

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Retinal prosthesis is a promising treatment modality to restore vision by applying electrical stimulation to the retina of blind patients [1]. Retinitis pigmentosa (RP) is a common retinal disease that causes vision loss due to the death of photoreceptors [2]. Generally, the photoreceptors degenerate earlier, and bipolar cells and retinal ganglion cells (RGCs) survive longer, which provides an opportunity to bypass the damaged photoreceptor layer and stimulate surviving retinal neurons using retinal prostheses [3]. Despite some clinical success in restoring vision in RP patients with retinal prostheses, post-implantation visual acuity (a commonly used measure of spatial resolution in the visual processing system) still falls below 20/200, the legal definition of blindness [4]. Therefore, developing electrical stimulation strategies that can improve the spatial resolution of retinal responses is needed to improve the visual acuity of retinal prostheses.
Here, we propose an electrical stimulation strategy that elicits spatially localized RGC responses. Previous studies have shown a trade-off between increasing RGC firing and spatial resolution [5,6]. For example, increasing the intensity of the electrical stimulation would increase the number of spikes in RGCs around the stimulation electrode. However, the current spreading also causes distant untargeted RGCs to be stimulated, reducing the spatial resolution. Alternatively, decreasing the stimulation intensity reduces the number of all RGC spikes. Therefore, there is a need to develop a stimulation strategy that can elicit the focal response of target RGCs.
Selectively stimulating target RGCs is even more difficult for the degenerate retina of RP patients. As retinal degeneration progresses, retinal degenerate (RD) RGCs produce fewer spikes compared to wild-type (WT) RGCs at the same stimulation intensity [7,8]. Applying stronger stimulation to RD RGCs, such as increased pulse amplitude or pulse duration, can bring their responses up to those of WT RGCs. However, this approach inevitably leads to a decrease in spatial resolution. In addition, in RD retinas, retinal remodeling causes excessive connectivity between retinal neurons, resulting in lower spatial resolution of RD responses than WT responses [6,9]. Previous studies have yet to examine the efficient stimulation strategy of RD RGCs with high spatial resolution.
In this study, we propose a novel stimulation method to improve spatial resolution by 1) selectively increasing the firing of target RGCs and 2) reducing the stimulation of other untargeted RGCs. The latter is achieved by keeping the pulse amplitude per pulse low, just above the stimulation threshold. The former is achieved by introducing multiple consecutive low-amplitude pulses to the retina within a short period. We call this stimulation method ‘burst stimulation.’ In this study, we compared the RGC response of WT and RD retinas to the conventional single high-amplitude pulse, with burst stimulation; multiple consecutive low-amplitude pulses. The trade-off between RGC firing efficacy and spatial resolution can be quantified by varying the number of pulses of the burst stimulation.
Go to : 
METHODS
Retinal preparation
We used two types of mice: normal retinal model, WT C57BL/6J strain (n = 22); RD model, C3H/HeJ (rd1) (n = 22). All mice at postnatal day 56 (P56) were purchased from Jackson Laboratories (The Jackson Laboratory). At P56 in rd1 mice, the retinas are no longer responsive to light, but the functional stability of the inner retinal neurons is well preserved [10]. The C57BL/6J mouse model is commonly used in vision research to investigate various retinal functions, including object discrimination, motion detection, and light adaptation [11-13]. The C57BL/6J mouse model has been extensively used to study normal RGC responses to electrical stimulation [9,14,15]. Our experiments on C57BL/6J mice aimed to validate the efficacy of burst stimulation before introducing it to degenerate retinal conditions.
The C3H/HeJ (rd1) mouse model serves as a representative RD model of RP, a degenerative disorder characterized by photoreceptor loss. The rd1 mouse model results from a congenital genetic mutation affecting the Pde6b gene, crucial for photoreceptor phototransduction [16]. Importantly, rd1 mice undergo rapid photoreceptor degeneration by P28, leading to complete photoreceptor layer loss and blindness by P56 [10]. This model closely mimics the degenerate retinal environment in RP patients with early-onset vision loss due to genetic abnormalities. The rd1 mouse model has been extensively used to study degenerate RGC responses to electrical stimulation [14,15,17]. This study further applied burst stimulation to the rd1 retina, aiming to assess its potential to enhance the spatial resolution of RGC responses in degenerate retinas.
Animal use protocols were approved by the Institutional Animal Care Committee of Chungbuk National University (approval number: CBNUA-1708-22-01). All procedures followed the guidelines of the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research.
Mice were anesthetized by intramuscular injection of 30 mg/kg tiletamine-zolazepam hydroxide (Zoletil 50; Virbac), 10 mg/kg zylazine hydrochloride (Rumpun; Bayer Korea), and 5,000 IU heparin sodium (Heparin; JW Pharmaceutical Corp.). The detailed procedures for the preparation of ex vivo retinal patches have been described in a previous study [18]. Briefly, after eye enucleation, the retina was isolated from the sclera and retinal pigment epithelium (RPE) and cut into 2 × 2 mm2 patches. The retinal patch was prepared under illumination of 4.3 nW/cm2 in an artificial cerebrospinal fluid solution (ACSF) (124 mM NaCl, 10 mM glucose, 1.15 mM KH2PO4, 25 mM NaHCO3, 1.15 mM MgSO4, 2.5 mM CaCl2, and 5 mM KCl) bubbled with 95% O2 and 5% CO2 to maintain a pH of 7.3–7.4 and a temperature of 32°C. The isolated retina was mounted on the RGC layer on a planar multi-electrode array (MEA) and continuously perfused with oxygenated solution (flow rate: 1–3 ml/min) during the experiment.
