

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Infrared (IR) radiation is classified into three wavelength bands: IR-A (0.78–1.4 µm), IR-B (1.4–3 µm), and IR-C (3–1,000 µm) as defined by the International Commission on Illumination.1 Among them, IR-C is classified as a far-infrared (FIR) ray that conveys electromagnetic energy (photons with quantum energy levels of 12.4 meV–1.7 eV).1 When living cells are irradiated with FIR, its energy is absorbed and alters the vibrational status of bonds in various molecules, including water, by changing six vibrational modes, such as symmetric and antisymmetric stretching, scissoring, rocking, wagging, and twisting, which can have diverse biological effects including the inhibition of cell proliferation.12
Several studies reported that FIR irradiation has beneficial effects on cardiovascular diseases, including congestive heart failure, hypertension, and atherosclerosis.34 For example, repeated FIR therapy has been reported to prevent atherosclerosis by reducing oxidative stress in patients with coronary risk factors.5 Furthermore, FIR irradiation improved arteriovenous fistula access flow and patency in hemodialysis patients.6 Although many vasoprotective effects of FIR irradiation have been reported, the detailed molecular mechanisms are unclear.
Vascular smooth muscle cells (VSMCs) are stromal cells located in the medial layer of the blood vessel wall and respond to vasoactive stimuli, such as hormones, metabolites, and neurotransmitters through which VSMCs play an important role in the maintenance of vascular integrity and homeostasis through regulation of blood pressure and distribution.789 In normal blood vessels, VSMCs exist mainly as a contractile phenotype and show a low phenotype turnover. On the other hand, when stimulated by pathological injuries such as inflammation, hypertension, and atherosclerosis conditions, VSMCs are activated and undergo phenotype switching from a contractile phenotype to a synthetic one.101112 Of various growth factors involved in the VSMC phenotype switching, the platelet-derived growth factor (PDGF) is an important growth factor that stimulates synthetic phenotype switching of VSMCs, leading to enhanced VSMC migration and proliferation in the development of atherosclerosis.111314 Clinically, it was reported that restenosis occurs within the first six months in 30–40% of patients who have previously undergone percutaneous balloon angioplasty.15 Furthermore, the aberrant proliferation of VSMCs has been considered critical in developing in-stent restenosis, transplant vasculopathy, and vein bypass graft failure.161718 Therefore, the functional normalization of dysregulated VSMC proliferation might be useful in preventing and treating atherosclerosis and post-intervention restenosis.
AMP-activated protein kinase (AMPK) is pivotal in regulating cellular energy metabolism, including glucose and lipid metabolism, and cell proliferation and autophagy in response to metabolic stress.1920 Of the many downstream targets of AMPK, the mammalian target of rapamycin (mTOR)/p70 S6 kinase (p70S6K) signaling pathway responsible for the regulation of protein synthesis, cell proliferation, and metabolism1921 is blocked when AMPK is activated, leading to the inhibition of cell proliferation in various types of cells.22232425 A recent study reported that activation of AMPK by telmisartan, an angiotensin II type 1 receptor blocker, decreases basal and PDGF-stimulated VSMC proliferation via inhibiting the mTOR/p70S6K signaling pathway.25 Nevertheless, no study has been reported on whether FIR irradiation inhibits VSMC proliferation and the AMPK/mTOR/p70S6K signaling axis.
Although excessive VSMC proliferation contributes to the development of various vascular diseases, including atherosclerosis and in-stent restenosis, the effects of FIR irradiation on VSMC proliferation and its mechanism of action remain elusive. This study investigated the molecular mechanism through which FIR irradiation inhibits rat VSMC proliferation.
