

 PDF
PDF Citation
Citation Print
Print



The Songwon Medical Scientist Award is the highest scientific award of the Korean Endocrine Society to honor an individual below 45 years old who has made excellent scientific contributions and planned outstanding research proposal to progress in the field of endocrinology and metabolism. The Songwon Medical Scientist Award is named after the pen name of Professor Kab Bum Huh, who had been emphasizing the importance of research and educations in field of not only endocrinology but also overall medicine. Professor Cheol Ryong Ku received the 1st Songwon Medical Scientist Award at the 10th Seoul International Congress of Endocrinology and Metabolism of the Korean Endocrine Society in October 2022.
INTRODUCTION
Approximately 15% of all primary intracranial tumors are pituitary neuroendocrine tumors [1]. Growth hormone (GH)-producing pituitary adenomas are the second most common hormone-producing pituitary tumors after prolactin-producing adenomas [1]. Acromegaly presents with various symptoms such as enlargement of the extremities and tongue, cardiac hypertrophy, osteoarthropathy, metabolic disorders, and malignancy due to prolonged excess GH [2,3]. If GH overproduction is not controlled, these complications can lead to high mortality rate [4,5].
Improvements in surgical procedures, the development of new radiotherapy techniques, and the availability of new medical therapies have resulted in considerable changes to the treatment and management of acromegaly [6,7]. However, a significant number of patients may not achieve remission owing to multiple clinical parameters [8-12]. Therefore, investigating the underlying biological mechanisms can help to identify targets for more effective treatments. This review outlines the need for a deeper understanding of patients with acromegaly to improve disease control and compliance.
Rather than examining individual genes, proteins, or metabolites, the focus shifts to analyzing complete sets: the genome, proteome, metabolome, and radiomics. These comprehensive approaches are categorized as “omics,” reflecting encompassing nature denoted by the suffix “-ome” [13]. Omics techniques are used in translational medicine, where research outcomes are translated into practical clinical applications, leveraging their inherent capacity for personalized medicine [14]. The impartial application of omics enables the evaluation of diagnosis, prognosis, and prediction of treatment responses, facilitating the customization of medical approaches, ranging from risk assessment to the handling of treatment resistance [15].
Despite recent efforts to elucidate the mechanisms and etiology of GH-secreting pituitary adenomas, the connection between genetic alterations and clinical characteristics remains unclear, and the factors influencing treatment responsiveness remain unknown. Moreover, genetic investigations specifically targeting GH-secreting pituitary tumors are limited. Therefore, a deeper understanding of the molecular basis of acromegaly is essential for developing targeted therapies and personalized treatment strategies.
In this review, we summarized the molecular aspects revealed by multiomics approaches that have shed new insight into the understanding of clinical behavior and novel therapeutic targets for patients with acromegaly and conclude with a discussion of future prospects.
GENOMICS
Among the various omics technologies, genomics, the most established domain, involves the investigation of linked genetic variations to discover novel therapeutic reactions or disease prognoses [16]. In this context, the impact of understanding genetic variations, as observed in genome-wide association studies (GWAS), coupled with omics discoveries, has been identified [17]. Genomic studies have identified several genetic alterations associated with acromegaly, including germline variants, somatic mutations, and copy number variations.
Germline mutations occur in several known genes, including those in multiple endocrine neoplasia type 1 (MEN1), protein kinase A regulatory subunit 1α (PRKARIA), aryl hydrocarbon receptor-interacting protein (AIP), G-protein coupled receptor 101 (GPR101), cyclin dependent kinase inhibitor 1B (CDKN1B), succinate dehydrogenase subunit (SDHx), MYC associated factor X (MAX) genes, as well as familial cases with currently unknown genes [18-21].
The most commonly identified somatic mutations in acromegaly are activating mutations in the guanine nucleotide-binding protein alpha-stimulating (GNAS) gene [22]. These mutations cause a gain-of-function effect in GNAS, leading to increased production of cyclic adenosine monophosphate (cAMP), which in turn promotes cell proliferation and GH release. Notably, specific mutations have been identified at critical positions, such as codon 201, where arginine can be substituted with cysteine, histidine, or serine, and codon 227, where glutamine can be replaced with arginine or leucine [23]. Approximately 30% to 40% of sporadic acromegaly cases exhibit GNAS mutations [22]. GNAS mutations in GH-secreting pituitary tumors are associated with higher preoperative insulin-like growth factor-1 (IGF-1) levels, higher surgical remission rates, favorable responses to somatostatin analogues, and lower immediate postoperative nadir GH levels [24].