Multielectrode recording system and signal processing
The data acquisition system (MEA60 system; Multichannel Systems GmbH) included a planar 64–channel perforated MEA (60pMEA200/30iR), an amplifier (MEA1060), temperature control units (TC01), data acquisition hardware (Mc_Card), and software (Mc_Rack). The MEA contained 64 circular electrodes in an 8 × 8 grid layout with electrode diameters of 30 µm and interelectrode distances of 200 µm. The electrodes were coated with porous titanium nitride and embedded in a perforated polyimide foil that provided sufficient oxygen and nutrient supply to the retina. Multielectrode recordings of retinal activity were obtained from 59 out of 64 electrodes, except for one reference electrode and four inactive electrodes with a bandwidth ranging from 1 to 3,000 Hz at a gain of 1,200. The data-sampling rate was 25 kHz for each electrode. From the raw waveform of the retinal recording, RGC spikes were isolated using a 100–Hz high-pass filter. The threshold for spike detection was set to four times the standard deviation of background noise. The recorded data were processed with spike sorting software (Offline Sorter; Plexon Inc.) for each MEA channel to separate multiunit activities containing different spike waveforms into individual cell units using principal component analysis [19]. In summary, we categorized RGCs based on distinct action potential (spike) waveforms. Using 30 µm diameter electrodes, about three RGCs are typically identified at one electrode, each showing diverse spike waveforms. Principal component analysis was used to classify these distinct shapes into separate RGCs by grouping similar spike waveforms, aiding the identification of three distinct RGCs.
Electrical stimulation
The RGC layer, not the retina's photoreceptor layer, was mounted on the MEA; therefore, our electrical stimulation mimicked the epiretinal prosthesis. Using a stimulus generator (STG 1004; Multichannel Systems GmbH), the current pulse train was delivered to the retinal preparation through one of the 60 channels (mostly channel 44 in the middle of the MEA), with the remaining channels serving as recording electrodes. The remaining channels of the MEA were binned into six groups (200–400, 400–600, 600–800, 800–1,000, 1000–1,200, and 1,200–1,400 µm) based on the distance between the stimulation and recording electrodes of the MEA. Four types of electrical stimulus protocols were used in this study.
Protocol 1: Increasing the pulse amplitude of a single pulse: Stimulus protocol 1 consisted of symmetrical cathodic phase 1st biphasic pulses. The 1st phase-pulses regulate the RGC response, and the 2nd phase-pulses are used to balance the previously applied charges for preventing electrode erosion and retinal tissue damage through charge accumulation [20]. Pulse duration was fixed at 250 µs/phase, and pulse amplitudes of 10, 20, 30, 40, and 50 µA/phase were applied (Fig. 1A). Thus, the stimulus charge applied to the retina was 2.5, 5.0, 7.5, 10.0, and 12.5 nC/phase. For each pulse amplitude, biphasic current pulses were applied once per second and repeated ten times. Stimulus protocol 1 was used in Fig. 2 and 3.
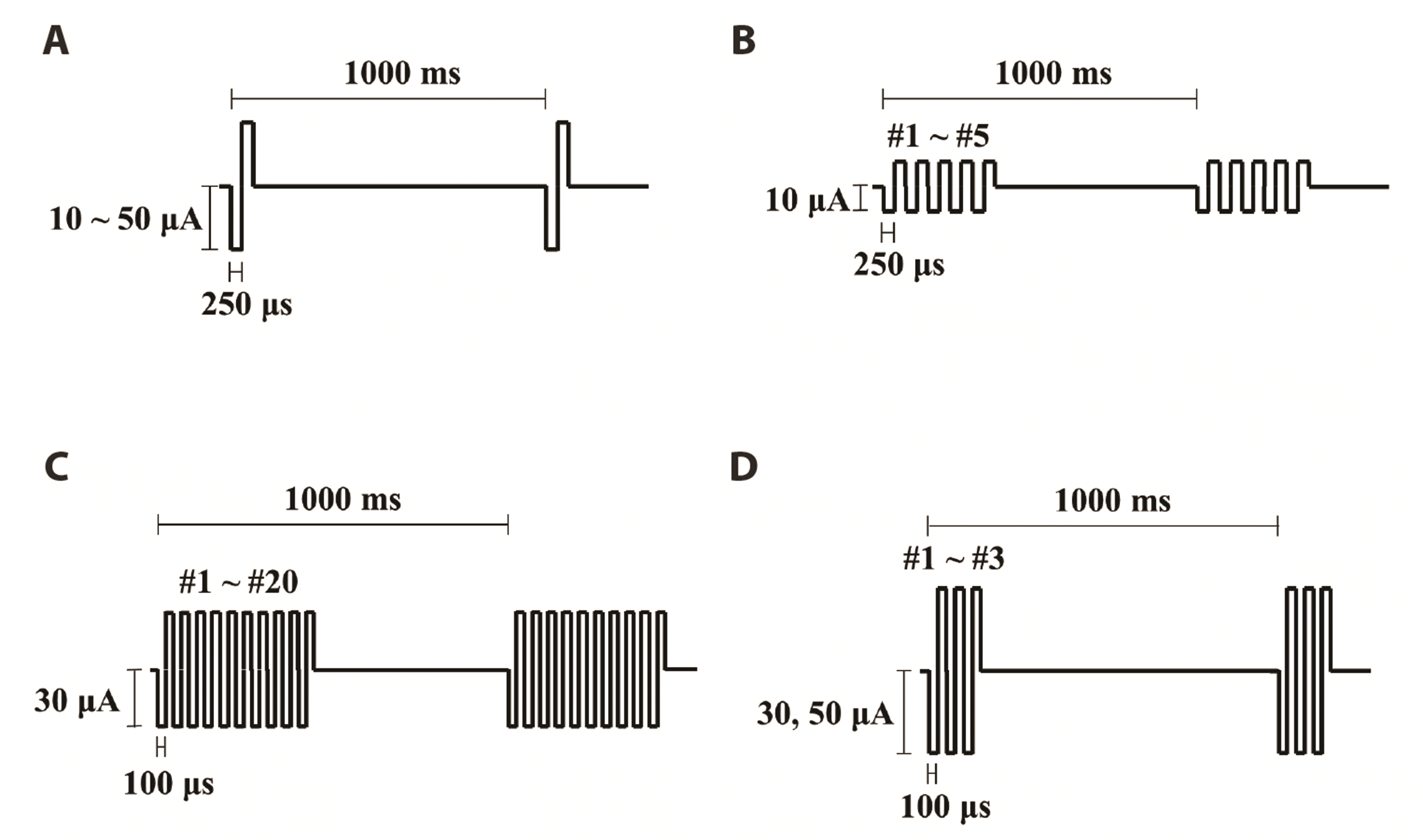 | Fig. 1Stimulus protocols used in the experiment.(A) Single-pulse stimulation. Pulse amplitudes range from 10 to 50 μA with pulse duration fixed at 250 μs are applied once per second. (B) Burst stimulation. The number of pulses per burst ranges from 1 to 5, with pulse duration fixed at 250 μs. (C) Burst stimulation. 1–20 pulses per burst. Pulse amplitude: 30 μA. Pulse duration: 100 μs. (D) Burst stimulation. 1–3 pulses per burst. Pulse amplitudes: 30 or 50 μA. Pulse duration: 100 μs.