METHODS
Materials
Dimethyl sulfoxide (DMSO) was obtained from Sigma-Aldrich (St. Louis, MO, USA). Three-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) were purchased from Cayman Chemicals (Ann Arbor, MI, USA). Compound C and recombinant rat PDGF-BB protein were purchased from Calbiochem (Darmstadt, Germany) and Novus Biologicals (Centennial, CO, USA), respectively. Antibodies against p-mTOR-Ser2448, mTOR, p-p70S6K-Thr389, p70S6K, p-AMPK-Thr172, and AMPK were obtained from Cell Signaling Technology (Beverly, MA, USA). Antibody against glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was acquired from AbFrontier (Seoul, Korea). Dulbecco’s modified Eagle’s medium (DMEM) was obtained from Fisher Scientific (Ottawa, Canada). Dulbecco’s phosphate-buffered saline (DPBS), fetal bovine serum (FBS), penicillin and streptomycin antibiotics, trypsin–EDTA solution, and plasticware for cell culture were supplied by Gibco-BRL (Gaithersburg, MD, USA). All other chemicals used were of the purest analytical grade available.
Cell culture and drug treatments
Rat aortic VSMCs were isolated and cultured as previously described.25 Briefly, six-week-old male Sprague-Dawley (SD) rats were euthanized with CO2 gas and subsequent cervical dislocation, and their thoracic aortas were immediately dissected. Connective tissues were removed, and the endothelium was denuded by gentle rubbing. The endothelium-deprived aortas were cut into 1-mm pieces and grown in DMEM supplemented with 10% FBS at 37°C under a 5% CO2 humidified air. Cells between passages 3 and 7 were used for the experiments. VSMCs grown to 90% confluence in 60-mm culture dishes were incubated in the absence or presence of FIR radiation in DMEM supplemented with 2% FBS. In some experiments, cells were co-treated with the indicated drugs or chemicals for the indicated times.
FIR irradiation
Rat VSMCs or isolated rat aortas were irradiated with FIR using a ceramic infrared radiator (model No. IOT/90-250; Elstein-Werk M. Steinmetz GmbH & Co. KG, Northeim, Germany), as performed previously in the authors’ laboratory.2627 The emission wavelength of the FIR radiator ranged from 1 to 20 µm with a 4 µm peak wavelength (Supplementary Fig. 1), and the irradiance at the surface of the FIR radiator and the place of a 60-mm culture dish was 2,530 mW/cm2 and 65 mW/cm2, respectively. Using the FIR generator, VSMCs or aortas cultured in a 60-mm culture dish with 3 mL medium were exposed to FIR ray at room temperature (RT) for the indicated times (0, 15, 30, or 45 minutes). For hyperthermal control experiments, VSMCs in a 60-mm culture dish were placed on the heat block set at 39°C for 30 minutes.
Western blot analyses
VSMCs irradiated with FIR radiation in the absence or presence of various chemicals were lysed in lysis buffer (20 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% Triton X-100, 1 mM EDTA, 1 mM EGTA, 1 mM PMSF, 10 mM β-glycerophosphate, 1 mM NaF, 1 mM Na3VO4, and 1× protease inhibitor cocktail from Roche Molecular Biochemicals [Indianapolis, IN, USA]). In addition to VSMCs, the endothelium-deprived rat aortas were either irradiated with FIR radiation or not. The proteins were extracted by chopping the aortic tissues using iris scissors in an ice-cold lysis buffer as previously described.28 Protein concentration was determined using a BCA protein assay kit from Thermo Scientific (Rockford, IL, USA). Equal quantities of protein (20 µg) were separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a nitrocellulose membrane from GE Healthcare Life Sciences (Pittsburgh, PA, USA). The blots were probed with the appropriate primary antibodies, followed by the corresponding secondary antibodies, all from Invitrogen (Carlsbad, CA, USA), and developed using ECL reagents from Amersham Biosciences (Arlington Heights, IL, USA). The primary antibody dilutions used for western blot analyses were as follows: p-mTOR-Ser2448 (1:1,000), mTOR (1:1,000), p-p70S6K-Thr389 (1:1,000), p70S6K (1:1,000), p-AMPK-Thr389 (1:1,000), AMPK (1:1,000), and GAPDH (1:3,000).
