

 PDF
PDF Citation
Citation Print
Print



Introduction
Primary thyroid cancers exhibit substantial differences in their biological and clinical behaviors. This wide variability is not only rooted in genetic heterogeneity, but also in metabolic heterogeneity. Alterations in cellular energy metabolism are a major feature of metabolic heterogeneity in several cancers.1) Accumulating evidence has shown that cellular energy metabolism in remodeled tumors affects tumor behavior, which determines treatment sensitivity.2,3) Although thyroid cancers harbor genetic mutations that may provide the underlying cause of metabolic heterogeneity4,5) and altered cellular energy metabolism, the clinical significance of altered cellular energy metabolism remains to be explored.
Tumor cells, including thyroid cancer cells, exhibit a higher rate of glucose consumption than that required for adenosine triphosphate (ATP) synthesis.6) Tumor cells promote glucose entry by transporting glucose across glucose transporters (GLUTs) in the plasma membrane.7) Transported glucose is used for glycolysis to generate ATP and is also utilized for de novo serine biosynthesis for nucleotide synthesis and cell proliferation.7,8) Upregulation of serine, glycine, and one-carbon pathway (SGOCP) in tumors is known to be associated with poor prognosis. Serine is a major one-carbon donor to regulate the one-carbon pathway.9,10)
Serine availability though de novo serine biosynthesis plays or uptake from extracellular matrix is important to regulate the activities and functions of SGOCP. Phosphoglycerate dehydrogenase (PHGDH) is one of the rate-limiting enzyme in the de novo serine biosynthesis.10) PHGDH convert from 3-phosphoglycerate to 3-phosphohydroxypyruvate through an oxidation process, and then, phosphoserine amino transferase 1 (PSAT1) convert from 3-phosphohydroxypyruvate to 3-phosphoserine. Phosphoserine phosphatase (PSPH) finally dephosphorylates 3-phosphoserine to generate serine. PHGDH is highly activated in various tumors, including melanoma, colon, breast, and lung cancers, and is involved in tumor survival and cell proliferation. In thyroid cancer, PHGDH expression was known to be related with aggressiveness and cancer cell stemness.4) However, the role of PHGDH in glucose utilization and the metabolic flux of the serine biosynthetic pathway is not fully understood.
To answer this question, we characterized the role of de novo serine biosynthesis from glucose via PGHDH as an emerging target in papillary thyroid cancer. We hypothesized that elevated PHGDH levels are associated with high glucose uptake and also contribute to tumor aggressiveness in thyroid cancer.
Go to : 
Materials and Methods
Subjects
A total of 230 patients from Chungnam National University Hospital were enrolled in this study. The initial cancer diagnosis was confirmed through ultrasound-guided fine needle aspiration biopsy and was finally confirmed through full pathology review after thyroidectomy. Cancer diagnosis was performed by the World Health Organization (WHO) classification. All patients gave informed consent and this study was approved by the Institutional Review Board of the Chungnam National University Hospital (IRB-2020-11-004).
Immunohistochemistry
For immunostaining, we used a tumor microarray with the help of the Department of Pathology at Chungnam National University. Tissue samples obtained through surgery were first fixed in 10% formalin for 16 hours at room temperature. For immunostaining, tissues were dehydrated in graded ethanol, cleared in xylene, and embedded in paraffin wax. Paraffin-embedded tissue sections (4 μm thick) were incubated at 56°C for 3 h. We incubated tissue sections with anti-GLUT1 (Abcam) and anti-PHGDH (Cell signaling Technology) overnight at 4°C. Finally, UltraVision LP high-capacity detection system (Thermos Fisher Scientific, Waltham, MA, USA) was used to detect the expression of GLUT1 and PHGDH.
Gene Scores Using the Cancer Genome Atlas (TCGA) Database
Transcriptomic data for papillary thyroid carcinoma (THCA) collected as part of the TCGA project. We used the available data from the TCGA data portal (https://tcga-data.nci.nih.gov). Gene expression data from total 505 papillary thyroid cancer patients were analyzed. Gene scores calculated by the Gene set variation analysis (GSVA), the type of gene set enrichment method.
