morphology, viability, capacity, MMP, and DFI). This is always the result of a deficiency in the flagella or neck region [25, 26]. Other studies used Septin4-null sperm or flagella modified with no annulus, in which the annulus has been replaced by a narrow section missing cortical material, resulting in a hairpin-like structure [9, 27, 28]. The annulus of the spermatozoa has long been thought to have two important putative functions: one is a diffusion barrier function, which is critical for fertilization by retaining proteins in diverse compartments of the sperm tail to the neck [29, 30]. The second one could be on the morphological planner function, which directs flagellum growth and mitochondrial connection along the axoneme [31]. Septin4 null sperm were shown to be deficient in both of these pathways. The morphology of the human sperm annulus/flagellum has long been known, but the method by which it correlates is poorly understood. This result was consistent with that of Yeh et al. [32] Male SEPT4–/– mice are sterile due to sperm flagellum morphology and motility defects. The annulus is interrupted in SEPT4–/– null spermatozoa by a weak section lacking cortical substance. Septin4’s critical involvement in sperm head shape and nuclear DNA damage were proposed. The Septin4 related complex, which includes SEPT12, 6, and 7, is one of the explanations for the link between Septin4 gene expression and DNA damage [9]. Septin-4 expression is reduced, resulting in spermatid maturation arrest, fractured acrosomes, twisted tails, disorganized mitochondria, and nuclear DNA damage [33]. This finding is consistent with the discovery of malformed nuclei in SEPT4–/– mutant sperm, which could not fertilize oocytes but could be cured by injecting mutant sperm into oocytes, suggesting mutant sperm carry an intact haploid genome [34]. This is the first study to show that the expression level of Septin4 is linked to sperm stress oxidative markers, TAC, and antioxidant enzymes. Excess cytoplasm produces endogenous reactive oxygen species (ROS) in teratozoospermia patients with a proportion of aberrant sperm damage over the threshold of 96%, according to several studies [35, 36]. ROS is produced in these patients by a malfunction in the spermatogenesis process, which results in a large number of sperm with cytoplasmic residues [37]. When spermatozoa are exposed to ROS, DNA damage and lipid peroxidation can occur [38]. High lipid peroxidation can impair acrosome response, fertilization, and sperm-oocyte fusion [39]. Lipid peroxidation has been linked to poor sperm quality in several studies, affecting sperm quantity, motility, and morphology [40]. The relationship between oxidative stress and antioxidant enzyme may be causal and related to the molecular level in patients with (asthenozoospermia and astheno-teratozoospermia). Septin4 may potentially have a role in oxidative stress, which is caused by a nitric oxide (NO) and ROS imbalance. Angiogenesis is mediated and modulated by oxidative stress, which causes vascular endothelial damage [41]. Septin4 has been discovered as an oxidative stress factor that interacts with the apoptosis-related protein poly(ADP-ribose) polymerase 1 to cause oxidative vascular endothelial damage. Septin4 knock-down and overexpression have demonstrated [42]. Septin4 is also a physiological substrate of the ubiquitin ligase WW domain-containing protein 2 (WWP2), which plays a role in oxidative stress-induced vascular endothelial damage. WWP2 causes Septin4 to degrade, preventing the SEPT4/PARP1 complex from forming and suppressing endothelial damage and vascular remodeling [43]. Septin4 is thought to play a crucial role in motility and shape, and its absence may be linked to asthenozoospermia and astheno-teratozoospermia.
In conclusion, in the present study, we observed beneficial effect of Septin4, which improves sperm parameters, decreases MDA production and increases antioxidant enzyme levels, in addition to increasing Septin4 levels. Accordingly, normal human spermatogenesis requires an integrated antioxidant capability as reduced antioxidant enzyme levels may be attributed with defective sperm function. As a result of our findings, we came to the conclusion that Septin4 gene expression provides clinical useful information for the diagnosis of male infertility.
Go to :
 XML Download
XML Download