|
 | Fig. 2Comparison of electrically-evoked RGC spikes between single-pulse and burst stimulation in the WT retina.(A) Left inset: MEA positions of the stimulation (channel 26, marked with a red dot) and recording (channel 36, marked with a yellow dot) electrodes spaced 200 μm apart. Left panel: Typical responses of the WT RGC to single-pulse stimulation are shown as raster plots. Electrically-evoked RGC spikes are highlighted. The time window for the raster plots is set to ± 0.1 sec, with 0 sec representing the stimulus onset (red dashed line). For each raster plot (10 trials), the pulse amplitude increases from top to bottom (10–50 μA). The pulse duration is fixed at 250 μs/phase. Middle panel: Typical responses of the WT RGC to burst stimulation are shown as raster plots. For each raster plot, the pulse number increases from top to bottom (#1–#5). Right panel: RGC response curves to single-pulse and burst stimulation in WT RGCs located 200 μm away from the stimulation electrode. Error bars represent the standard error of the mean (SEM). Statistical differences in response curves are indicated by asterisks (*p < 0.05). (B) Left and Middle panels: same representative raster plots of WT RGCs located 600 μm away from the stimulation electrode. Right panel: RGC response curves to single-pulse and burst stimulation in WT RGCs located 600 μm away from the stimulation electrode. Statistical differences in the graphs are indicated by asterisks (*p < 0.05, ***p < 0.001). (C) Changes in normalized responses with the distance between stimulation and recording electrodes. Left panel: single-pulse stimulation. The inset (spatial MEA color map) shows the normalized RGC responses in one representative retinal patch upon electrical stimulation with a pulse amplitude of 50 μA. The red dot indicates the location of the stimulation electrode. Right panel: burst stimulation. The inset (spatial MEA color map) shows the normalized RGC responses in one representative retinal patch upon electrical stimulation with five pulses per burst. RGC, retinal ganglion cell; WT, wild-type; MEA, multi-electrode array.
|
 | Fig. 3Comparison of electrically-evoked RGC spikes between single-pulse and burst stimulation in the RD retina.(A) Left inset: MEA positions of the stimulation (channel 52, marked with a red dot) and recording (channel 53, marked with a yellow dot) electrodes spaced 200 μm apart. Left panel: Typical responses of the RD RGC to single-pulse stimulation are shown as raster plots. Electrically-evoked RGC spikes are highlighted. The time window for the raster plots is set to ± 0.1 sec, with 0 sec representing the stimulus onset (red dashed line). For each raster plot (10 trials), the pulse amplitude increases from top to bottom (10–50 μA). The pulse duration is fixed at 250 μs/phase. Middle panel: Typical responses of the RD RGC to burst stimulation are shown as raster plots. For each raster plot, the pulse number increases from top to bottom (#1–#5). Right panel: RGC response curves to single-pulse and burst stimulation in RD RGCs located 200 μm away from the stimulation electrode. Error bars represent the standard error of the mean (SEM). Statistical differences in response curves are indicated by asterisks (*p < 0.05, **p < 0.01). (B) Left and Middle panels: same representative raster plots of RD RGCs located 600 μm away from the stimulation electrode. Right panel: RGC response curves to single-pulse and burst stimulation in RD RGCs located 600 μm away from the stimulation electrode. Statistical differences in the graphs are indicated by asterisks (*p < 0.05, **p < 0.01). (C) Changes in normalized responses with the distance between stimulation and recording electrodes. Left panel: single-pulse stimulation. The inset (spatial MEA color map) shows the normalized RGC responses in one representative retinal patch upon electrical stimulation with a pulse amplitude of 50 μA. The red dot indicates the location of the stimulation electrode. Right panel: burst stimulation. The inset (spatial MEA color map) shows the normalized RGC responses in one representative retinal patch upon electrical stimulation with five pulses per burst. RGC, retinal ganglion cell; RD, retinal degenerate; MEA, multi-electrode array.
|
Protocols 2, 3, and 4: Burst stimulation: Stimulus protocol 2 was fixed at a pulse duration of 250 µs/phase, and a pulse amplitude of 10 µA/phase (Fig. 1B). Pulse conditions consisting of 10 µA amplitude and 250 µs duration are stimulation thresholds for RGC firing, which are used to minimize the destruction of spatial resolution by current spreading [14,21]. Multiple consecutive-biphasic pulses were applied once per second. We call this protocol burst stimulation. The number of consecutive pulses applied during burst stimulation ranged from 1 to 5, matching the amount of injected charge for each pulse amplitude (2.5, 5.0, 7.5, 10.0, and 12.5 nC) used in protocol 1. Two consecutive biphasic pulses are called a doublet, three as a triplet, four as a quadruplet, etc. For each pulse number, biphasic current pulses were repeated ten times. Stimulus protocol 2 was used in Fig. 2 and 3.