Transfection of dominant negative (dn)-AMPKα1 constructs
The dn-AMPKα1 cDNA construct was a kind gift from Prof. Joohun Ha (Kyung Hee University, Seoul, Korea). Transfection of pcDNA3.1 vector containing rat dn-AMPKα1 cDNA carrying a point mutation of GAC → GCC at the 157th codon (D157A mutant) was performed as described elsewhere.2529 Briefly, the dn-AMPKα1 cDNA construct was transfected to VSMCs grown to 70% confluence in 60-mm or 35-mm culture dishes, using Solfect reagent from Biosolyx (Daegu, Korea), according to the manufacturer’s instructions. Equal amounts of pcDNA3.1 vector were transfected for control. After 5 hours incubation at 37°C, the cells were further incubated in DMEM containing 10% FBS for 24 hours before FIR irradiation.
Measurement of cell proliferation
Viable cells were measured using an MTT assay. VSMCs were seeded into 35-mm culture dishes in DMEM supplemented with 10% FBS at 37°C for 24 hours. When the cells reached 60% confluence, the medium was replaced with 2% FBS DMEM, followed by FIR irradiation for the indicated times (0, 15, 30, or 45 minutes), and the cells were allowed to proliferate for 24 hours. In a separate experiment, cells were irradiated with FIR ray for 30 min, allowed to proliferate for 24 hours or treated with 1 mM AICAR for 24 hours, and then an MTT solution (5 mg/mL) was added to each 35-mm culture dish (1:10) and further incubated for 2 hours. The supernatants were aspirated, and the dishes were washed once with DPBS. Formazan was then dissolved in 1 mL DMSO. An aliquot (200 µL) of this solution was transferred to 96-well plates. Cell proliferation was assessed by measuring the absorbance at 570 nm using a microplate reader (Microplate reader model 680; Bio-Rad Laboratory, Irvine, CA, USA).
Cell death assay
The VSMC death was assessed using a LIVE/DEAD Kit (Thermo Fisher Scientific, Waltham, MA, USA). Briefly, after the cells were incubated on coverslips in 35-mm culture dishes in the absence or presence of FIR irradiation for the indicated times (0, 15, 30, or 45 minutes), LIVE/DEAD assay reagents (0.1 µM calcein-acetoxymethyl ester and 16 µM ethidium homodimer-1) were added in the culture medium for 20 minutes at RT. The cells were fixed in 4% (wt/vol) paraformaldehyde and washed three times in DPBS. Images of the labeled cells were acquired at 200× magnification using fluorescent microscopy (Olympus, Tokyo, Japan), and live or dead cells were counted. Cell death was expressed as % of dead cells relative to total cells.
Detection of intracellular Ca2+ localization
Intracellular Ca2+ localization was detected by Fura-2 AM (Invitrogen), an intracellular Ca2+ indicator, according to the manufacturer’s protocol. Briefly, VSMCs grown on coverslips were either irradiated with FIR for 30 minutes or not, treated with 2 µM Fura-2 AM (Invitrogen), and then fixed with 4% (wt/vol) paraformaldehyde for 20 min. The nuclei were also detected using 1 µM Hoechst33342 (Tocris Bioscience, Ellisville, MO, USA). Images of intracellular Ca2+ localization were photographed using a confocal microscope (K1-Fluo; Nanoscope Systems Inc., Daejeon, Korea).
Measurement of cellular adenosine 5′-triphosphate (ATP) levels
The cellular ATP levels were measured using a luminescent ATP detection assay kit (Cayman Chemicals) according to the manufacturer’s protocol. Briefly, VSMCs were seeded into 35-mm culture dishes. When the cells reached up to 90% confluence, the medium was replaced with 2% FBS DMEM, followed by FIR irradiation for 30 minutes. The cells were washed once with ice-cold DPBS, and the cell lysates were obtained using a 1× ATP assay sample buffer (40 µL). The supernatants were collected by centrifugation, and ATP reaction buffer (110 µL) containing D-luciferin and luciferase was added to 10 µL of each cell supernatant. The luminescence was then measured using a microplate reader (Molecular Devices, Sunnyvale, CA, USA). The final cellular ATP levels were obtained by normalizing the luminescence to the total protein contents.