Genomic stability of GH-secreting adenomas has been extensively studied. In a GWAS involving 128 sporadic pituitary adenomas, three susceptibility loci were identified, two on chromosome 10 and one on chromosome 13 [25]. In addition, copy number variations have also been reported. Interestingly, GH-secreting pitutiary tumors do not exhibit classical oncogene mutations when analyzed by whole-genome sequencing and somatic copy number analysis, indicating potential defects in cell cycle regulation or signaling pathways [26]. Furthermore, genomic profiling of 39 GH-secreting tumors revealed heterogeneity in copy number alterations, indicating genomic instability. Some tumors exhibit substantial genomic disruption and aneuploidy, affecting up to 45% of the affected genome [27]. These tumors also harbored GNAS1 mutations. In a multiplex next-generation sequencing analysis of 16 GH-secreting pituitary tumors, copy number variations were more prevalent in GH-secreting pituitary tumors than in non-hormone-secreting pitutiary tumors [28].
EPIGENOMICS
Epigenetic modification
Given the centrality of gene expression changes and the limitations of genetic abnormalities in GH-secreting pituitary tumors, epigenetic modifications have received considerable attention. Numerous genes involved in cell growth and signaling show altered methylation status, including cell cycle regulators such as cyclin dependent kinase inhibitor 2A (CDKN2A), CDKN1B, RB transcriptional corepressor 1 (Rb), and fibroblast growth factor receptor 2 (FGFR2) [29-31]. GH-secreting pituitary tumors display relative hypomethylation of genes involved in ion channel signaling, including voltage-gated potassium channel subunit beta-2 (KCNAB2), calcium-activated potassium channel subunit beta-4 (KCNMB4), and calcium voltage-gated channel subunit alpha1C (CACNA1C), which can be hypermethylated in nonfunctional tumors [32].
Increased global acetylation leads to increased expression of pituitary tumor-transforming gene 1 (PTTG1), bone morphogenic protein 4 (BMP4), and dopamine receptor 2 (DR2) in pituitary tumor cells. This suggests that widespread changes in epigenetic modifications could induce shifts in gene expression [33,34]. Overexpression of histone deacetylases (HDAC) within the sirtuin family (SIRT) have been observed in somatotropinomas and is correlated with smaller tumor size [35]. Peptidylarginine deaminase (PAD) enzymes plays a role in histone citrullination, a process that influences chromatin activity. The enhanced presence of PAD enzymes in somatoprolactinomas has been linked to increased mRNA targeting of the oncogenes high-mobility group A 1 (HMGA1), IGF-1, and the neuroblastoma MYC oncogene (N-MYC) by micro-RNAs (miRNAs) [36].
Epigenomic profiling can contribute to the understanding of tumorigenesis in pituitary adenomas. However, there are no significant differences in tumor subtype or prognosis.
Noncoding RNAs
Approximately 75% of the transcribed genes yield RNA molecules that do not participate in protein synthesis; instead, they generate noncoding RNAs [37]. These noncoding RNAs encompass miRNAs, long noncoding RNAs, circular RNAs, and P-element Induced WImpy testis in Drosophila (PIWI)-interacting RNAs [38]. Although their biological significance is not fully understood in most cases, several miRNAs have been shown to be involved in the control of pituitary tumorigenesis, aggressive features, and drug responsiveness.
miR-15a and miR-16-1 demonstrate decreased expression in GH-secreting pituitary tumors, and miR-16 is known to target GH receptor, IGF-1, IGF1 receptor, and IGF2 receptor expression [39,40]. Downregulated miRNAs were found to regulate the expression of three genes linked to proliferation: HMGA1, controlled by miR-34b and miR-548c-3p, HMGA2 regulated by miR-34b, miR-326, miR-432, miR-548c-3p, and miR-570; and E2 promoter binding factor 1 (E2F1), which is affected by miR-326 and miR-603 [41]. Overexpression of these three genes is associated with cell proliferation [41].
The dysregulation of miR-126, miR-381, and miR-338-3p controls cell proliferation, invasion, and response to therapy by affecting PTTG1 [42-44]. MiR-26b and miR-128 act on the phosphatase and tensin homologue (PTEN)/phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) pathway, which stimulates epithelial-mesenchymal transition (EMT) [45,46]. MiR-503, known as a tumor suppressor, was found to be downregulated, whereas miR-525-5p, a protector against EMT, was upregulated in GH-secreting adenomas. Dysregulation of both has been reported to control EMT during tumorigenesis [47,48].