Cell Lines and Transfection
We get the cell lines from Professor Young Joo Park in Seoul National University. Human PTC cell lines (TPC-1, BCPAP, and KTC1) were maintained in Dulbecco’s Modified Eagle’s medium (DMEM; Invitrogen). Nthy-ori 3-1, an immortalized thyroid follicular epithelial cell line, were maintained in Roswell Park Memorial Institute (RPMI) medium (Invitrogen). We defined the low-glycemic condition as DMEM containing 5 mM D-glucose, and the high-glycemic condition as DMEM medium containing 25 mM D-glucose, and then cultured the cells. Each cell line was supplemented with 10% fetal bovine serum (FBS, Gibco, CA, USA), 100 U/mL penicillin and 100 g/mL streptomycin (Invitrogen) in a humidified incubator containing 5% CO2.
RNA Extraction and Quantitative PCR
TRIzol reagent (Invitrogen, USA) was used to get the RNA extraction. Complementary DNA (cDNA) was finally synthesized by M-MLV reverse transcriptase (Invitrogen, USA) and oligo-dT primers (Promega, USA). We analyzed the gene expressions, such as Glucose transporter 1 (GLUT1), PSAT1, PHGDH, and Hypoxia-inducible factor 1 A (HIF-1a), based on Quantitative PCR. cDNA, SYBR Green PCR mix (Applied Biosystems, USA), and specific primers were used for Quantitative PCR. Relative expression was calculated by normalization to GAPDH using a 7500 Fast Real-Time PCR System (Applied Biosystems).
Western Blot Analysis
Cells lysis were obtained using radioimmunoprecipitation assay (RIPA) buffer (150 mM sodium chloride, 1 mM sodium orthovanadate, 1 mM sodium phenylmethylsulfonyl fluoride, 30 mM Tris [pH 7.5], 1% Nonidet P-40, 10% glycerol, phosphatase and protease). Usually, 30-50 μg of protein were used. We used commercially available antibodies such as anti-PHGDH (Cell signaling Technology, USA), anti-GLUT1 (Abcam, USA), and anti-β-actin (Sigma-Aldrich, USA). The secondary antibody from Bio-Rad (Hercules, USA) were used to get the blot images. Images were acquired using the Odyssey imaging system. Image Studio Digits (LI-COR Biosciences, USA) were used to quantify the images.
DL-Serine Assay
For serine analysis, we analyzed both D- and L-serine levels in the cells using a DL-Serine Assay Kit (Abcam, Cambridge, UK). Cells were homogenized on ice with ice-cold serine assay buffer, centrifuged, and the supernatant was collected. To eliminate potential sources of interference, Sample Cleanup Mix was added to the samples during 15 min at 37°C. The samples were transferred to 10 kDa spin columns, centrifuged for 10 min, and the filtered samples were collected. The filtered samples were seeded in black, flat bottom 96 well plate. Samples were prepared in at least three sample wells for the measurement of D-serine only, total serine (both D- and L-serine), and sample background control. Detailed methods followed previous study.11)
Human PHGDH Small Interfering (si) RNA and PHGDH Inhibitor (NCT-503)
To explore the role of PHGDH, we explored the tumorigenesis using gene knock down system and PHGDH inhibitor (NCT503). Human PHGDH siRNA or negative scramble siRNA (siControl) from Thermo Fisher Scientific were used to knock down of PHGDH. Lipofectamine RNAiMAX transfection reagent (Invitrogen) were used. NCT-503 is a PHGDH inhibitor that reduces glucose-derived serine production and suppresses PHGDH-dependent cancer.12) Therefore, we determined whether cell migration was reduced under different glucose concentrations in thyroid cancer cells. We purchased NCT503 from Selleckchem company (USA).
Cell Migration Assay
Transwell chamber (Corning Costar, USA) with 6.5 mm diameter polycarbonate filters with 8 μm pore size were used to investigate the cell migration phenotype. The lower surface of the filter was coated with gelatin for the migration assay. The cells were fixed and stained with crystal violet. Chemotaxis was quantified by counting the number of cells that migrated to the lower surface of the filter using an optical microscope (×200 magnification).
Statistics
Student’s t-test or the Mann-Whitney U test were used to explore the statistical significance of between groups. Data were expressed as the as the mean±SD for and continuous variables. To explore the differences between groups the chi-square test were used. Statistical significance was set at a two-tailed p-value <0.05. All statistical analyses were performed with SPSS version 22.0 (IBM SPSS Statistics for Windows, version).