Animals
All animal experiments were conducted in accordance with the approved institutional guidelines for animal care and use at Yeungnam University (approval No. YUMC-AEC2021-020). Six-week-old male SD rats were maintained for one week at the beginning of the experiment in a temperature- and humidity-controlled room (22 ± 1°C and 50 ± 10%, respectively) under a 12-hour alternate light/ dark cycle. All rats were given water and fed a standard chow from Purina Mills, LLC (St. Louis, MO, USA) ad libitum throughout the experiments.
RESULTS
Cell proliferation is decreased in FIR-irradiated rat VSMCs
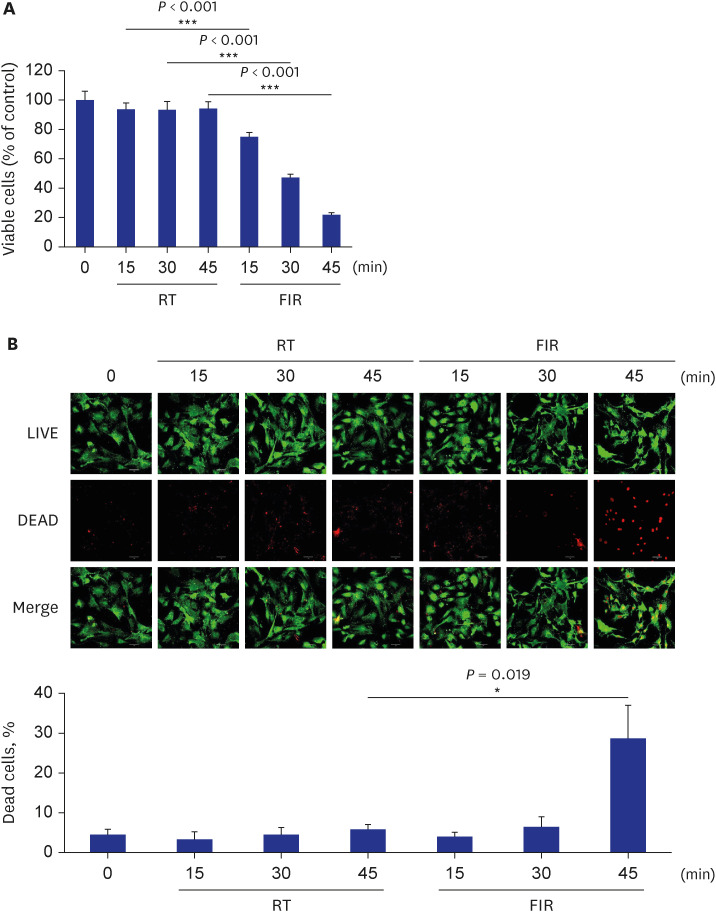
Although aberrant VSMC proliferation in vessels contributes to the development of many cardiovascular diseases, such as atherosclerosis and restenosis,30 the effect of FIR irradiation on VSMC proliferation is unclear. Therefore, this study examined whether FIR irradiation decreases cell proliferation using rat VSMCs. As shown in Fig. 1A, FIR irradiation inhibited VSMC proliferation in an FIR-irradiation-time-dependent manner. When the cells were irradiated with FIR ray for 30 minutes, cell viability decreased to 50% compared to RT control (Fig. 1A). Interestingly, viable cells were reduced to 20% of RT control when cells were exposed to FIR radiation for 45 minutes (Fig. 1A), suggesting that prolonged FIR irradiation (45 minutes) may not only inhibit cell proliferation but also induce cell death. These results prompted us to examine whether FIR irradiation causes cell damage in VSMCs at all experimental points. To this end, a LIVE/DEAD assay was performed. As shown in Fig. 1B, FIR irradiation for 30 minutes did not induce VSMC death with 5% of basal cell death, whereas prolonged FIR irradiation (45 minutes) elicited cell death up to 30% of total cells. Since no significant cell death was observed in VSMCs exposed to FIR radiation for 30 minutes (Fig. 1B), all subsequent experiments were carried out under this condition, unless mentioned otherwise.