Stimulus protocol 3 was fixed at a pulse duration of 100 µs/phase and a pulse amplitude of 30 µA/phase (Fig. 1C). Pulse conditions consisting of 30 µA amplitude and 100 µs duration is the stimulation threshold for RGC firing [14,21]. Multiple consecutive-biphasic pulses were applied once per second. The number of pulses per burst applied during burst stimulation varied from 1 to 20. For each pulse number, biphasic current pulses were repeated ten times. Stimulus protocol 3 was used in Fig. 4 and 5.
 | Fig. 4Increasing the number of pulses per burst in the WT retina.(A) Typical responses of the WT RGC to burst stimulation are shown as raster plots. Electrically-evoked RGC spikes are highlighted. The time window of the raster plot is set to ± 0.1 sec, with 0 sec representing the stimulus onset (red dashed line). In each raster plot (10 trials), the pulse number increases from top to bottom (#1–#5, #9–#13, #16–#20). The pulse amplitude and pulse duration are fixed with 30 μA and 100 μs/phase. Only RGCs closest 200 μm to the stimulation electrode were analyzed. (B) RGC response curves as a function of pulse number. Error bars represent the standard error of the mean (SEM). Statistical differences in the graphs are indicated by asterisks (*p < 0.05). RGC, retinal ganglion cell; WT, wild-type.
|
 | Fig. 5Increasing the number of pulses per burst in the RD retina.(A) Typical responses of the RD RGC to burst stimulation are shown as raster plots. Electrically-evoked RGC spikes are highlighted. The time window of the raster plot is set to ± 0.1 sec, with 0 sec representing the stimulus onset (red dashed line). In each raster plot (10 trials), the pulse number increases from top to bottom (#1–#5, #9–#13, #16–#20). The pulse amplitude and pulse duration are fixed with 30 μA and 100 μs/phase. Only RGCs closest 200 μm to the stimulation electrode were analyzed. (B) RGC response curves as a function of pulse number. Error bars represent the standard error of the mean (SEM). Statistical differences in the graphs are indicated by asterisks (*p < 0.05). RGC, retinal ganglion cell; RD, retinal degenerate.
|
Stimulus protocol 4 consisted of pulse amplitudes and durations of 30 µA, 100 µs or 50 µA, 100 µs (Fig. 1D). The number of pulses per burst applied during burst stimulation varied from 1 to 3. For each pulse number, biphasic current pulses were applied once per second and repeated ten times. Stimulus protocol 4 was used in Fig. 6.

Pharmacological treatment
Drug solutions were applied via perfusion for at least 30 min before recording. To increase cytosolic calcium concentration at the presynaptic terminals of bipolar cells, we used 10 µM thapsigargin (Th) (T9033; Sigma-Aldrich), which blocks the sarco/endoplasmic reticulum Ca2+–ATPase (SERCA) involved in the calcium influx from the cytoplasm into the endoplasmic reticulum [22]. To lower the cytosolic calcium concentration at the presynaptic terminals of bipolar cells, we used a high-divalent (Hi-Di) solution, which blocks voltage-gated calcium channels involved in the calcium influx from the outside of the cell into the cytoplasm [23]. The composite of the Hi-Di solution was 135 mM NaCl, 5.5 mM glucose, 10 mM HEPES, 15 mM NaHCO3, 5 mM MgCl2, 10 mM CaCl2, and 3.5 mM KCl with a pH of 7.3–7.4.
Treating rd1 retinas with 10 µM Th for 30 min increased RGC firing and enhanced local field potential amplitudes. A subsequent 30−min ACSF perfusion reversed both to basal level. Applying Hi-Di solution for 30 min, blocking voltage-gated calcium channels, almost completely suppressed RGC firing and local field potentials. Post-treatment recovery with a 30−min perfusion of ACSF solution confirmed the safety of the chosen drug concentrations and treatment durations. These results validate the suitability of the selected conditions.
Data analysis
To determine the effect of burst stimulation on WT and RD RGC responses, we measured the number of RGC spikes after stimulus onset. The temporal structure of the RGC response to electrical stimulation was investigated using a post-stimulus raster.
In general, when electrical stimulation is applied to the retina, there are three pathways that induce RGC responses [24].
(1) Directly-evoked RGC responses in which RGCs are directly activated by electrical stimulation, showing short-latency spikes within 10 ms
(2) Bipolar cell-mediated RGC responses with median-latency spikes within 50 ms via synaptic relay after bipolar cell activation
(3) Photoreceptor-mediated RGC responses with long-latency spikes within 100 ms via synaptic relay after photoreceptor activation
This study eliminated directly-evoked RGC spikes within 10 ms after stimulus onset because stimulus artifacts obscure short-latency RGC spikes [25]. In addition, following electrical stimulation, no photoreceptor-mediated RGC response was observed in rd1 mice of P56 due to complete photoreceptor loss. Thus, all RGCs observed in this study fired spikes within 50 ms after stimulation, indicating that electrically-evoked bipolar cell responses are transmitted to RGCs via synaptic relays [24]. We call this response a bipolar cell-mediated RGC response. RGC response strength was quantified by counting the number of spikes per one stimulus trial within 50 ms after stimulation. Normalized responses were obtained by normalizing the number of electrically-evoked RGC spikes to the range [0–1] using feature scaling based on min-max normalization.