In acromegaly, as observed in other tumors, miRNAs also play a role in resistance to treatment. For instance, miR-34a was shown to correlate with AIP expression, which is an important marker of response to somatostatin analogue treatment [49,50]. Somatostatin receptor (SSTR) 2 is regulated by miR-185 in vitro and is upregulated in non-responders to somatostatin analogues [51]. However, no direct correlation was observed between mir185 and SST2 expression in vivo. miR-125a-5p and miR-524-5p are decreased in response to somatostatin analogues [42]. miR-866-5p, a type of oncomiR, is upregulated In responsive tumors [42].
TRANSCRIPTOMICS
Transcriptomics, an additional high-throughput methodology, involves concurrent qualitative and quantitative analyses of specific mRNA species and focuses on dysregulated genes in acromegaly [52]. The integration of transcriptomic data provides a comprehensive understanding of tumor aggressiveness and treatment responsiveness in patients with acromegaly.
Signal transducer and activator of transcription 3 (STAT3) is a signaling molecule that has been implicated in GH hypersecretion by somatotrophic cells and therefore holds potential as a therapeutic target for GH-secreting pituitary tumors. GH-secreting tumors exhibit increased STAT3 expression, which leads to enhanced GH synthesis [53]. Blocking STAT3 activity inhibits the growth of somatotroph tumor xenografts and suppresses GH secretion in cultured cells derived from human GH-secreting pituitary tumors [53].
PTTG regulates cell cycle by interacting with p53 [54]. More than 70% of GH-secreting pituitary tumors exhibit elevated PTTG expression, which significantly contributes to cellular senescence [55]. PTTG expression may be indicative of aggressiveness in various types of pituitary adenoma [56].
Microarray analysis of GH-secreting pituitary tumors identified E-cadherin mRNA expression to be negatively associated with tumor size and invasiveness, and positively associated with GH and IGF-I levels in serum and response to somatostatin analogues [57]. Several genes, including epithelial splicing regulatory protein 1 (ESRP1), pleomorphic adenoma gene 1 (PLAG1) like zinc finger 1 (ZAC1), and beta arrestin-1, is reported as predictors of responsiveness to medical treatment [57-60].
Rapid polymerase chain reaction amplification of cDNA ends using human pituitary tumors identified a novel alternatively spliced somatostatin 5 (sst5) receptor variant [61]. The presence of truncated sst5 (sst5TMD4) is associated with increased aggressive features and worse prognosis in GH-secreting pituitary tumors [62]. Enhanced p21 expression is linked to less aggressive GH-secreting pituitary tumors and likely exerts a mitigating effect on cell proliferation [21].
Some studies have identified somatic genetic alterations in GH-secreting pituitary tumors using whole-exome sequencing. Whole-exome sequencing detected arm-level somatic copy number alterations (SCNA) in 42 pituitary macroadenoma samples at extensive sites across the genome in 29% of the specimens. Chromosomal alterations are more frequent in GH-secreting pituitary tumors [63,64]. Both cAMP and Fanconi anaemia DNA damage repair pathways are affected by SCNA in GH-secreting pituitary tumors [63]. Another study reported that in addition to the cAMP pathway, calcium signaling might be involved in the pathogenesis of GH-secreting pituitary tumors [64]. One study proposed novel somatic variants (48 genes with 59 variants) in patients with acromegaly with neither GNAS variants nor family history [65]. Single cell RNA sequencing of pituitary-specific positive transcription factor 1 positive pituitary adenomas (PIT1-PAs) proposed interferon-γ and cadherin 2-targeted drugs are promising therapeutic methods for aggressive PIT1-PAs [66].
PROTEOMICS
Proteomics is another omics-related technology that examines the protein content of an organism, tissue, or cell to understand the function or structure of a specific protein. This technology can be used in various research settings with different capacities to identify diagnostic markers, produce vaccines, and even interpret the protein pathways of disorders [67]. Herein, large-scale protein characterisation using this high-throughput proteomic strategy may benefit from mass spectroscopy techniques [68].
Although limited, proteomics studies have been conducted in patients with acromegaly. ATPase sarcoplasmic/endoplasmic reticulum Ca2+ transporting 2 (ATP2A2) and AT-rich interaction domain 5 B (ARID5B) correlated with the GH change rate in the octreotide loading test, and WWC family member 3 (WWC3), serine incorporator 1 (SERINC1), and zinc finger AN1-type containing 3 (ZFAND3) correlated with the tumor volume change rate after somatostatin analogue treatment [69]. One study evaluated biomarkersare changed after surgical treatment and revealed intensities of two isoforms of transthyretin, haptoglobin (HP) a2, haemoglobin β subunit (HBB) and two isoforms of apolipoprotein A1 (APOA1), and complement C4B (C4B) precursor could be used as potential biomarkers [70].