Go to :
Results
High Expressed PHGDH is Positively Correlated with GLUT1 Expression, and Related to Tumor Aggressiveness
To analyze the correlation between PHGDH and GLUT1, we stained tissues from patients with PTC. First, we found that PHGDH expression was higher in tumor samples than in normal samples (Fig. 1A, B). To identify the relationship between PHGDH and GLUT1 expression, 230 patients were enrolled in the study. Among the 230 patients, 56 showed no expression of PHGDH, and 174 patients were positive for PHGDH (Table 1). PHGDH expression were significantly associated with high tumor size (≥2 cm) (p=0.021), extracapsular invasion (p=0.017), central lymph node metastasis (p=0.001), lateral lymph node metastasis (p=0.006), and recurrence (p=0.036). Interestingly, we observed an association between PHGDH and GLUT1 expression (Fig. 1C). PHGDH expression was significantly associated with High GLUT1 expression (p=0.009). Our data suggest a link between the de novo serine biosynthetic pathway and glucose import in PTC and their impact on tumor aggressiveness.
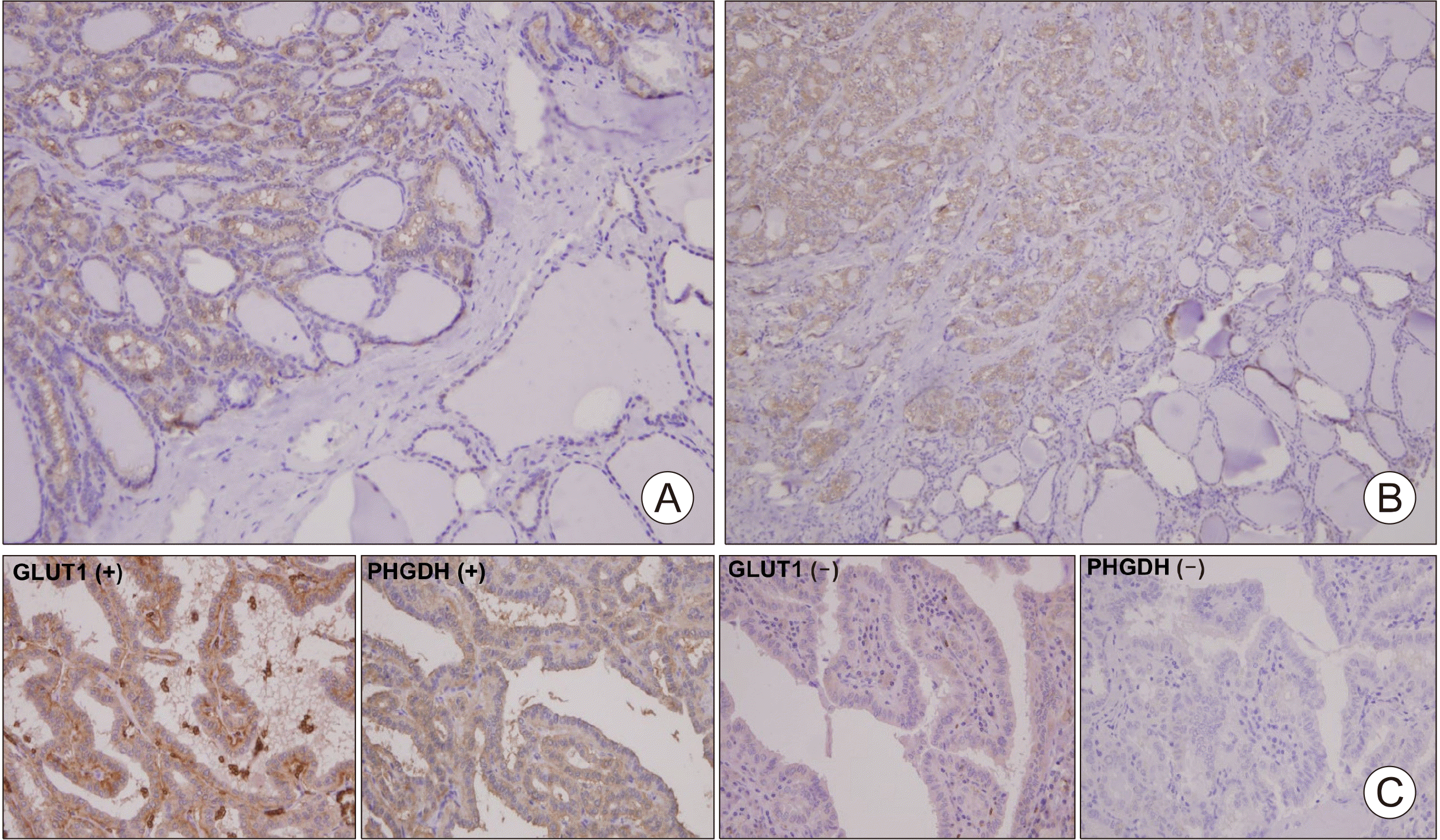 | Fig. 1Immunohistochemical evaluation of PHGDH and GLUT1 expression in papillary thyroid carcinoma (PTC) tissue. (A, B) Representative immunohistochemical images of PHGDH (×50). (C) Representative immunohistochemical images of PHGDH and GLUT1 (×100).