All results, including the number of electrically-evoked RGC spikes, were averaged for the analyzed RGCs. Error bars indicate the standard error of the mean (SEM). A paired t-test was performed for statistical analysis between the two groups (*p < 0.05, **p < 0.01, ***p < 0.001). Kruskal–Wallis test was performed with Tukey–Kramer, Bonferroni, and Scheffe post-hoc tests to determine the statistical difference among three or more groups (*p < 0.05).
Go to :
RESULTS
Burst stimulation preserves the spatial resolution of the RGC population and elicits more RGC spikes
We compared RGC responses in the normal retina to two types of electrical stimulation: single-pulse stimulation with high amplitude versus burst stimulation with multiple low-amplitude pulses (Fig. 2). In this experiment, 234 WT RGCs from eight retinal patches were used for analysis.
Both types of stimulation elicited RGC responses closest to the stimulating electrode. Specifically, Fig. 2A shows the electrically-evoked responses of RGCs recorded at the closest distance of 200 µm from the stimulation electrode. For single-pulse stimulation, we observed an increase in the number of RGC spikes with increasing amplitude (The left and right panels of Fig. 2A). We also found an increase in the number of RGC spikes for stimulation with increasing pulse number per burst (The middle and right panels of Fig. 2A). However, there was a difference between the maximum number of spikes that single-pulse and burst stimulation could elicit. At the highest stimulus charge, 12.5 nC, single-pulse stimulation with an amplitude of 50 µA elicited more RGC spikes (*p < 0.05) than burst stimulation with five pulses (The right panel of Fig. 2A).
Single-pulse stimulation with high amplitude elicited RGC responses distant from the stimulation electrode, while burst stimulation with multiple low-amplitude pulses elicited no RGC responses. Fig. 2B shows the electrically-evoked responses of RGCs recorded at 600 µm from the stimulation electrode. For single-pulse stimulation, we found that the number of RGC spikes increased with increasing amplitude (The left and right panels of Fig. 2B), resulting in lower spatial resolution. No electrically-evoked response was observed for burst stimulation as the number of pulses per burst increased (The middle and right panels of Fig. 2B).
We observed that single-pulse stimulation destroyed the spatial resolution of RGC populations, while burst stimulation preserved it. Fig. 2C compared the spatial changes in electrically-evoked RGCs between single-pulse and burst stimulation. For single-pulse stimulation, RGC responses showed a relatively gradual decreasing pattern (The left panel of Fig. 2C). In contrast, for burst stimulation, RGCs showed a sharp decrease in response strength as they moved away from the stimulation electrode (The right panel of Fig. 2C). This suggests that only RGCs closest to the stimulation electrode responded locally. Even when considering spatial MEA color maps of normalized RGC responses in a single retinal patch, burst stimulation with five pulses per burst resulted in more spatially localized RGC responses than single-pulse stimulation with 50 µA amplitude. To quantify the spatial distribution of RGC populations responding to electrical stimulation, we used 0.5 from the normalized RGC response as the margin distance for significant RGC activation. For single-pulse stimulation, the distance of RGCs was approximately 464 µm at 50 µA, whereas for burst stimulation, the distance of RGCs was narrower (~302 µm at five pulse counts).
The finding that burst stimulation increases RGC firing while maintaining spatial resolution was also consistently observed in the degenerate retina. In the degenerate retina experiment, 258 RD RGCs from eight retinal patches were used for analysis. For single-pulse stimulation, when the amplitude was large, RD RGCs showed electrically-evoked responses at both 200 µm and 600 µm distances (Fig. 3A, B). However, for burst stimulation, when a large number of pulses were applied to the retina, only RGCs 200 µm away, close to the stimulation electrode, showed electrically-evoked responses. Quantifying the spatial distribution of RGC populations responding to electrical stimulation, we observed that RGCs were more widely distributed (~846 µm) at 50 µA for single-pulse stimulation. In contrast, for burst stimulation, they were much more narrowly distributed (~351 µm) at five pulse counts (Fig. 3C).
Overall, single-pulse stimulation resulted in spatially extended RGC responses, whereas burst stimulation resulted in spatially confined RGC responses. Notably, RD RGCs showed a wider spatial distribution of electrically-evoked responses to single-pulse stimulation compared to WT retinas (The left panels of Fig. 2C vs. Fig. 3C). On the other hand, burst stimulation showed a similar spatial distribution between WT and RD RGCs, suggesting that burst stimulation is an advantageous stimulation method when applied to the degenerate retina of prosthesis-implanted patients.
Doublet or triplet pulse stimulation is sufficient to induce RGC spikes
Burst stimulation increased RGC firing, but not as much as a single pulse of the highest amplitude (50 µA) (Fig. 2A, 3A). Therefore, unlike previous experiments that limited the number of pulses to five during burst stimulation, we increased the number of pulses per burst to 20 to investigate how RGC responses would change. In this experiment, we chose only RGCs at a distance of 200 µm from the stimulation electrode that showed the largest response to burst stimulation, and 39 WT RGCs and 35 RD RGCs from five retinal patches were used in the analysis. As a result, we found an increase in the number of RGC spikes when stimulated with a doublet or triplet pulse compared to a single pulse in both WT and RD retinas (Fig. 4, 5). However, no further significant changes were observed when using more than four pulses per burst, suggesting that an unnecessarily large number of pulses are not required to induce RGC spikes, and that doublet or triplet pulse stimulation is sufficient.