Fig. 1
FIR irradiation decreases VSMC proliferation. (A) Rat VSMCs were exposed to FIR radiation for various times (0, 15, 30, or 45 minutes), and VSMC proliferation was measured by MTT assay as described in the Methods. (B) Rat VSMCs were exposed to FIR radiation for various times (0, 15, 30, or 45 minutes), and live or dead cells were measured using a LIVE/DEAD assay kit as described in the Methods. Scale bar indicates 50 µm. All experiments were independently performed at least four times, and the fluorescent images shown are representative of at least four experiments (n = 4). The bar graphs depict the mean percent alterations above/below the control levels (± standard deviation).
FIR = Far-infrared, VSMC = vascular smooth muscle cell, MTT = 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, RT = room temperature.
Differences were considered statistically significant at *P < 0.01 and ***P < 0.001.
FIR irradiation decreases mTOR/p70S6K phosphorylation, and cellular ATP levels that increase AMPK phosphorylation in VSMCs and isolated aortas
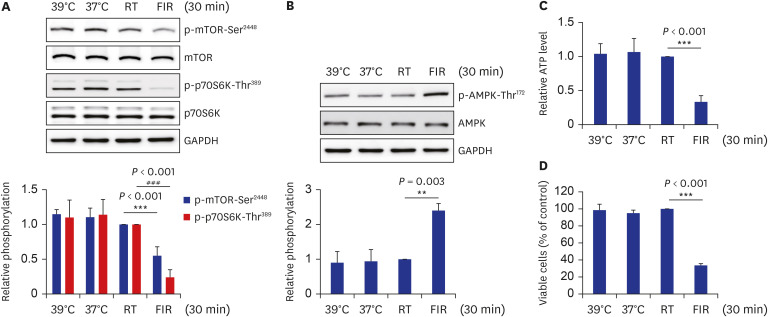
AMPK activation by AICAR was reported to inhibit the mTOR/p70S6K signaling pathway and, consequently, cell proliferation in several multiple myeloma cell lines.31 Furthermore, a recent study reported that telmisartan-activated AMPK mitigates VSMC proliferation via inhibiting p-mTOR-Ser2448 and p-p70S6K-Thr389.25 Therefore, this study examined whether the AMPK/mTOR/p70S6K signaling axis changed after irradiating FIR ray to VSMCs. FIR irradiation decreased p-mTOR-Ser2448 and p-p70S6K-Thr389 and elevated p-AMPK-Thr172 in an FIR irradiation time-dependent manner with no change in the total protein expression (Fig. 2A and B). The cellular ATP levels were measured to clarify the underlying mechanism through which AMPK was activated in FIR-irradiated cells. The ATP levels in cells exposed to FIR radiation for 30 minutes were reduced to 30% of RT control (Fig. 2C). Furthermore, the inhibitory effects of FIR irradiation for 30 minutes on VSMC proliferation and AMPK/mTOR/p70S6K signaling axis were greater than those of 1 mM AICAR treatment (Supplementary Fig. 2). Level of p-AMPK-Thr172 was increased 2-fold in FIR-irradiated cells compared to that in 1 mM AICAR-treated cells (Supplementary Fig. 2A), and levels of p-mTOR-Ser2448 and p-p70S6K-Thr389 in FIR-irradiated cells were decreased to 50% of those in 1 mM AICAR-treated cells (Supplementary Fig. 2B). In accordance with these results, cell proliferation in FIR-irradiated cells was observed to be 60% compared to that in 1 mM AICAR-treated cells (Supplementary Fig. 2B). Next, we attempted to determine if these in vitro findings are recapitulated in isolated rat aortas. In line with in vitro results, FIR irradiation for 30 min increased p-AMPK-Thr172 and decreased p-mTOR-Ser2448 and p-p70S6K-Thr389 in endothelium-deprived rat aortas (Fig. 2D and E). These results suggest the possibility that FIR irradiation decreases VSMC proliferation via the AMPK-mediated inhibition of the mTOR/p70S6K signaling pathway, which is possibly mediated by similar mechanisms in vitro, ex vivo, and perhaps in vivo, and its relevance to physiological function.