|
Table 1
Relationship between PHGDH expression and clinicopathologic factors in patients with thyroid cancer (n=230)
Data are presented as n (%) or mean±standard deviation unless otherwise noted. p-values were obtained from unpaired t-tests for continuous parametric variables and the Mann–Whitney U test for nonparametric variables. The chi-squared test and Fisher’s exact test were used to evaluate the significance of the correlations of PHGDH expression with clinical and pathological parameters. AJCC: American Joint Committee on Cancer, GLUT1: glucose transporter 1, ETE: extrathyroidal extension, LNM: lymph node metastasis, LVI: lymphovascular invasion, PHGDH: phosphoglycerate dehydrogenase
![]()
The Glucose Utility are Significantly Associated with de novo Serine Biosynthetic Pathway in Papillary Thyroid Cancer
Based on our observation that PHGDH and GLUT1 are upregulated in thyroid cancer tissues, we observed an association between glucose uptake and serine and glycine metabolism in thyroid cancer using TCGA database (Fig. 2A). Glucose import scores were significantly correlated with the de novo serine biosynthetic pathway (r=0.459, p-value<0.001), l-serine metabolic process (r=0.410, p-value<0.001), and one-carbon metabolic process (r=0.244, p-value< 0.001) (Fig. 2A). These results suggested that the metabolic flux of the serine biosynthetic pathway could be increased in relation to glucose import. Next, we found that the expression of PHGDH was increased in TPC-1, BCPAP, and KTC1 cells compared to in Nthy-ori 3-1 (Fig. 2B). To identify the role of PHGDH in glucose utilization, we cultured cells with different glucose concentrations in two groups, 5 mM glucose and 25 mM glucose. PHGDH and GLUT1 expression was increased under high-glucose conditions in the PTC cell lines (Fig. 2C). Moreover, glucose uptake significantly increased in both TPC-1 and BCPAP cells cultured in high-glucose media (Fig. 2D). L-serine and total-serine levels were also significantly increased by treatment with 25 mM glucose in both TPC-1 and BCPAP cells (Fig. 2E). Next, we investigated the expression of serine- and one-carbon metabolism-related genes, including PHGDH and GLUT1. High glucose condition significantly induced upregulation of GLUT1, PSAT1, PHGDH, and HIF1α in both TPC-1 and BCPAP (Fig. 2F-I). Our data suggest that in PTC, glucose utility is associated with the de novo serine biosynthetic pathway through the rate-limiting enzyme of the serine synthetic pathway, PHGDH.