RD RGCs fire similarly to WT RGCs with burst stimulation
We compared the firing level with burst stimulation in WT and RD RGCs. We analyzed only RGCs at a distance of 200 µm from the stimulation electrode, and we applied doublet and triplet stimulation to WT and RD retinas, as determined in Fig. 4 and 5. We used 47 WT RGCs and 41 RD RGCs from six retinal patches. When single pulses of 30 µA and 50 µA were applied to the retina, the spike number of WT RGCs was higher than those of RD RGCs (WT single pulse vs. RD single pulse in Fig. 6). However, when burst stimulation was applied to the RD retina, the number of electrically-evoked RGC spikes became similar to that of WT RGCs with a single pulse (WT single pulse vs. RD burst in Fig. 6).
Burst stimulation activates RGCs by enhancing calcium-mediated synaptic transmission compared to single-pulse stimulation
We investigated the physiological mechanisms by which burst stimulation increases RGC firing compared to single-pulse stimulation in the RD retina. Since the electrically-evoked RGC spikes observed in this study were bipolar cell-mediated responses occurring within 50 ms after stimulation, we hypothesized that burst stimulation increases neurotransmitter release from the presynaptic terminals of bipolar cells to RGCs. To test this hypothesis, we performed experiments to increase or decrease the intracellular calcium concentration, which plays an essential role in releasing glutamate from synaptic vesicles in bipolar cells. To increase the calcium concentration in the presynaptic terminals, we administered Th (10 µM), and to decrease the calcium concentration, we perfused the retina with a Hi-Di solution instead of normal ACSF. We used three retinal patches from RD mice and 21 RGCs recorded at 200 µm from the stimulation electrode, for the pharmacological experiments.
First, Fig. 7A, B shows the representative spontaneous activity of RD RGCs recorded before and after treatment with Th and the Hi-Di solution. Before treatment, RD RGCs showed hyperactive spontaneous firing and abnormal oscillation of local field potentials. After Th treatment, higher spontaneous firing and more periodic oscillations were observed compared to without Th. On the other hand, after the Hi-Di solution treatment, the oscillations completely disappeared, and only a few spikes were observed.
 | Fig. 7Calcium-mediated RGC responses to burst stimulation in the RD retina.(A) Representative 1-second raw traces of RGCs in the RD retina without and with thapsigargin (Th) or Hi-Di solution. (B) Spontaneous firing rates of RGCs for RD, RD + Th, and RD + Hi-Di. Statistical differences in the graphs are indicated by asterisks (***p < 0.001). (C) Typical responses of RD RGCs to burst stimulation are shown as raster plots. Electrically-evoked RGC spikes are highlighted. The time window of the raster plot is set to ± 0.1 sec, with 0 sec representing the stimulus onset (red dashed line). In each raster plot (10 trials), the pulse number increases from top to bottom (#1–#5). The pulse amplitude and pulse duration are fixed with 30 μA and 100 μs/phase. Only RGCs 200 μm closest to the stimulation electrode are analyzed. (D) Normalized RGC response curves to burst stimulation. Error bars represent the standard error of the mean (SEM). Statistical differences in the response curves are indicated by asterisks (*p < 0.05, **p < 0.01, ***p < 0.001). RGC, retinal ganglion cell; RD, retinal degenerate; Hi-Di, high-divalent.
|
Fig. 7C, D shows the responses of RD RGCs to burst stimulation. The pre-treatment RGC responses showed a significant increase in the number of RGC spikes by burst stimulation compared to a single pulse (the normalized response of triplet pulse compared to a single pulse was 0.12 higher; **p < 0.01). The number of RGC spikes was increased by burst stimulation after Th treatment (0.22 higher normalized response of triplet pulse compared to a single pulse; ***p < 0.001). However, the increase became smaller after Hi-Di solution treatment (0.04 higher normalized response of triplet pulse compared to a single pulse; *p < 0.05). These results suggest that the electrically-evoked RGC firing observed in the degenerate retina is associated with calcium-mediated synaptic transmission and that burst stimulation appears to enhance this network.
Go to :
DISCUSSION
Fig. 8 illustrates a schematic diagram to explain the burst stimulation-mediated response. When a single-pulse electrical stimulation is applied to the degenerate retina, the membrane potential of the bipolar cell depolarizes, which in turn opens the voltage-gated Ca2+ channels in the cell membrane. However, this causes only a minimal influx of calcium ions into the cell, releasing a small amount of glutamate from a limited number of synaptic vesicles. As a result, the membrane potential of the RGC remains below the firing threshold, resulting in low firing rates. On the other hand, burst stimulation with multiple electrical pulses activates the voltage-gated Ca2+ channels of bipolar cells, leading to a significant influx of calcium ions into the cell. This large calcium influx causes the release of a substantial amount of glutamate from numerous synaptic vesicles, which in turn increases the firing rates of RGCs. Additionally, Th, a blocker of the SERCA, induces an increase in intracellular calcium concentration, further amplifies glutamate release and contributes to the increased firing rates of RGCs.