METABOLOMICS
Metabolomics is a rapidly growing technology that has identified perturbations in various metabolic pathways, including glucose, lipid, and amino acid metabolism. These findings may contribute to the development of novel diagnostic biomarkers and targeted therapies for acromegaly.
Feng et al. [7] reported that all pituitary adenomas, including GH-secreting pituitary adenomas, display downregulated glucose metabolism and glycolysis compared to normal tissues. They revealed that isocitrate dehydrogenase (NADP(+)) 2 (IDH2) is a key player in the reprogrammed metabolism of such tumors, and confirmed that IDH2 is a potential target for inhibiting tumor cell growth and secretion by blocking IDH2 [71].
RADIOMICS
Radiomics transforms radiological images into extractable data by automatically quantifying comprehensive imaging characteristics [75]. Previous studies have shown promising results in predicting the molecular state, severity, and outlook of gliomas [76]. Recent investigations have also shown encouraging findings for predicting the subtype or treatment response of pituitary adenomas. Although basic observations, such as cavernous sinus invasion on magnetic resonance imaging (MRI), are also associated with prognosis, advancements in MRI methodologies have led to a variety of ongoing radiomic studies [77,78].
Park et al. [79,80] proposed that radiomics features have the potential to predict the granulation pattern of GH-secreting pituitary tumors. Earlier studies have suggested that densely granulated adenomas respond more favorably to somatostatin analogue treatment than sparsely granulated adenomas do. This is particularly relevant, given the limited accessibility of electron microscopy in clinical settings.
CONCLUSIONS
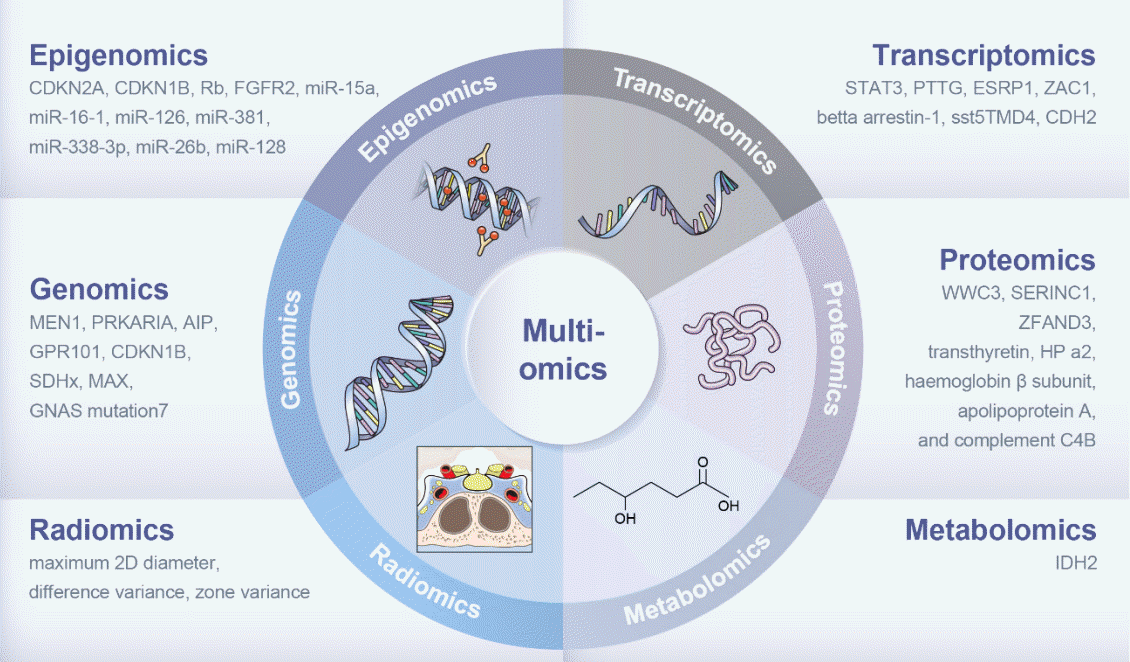
Nucleic acid-based methods, including genomics, epigenomics, and transcriptomics have been extensively used in patients with acromegaly, providing information on multiomics characteristics related to variable parameters such as drug responsiveness and recurrence (Fig. 1). To date, studies have focused on single-omics approaches to classify patients with acromegaly. Integrating various omics datasets offers a comprehensive perspective that enables us to derive more precise insights to understanding diseases and searching possible targets for treatment. Multiomics strategies have enlightened us in a manner that singular omics approaches alone cannot achieve. The integration of clinical data such as treatment response and disease outcomes through a multiomics approach can facilitate the identification of personalized treatment strategies. Further research and collaborative efforts are required to translate these findings into clinical practice.
 XML Download
XML Download