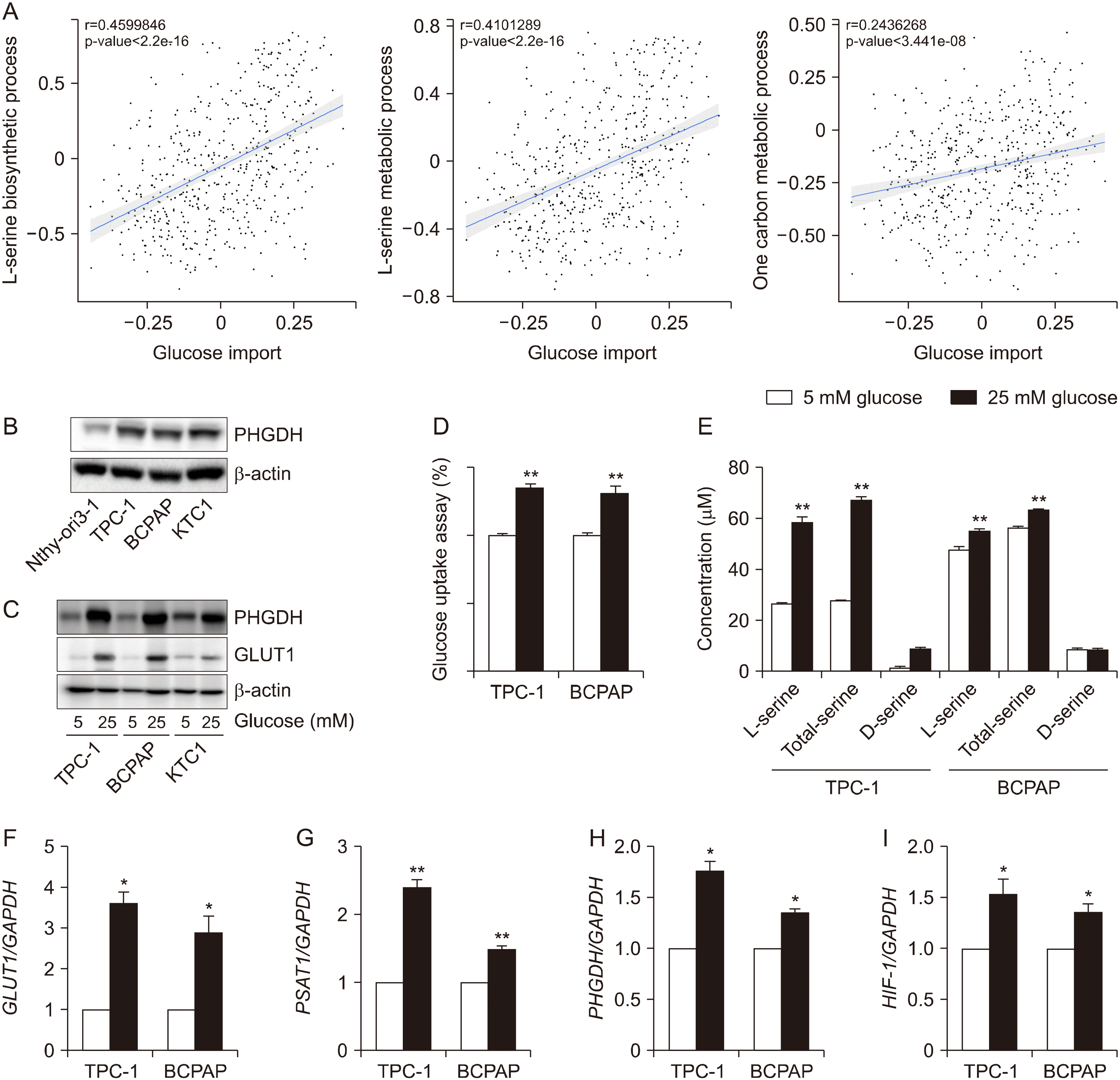 | Fig. 2The relationship of glucose import and serine biosynthetic pathway through PHGDH in papillary thyroid cancer. (A) The correlation plot of glucose import and serine related pathway using Gene score in TCGA-THCA cohort. (B) Representative Western blot images of PHGDH expression in thyroid cell lines. (C) Representative Western blot images of GLUT1 expression and PHGDH expression in thyroid cell lines in relation to glucose concentrations in media. (D) Glucose uptake in TPC-1 and BCPAP cells in relation to glucose concentrations in media. (E) Serine concentrations in TPC-1 and BCPAP cells in relation to glucose concentrations in media. (F-I) Real time PCR analysis of GLUT1, PSAT1, PHGDH, and HIF-1a in TPC-1 and BCPAP cells in relation to glucose concentrations in media. These experiments are representative of more than three independent experiments. *p<0.05, **p<0.01
|
The Effect of Downregulation or Inhibition of PHGDH were More Increased in High Glucose Utility Condition
To verify whether PHGDH affects tumor cell metastasis in vitro, transwell migration assays were performed using TPC-1 and BCPAP cells following PHGDH downregulation. PHGDH knockdown successfully induced a decrease in PHGDH levels in TPC-1 and BCPAP cells (Fig. 3A). In TPC-1 cells with low glucose, cell migration did not change; however, PHGDH knockdown significantly reduced cell migration in 25 mM glucose condition (Fig. 3B). In BCPAP, cell migration was decreased under high-glucose conditions compared to that under low-glucose conditions (Fig. 3C). To investigate the effect of pharmacological PHGDH inhibition using NCT-503, cells were treated with NCT503 for PHGDH inhibition. NCT-503 was slightly more reduces migration in TPC-1 under 25 mM glucose condition compared to 5 mM glucose (Fig. 3D). In addition, cell migration induced by NCT-503 in BCPAP cells was similar to that induced by TPC-1 (Fig. 3E). Thus, we verified that the pharmacological effect of PHGDH inhibition on the reduction of cell migration was related to glucose utility.