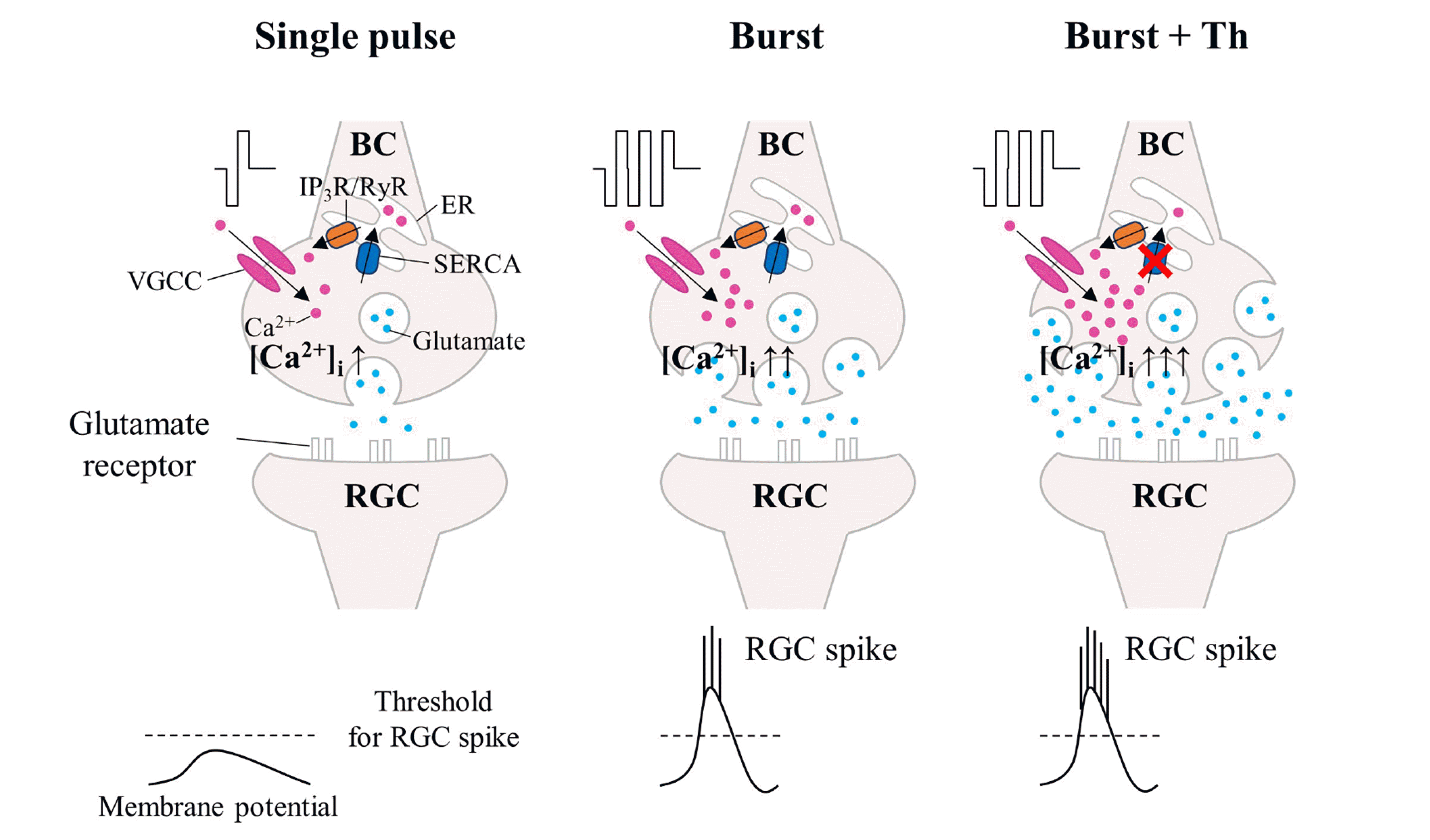 | Fig. 8Schematic diagram of calcium-mediated RGC firing to the burst stimulation.The burst stimulation strategy depends on an increase of intracellular calcium concentration, regulated by voltage-gated calcium channels (VGCCs) for calcium influx and sarco/endoplasmic reticulum Ca2+–ATPase (SERCA) for calcium reuptake. Burst stimulation activates VGCCs at bipolar cell (BC) synapses, boosting calcium influx and then glutamate release from the BC, therefore increasing RGC firing. Thapsigargin (Th) inhibits SERCA, preventing calcium reuptake, raising intracellular calcium concentration ([Ca2+]i), and then releasing glutamate from the bipolar cell, thus enhancing RGC firing. RGC, retinal ganglion cell; IP3R/RyR, inositol–1,4,5–triphosphate receptors/ryanodine receptors; ER, endoplasmic reticulum.
|
In summary, our results demonstrate that burst stimulation can effectively increase RGC firing while preserving the spatial resolution of RGC populations. We identified that doublet or triplet pulses per burst were optimal for eliciting RGC spikes, and burst stimulation induced similar enhanced spike rates in both WT and RD RGCs. Additionally, pharmacological studies suggest that RGC firing via burst stimulation is closely related to the enhanced calcium-mediated synaptic transmission in bipolar cells.
Burst stimulation has been widely used as a stimulation method for pain reduction in spinal cord stimulation research and was approved by the FDA in 2016 [26]. Several studies have reported that burst stimulation is more effective in reducing pain than single-pulse stimulation. While the burst stimulation method is well-established in other research fields, it is newly adopted for retinal stimulation in this study. While single-pulse stimulation remains the traditional protocol in clinical studies of retinal prostheses, it faces an inevitable trade-off between increasing spatial resolution and RGC firing [5,6]. Therefore, we adopted a pulsing strategy of minimizing pulse amplitude to maintain spatial resolution while using multiple consecutive pulses in a burst to increase RGC firing (Fig. 2, 3).
Previous studies have demonstrated that RD RGCs exhibit lower firing than WT RGCs upon the retina's electrical stimulation [7,8]. Consequently, determining optimal stimulation conditions to enhance RD RGC responses to a level similar to that of WT RGCs is a challenging task in the community of retinal prostheses. A straightforward approach is to apply stronger electrical stimulation to the RD retina; however, as previously mentioned, single-pulse stimulation with high amplitude significantly loses spatial resolution (Fig. 2, 3). Therefore, we propose a burst stimulation approach to increase RGC firing while maintaining spatial resolution (Fig. 2, 3, 6).