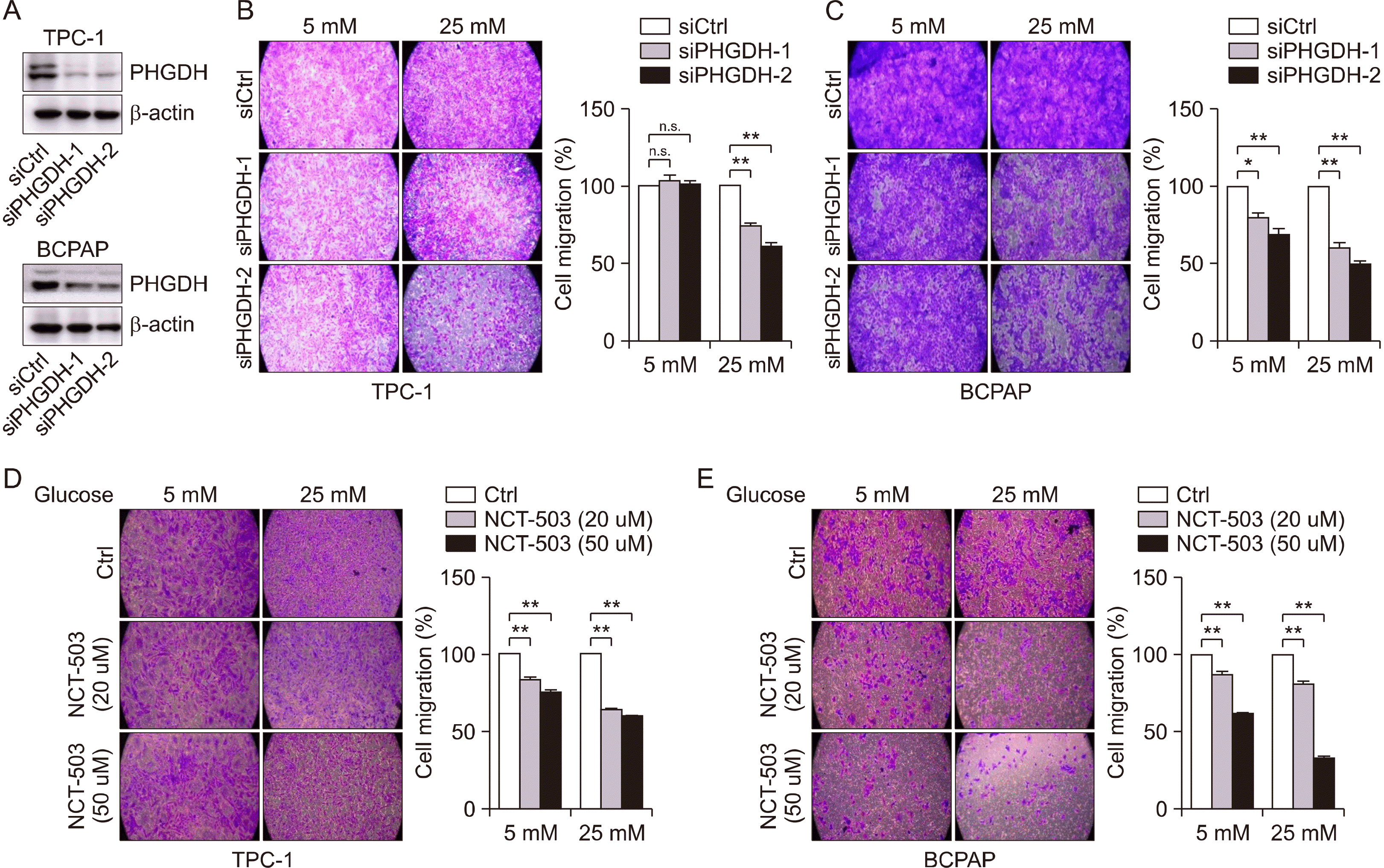 | Fig. 3Decrease of cell migration by siPHGDH or NCT503 in papillary thyroid cancer cell lines in relation to glucose utility. (A) Representative Western blot images of PHGDH expression in TPC-1 and BCPAP cells after knockdown of PHGDH. (B) Migration assay in TPC-1 cells with siPHGDH in relation to glucose concentrations in media. (C) Migration assay in BCPAP cells with siPHGDH in relation to glucose concentrations in media. (D) Migration assay in TPC-1 cells with NCT503 in relation to glucose concentrations in media. (E) Migration assay in BCPAP cells with NCT503 in relation to glucose concentrations in media. These experiments are representative of more than three independent experiments. *p<0.05, **p<0.01
|
Go to :
Discussion
In this study, we found that PHGDH expression is related to GLUT1, de novo serine synthesis is upregulated in high glucose conditions, and that it is a therapeutic target in papillary thyroid cancer. Cancer is increasingly considered a systemic disorder because it not only affects cells in localized tumors but also the whole body, especially in metastasis. In tumorigenesis, cancer-specific mutations and nutrient availability in the tumor microenvironment reprogram cellular metabolism to meet increased energy and biosynthesis needs.13,14) Recently, metabolic insights into thyroid cancer have been implicated in the diagnosis, prognosis, and therapeutic interventions because the thyroid gland is characterized by rapid energy metabolism for the regulation and synthesis of thyroid hormones.3) One of the most notable metabolic alterations observed in thyroid cancer is increased glycolytic activity, commonly known as the Warburg effect. Overexpression of glycolytic enzymes at the rate-determining steps, i.e. glucose import and phosphorylation, phosphofructokinase, and lactate export, increases glycolytic flux.15) In thyroid cancer, the upregulation of glycolytic enzymes, such as hexokinase, contributes to increased glycolysis.16,17) Moreover, 1H-NMR spectroscopy has been used to show that oxidative stress alters the secretion of various metabolites, such as lactate and amino acids.18) Thyroid cancer is known to overexpress hexokinase and has mitochondrial defects were contributed to upregulation of glycolysis.16,17) These results have been studied in oncocytic thyroid cancer, characterized by the accumulation of abnormal mitochondria in the cytoplasm. Recent study compared oncocytic