Burst stimulation offers a notable advantage over single long-duration pulse stimulation by achieving safe and effective retinal stimulation. Weitz et al.’s [27] have demonstrated the enhanced spatial resolution of direct RGC responses with long-duration pulses. Still, longer durations inevitably elevate the stimulation threshold charge density needed for RGC firing (Fig. 1 of [27]). Besides, the risk of exceeding safety limits and damaging retinal tissue becomes more pronounced with reduced electrode diameters from 200 µm to 30 µm (Fig. 3 of [27]). For example, applying a 25 ms pulse duration to WT retinas with a 30 µm electrode diameter resulted in a threshold charge density above the recommended limit of 1 mC/cm2 [27]. Several studies recommend a threshold charge density under 1 mC/cm2 to keep the retinal tissue intact [7,28-30]. The single long-duration pulse stimulation may be advantageous for the spatial resolution of direct RGC responses. Still, at the same time, it inevitably elevates the risk of damage to retinal tissue. The comparison of the effect of single long-duration pulse vs. burst stimulation is needed for future studies focusing on network-mediated RGC responses.
Previous studies consistently favor short-duration, high-amplitude pulses over long-duration, low-amplitude ones for eliciting RGC spikes at the same charge condition [7,31,32]. Expanding on this insight, our study strategically employs consecutive short-duration (100 µs), low amplitude (30 µA) biphasic pulses to achieve a safe charge density of 0.43 mC/cm2. This approach effectively enhances RGC firing and spatial resolution of network-mediated RGC responses (Fig. 2, 3, 6).
Burst stimulation offers a significant advantage over high-frequency stimulation by preventing RGC desensitization, a decreased firing due to repeated stimuli. High-frequency stimulation, which continuously uses the same single pulses, can result in RGC desensitization [33-35]. In contrast, burst stimulation transiently applies multiple pulses, minimizing the risk of RGC desensitization (Fig. 2–5). Compared to high-frequency and single long-duration pulse stimulation, burst stimulation is an adaptable approach that enhances RGC firing and spatial resolution of network-mediated responses while avoiding desensitization and safety concerns.
The burst stimulation strategy has the advantage of increasing RGC firing while preserving high-resolution retinal responses compared to single-pulse stimulation. However, more work is needed to improve the performance of burst stimulation. In this study, although burst stimulation increased RGC firing, the number of spikes was significantly lower than single-pulse stimulation with the highest amplitude of 50 µA (The right panels of Fig. 2A, 3A). Therefore, it is essential to optimize the burst stimulation protocol. The weak performance of burst stimulation on RGC firing may be because the pulses used in this study were biphasic. Because the immediately following anodic pulse canceled out the charge of the preceding cathodic pulse, the cumulative effect of the injected charge was lost as the number of pulses increased. In future studies, the burst stimulation pattern can be modified using monophasic pulses instead of biphasic pulses to maximize the charge accumulation effect, leading to more RGC firing. For instance, the stimulation pattern can be altered by applying multiple cathodic pulses first, followed immediately by multiple anodic pulses. However, it is essential to consider safety issues, such as tissue damage and electrode erosion, since perfect charge balancing is difficult to achieve with monophasic pulses [29].
The burst stimulation strategy offers a potential solution to the problem of low RGC firing caused by local return electrodes, which is a current challenge in artificial retinal research. Return electrodes have been placed near the stimulation electrode to prevent current spreading, increasing spatial resolution [36]. However, this approach has significantly decreased the number of electrically-evoked RGC spikes due to excessive electrical field limitation by local return electrodes [37,38]. Increasing pulse amplitude with conventional single-pulse stimulation to achieve higher RGC firing decreases spatial resolution in a vicious cycle. Burst stimulation can be an effective alternative to single-pulse stimulation for addressing this issue. For example, applying doublet or triplet pulse stimulation to the retina (Fig. 4–6) with grounding around the stimulation electrode can achieve high RGC firing and spatial resolution. Future studies combining burst stimulation with local return electrodes are needed.
Burst stimulation faces a limitation on pulse counts. Pulse counts must be restricted to avoid masking network-mediated RGC responses. Burst stimulation increases pulse counts, extending total stimulation time, and resulting in a higher charge than a single biphasic pulse. As shown in Fig. 1C, increasing 100 µs duration biphasic pulses elongates stimulation time: Single-biphasic pulse: 200 µs, Two, three, ten, twenty consecutive biphasic pulses: 400 µs, 600 µs, 2000 µs (2 ms), 4,000 µs (4 ms), respectively. Since this study focused on network-mediated RGC responses, typically observed around 20 ms post-stimulation, using 100 pulses could lead to a 20 ms total stimulation time, obscuring network-mediated RGC responses with stimulus pulses.
Fortunately, using more than 7 pulses (30 µA × 100 µs × 7 = 21 nC) is practically unnecessary due to safety limits (20 nC or less is recommended to avoid retinal tissue damage.) [7]. Fig. 4 and 5 also emphasize no significant RGC firing increment above triplet pulse stimulation, implying no practical usefulness of quadruplets and more pulses.
While this study successfully demonstrated the efficacy of burst stimulation in normal and degenerate mouse models, its feasibility for human clinical studies necessitates validation in a non-human primate model that closely resembles a human retinal system. Our previous work established an RD monkey model [39], highlighting the drawbacks of single high-amplitude pulse stimulation on RGC response resolution [6]. Our upcoming research will assess the efficacy of burst stimulation to enhance spatial resolution in the RD monkey model, potentially enabling translation into human clinical studies based on insights from the non-human primate model.
In conclusion, this study demonstrates that burst stimulation is a promising approach to induce focal responses in RD RGCs and increase their firing to levels similar to WT RGCs. The performance of burst stimulation can be optimized by modifying the stimulation pulse, such as using monophasic pulses. The application of burst stimulation in retinal prostheses can be beneficial. Therefore, the newly adapted burst stimulation strategy can significantly improve low vision in RP patients implanted with retinal prostheses.
Go to :
 XML Download
XML Download