and non-oncocytic thyroid cancer, and revealed higher glucose uptake and rerouting of glutamine to glutathione for redox homeostasis in the oncocytic cell line to compensate for its defective mitochondrial function.19) Moreover, 1H-NMR spectroscopy has been used to show that oxidative stress alters the metabolic profile in thyroid cancer, with increases in lactate and aromatic amino acids and a decrease in citrate.18) However, the de novo serine biosynthetic pathway from glucose is not fully understood in differentiated thyroid cancer.
Previous study explored the association of highly expression of PGHDH, SHMT2, and PSAT1 with poor prognosis in thyroid cancer.20) A recent study elucidated that PHGDH is involved in the cancer stemness in thyroid cancer using metabolomic profiling.4) Targeted metabolomics revealed altered glycolysis and de novo serine biosynthetic pathways in thyroid cancer cell lines.4) Moreover, PHGDH overexpression induces the expression of stemness markers. However, the association between glucose utility, such as glucose importation, and de novo serine biosynthesis is not fully understood. We found a correlation between glucose importation and the serine synthetic pathway that was confirmed in TCGA data, immunohistochemical analysis, and an in vitro study. High glucose concentrations increase glucose uptake and serine synthesis. Additionally, the inhibitory effect of PHGDH knockdown or PHGDH inhibition of PHGDH was more prominent in high-glucose media. Thus, our findings are consistent with previous studies and implicate the role of PHGDH in glucose utilization in papillary thyroid cancer.
Our study has some limitations. We could not directly identify glucose utility or serine synthesis based on the metabolomics method and performed in vitro experiments to investigate the role of PHGDH in thyroid cancer. To improve our knowledge, a precise metabolomics method and a large study, based on multiomics data are necessary. Despite these limitations, our data suggest the link of metabolic flux with tumor aggressiveness in thyroid cancer.
In conclusion, we found that serine biosynthesis from glucose is related with tumor aggressiveness in papillary thyroid cancer. PHGDH expression is significantly associated with glucose importation and tumor progression in thyroid cancer. Our findings suggest the possibility that cancer metabolism may represent a novel therapeutic strategy in thyroid cancer.
Go to :
 XML Download
XML Download