Fig. 2
FIR irradiation decreases mTOR/p70S6K phosphorylation and cellular ATP levels, increasing AMPK phosphorylation in VSMCs and isolated aortas. (A, B) Rat VSMCs were exposed to FIR radiation for various times (0, 15, or 30 minutes), and levels of p-mTOR-Ser2448, p-p70S6K-Thr389, and p-AMPK-Thr172 were evaluated by western blotting. (C) Rat VSMCs were irradiated with FIR ray or not for 30 minutes, cellular ATP levels were assessed using a luminescent ATP detection assay kit, as described in the Methods. (D, E) Endothelium-deprived rat aortas were irradiated with FIR ray or not for 30 minutes, and aortic proteins were extracted and subjected to western blot analyses. The levels of p-mTOR-Ser2448, p-p70S6K-Thr389, and p-AMPK-Thr172 were detected by western blotting. All experiments were independently performed at least four times, and the blots shown are representative of at least four experiments (n = 4). The bar graphs depict mean fold alterations above/below the controls (± standard deviation).
FIR = Far-infrared, mTOR = mammalian target of rapamycin, p70S6K = p70 S6 kinase, ATP = adenosine 5′-triphosphate, AMPK = AMP-activated protein kinase, VSMC = vascular smooth muscle cell, RT = room temperature.
Differences were considered statistically significant at *P < 0.05, #P < 0.05, ***P < 0.001, and ###P < 0.001.

AMPK-inhibited mTOR/p70S6K signaling pathway mediates FIR irradiation-decreased VSMC proliferation
Inhibitor studies were performed using compound C, a specific AMPK inhibitor, to reveal the role of AMPK in FIR irradiation-inhibited p-mTOR-Ser2448 and p-p70S6K-Thr389. As shown in Fig. 3A, pre-treatment with 10 µM compound C completely reversed FIR irradiation-reduced p-mTOR-Ser2448 and p-p70S6K-Thr389. dn-AMPKα1 (D157A mutant) constructs were introduced into VSMCs to confirm these results. As shown in Fig. 3B, overexpression of the dn-AMPKα1 gene was successful, as evidenced by the increased total AMPK expression and reduced p-AMPK-Thr172 in dn-AMPKα1 gene-transfected cells. Similar to the results for compound C, The FIR irradiation-induced decreases in p-mTOR-Ser2448 and p-p70S6K-Thr389 were restored in dn-AMPKα1-overexpressed VSMCs (Fig. 3C). Furthermore, ectopic expression of dn-AMPKα1 gene significantly reversed the FIR irradiation-repressed VSMC proliferation up to 30% (Fig. 3D). Hence, FIR irradiation-activated AMPK inhibits VSMC proliferation by downregulating the mTOR/p70S6K signaling pathway.
Fig. 3
FIR irradiation-activated AMPK inhibits VSMC proliferation by decreasing mTOR/p70S6K signaling pathway. (A) Rat VSMCs were exposed to FIR radiation for 30 minutes in the absence or presence of 10 µM compound C, and levels of p-mTOR-Ser2448 and p-p70S6K-Thr389 were detected by western blotting. (B, C) Cells were transfected with rat dn-AMPKα1 (D157A) gene or empty vector and exposed to FIR radiation for 30 minutes. Levels of p-AMPK-Thr172, p-mTOR-Ser2448, and p-p70S6K-Thr389 were assessed by western blotting. (D) Rat dn-AMPKα1 (D157A) gene- or empty vector-transfected VSMCs were exposed to FIR radiation for 30 minutes, and cell proliferation was measured using an MTT assay, as described in the Methods. All experiments were performed at least four times independently, and the blots shown are representative of at least four experiments (n = 4). Bar graphs depict mean fold alterations above/below the controls (± standard deviation).
FIR = Far-infrared, AMPK = AMP-activated protein kinase, VSMC = vascular smooth muscle cell, mTOR = mammalian target of rapamycin, p70S6K = p70 S6 kinase, dn = dominant negative, MTT = 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, RT = room temperature.
Differences were considered statistically significant at *P < 0.05, #P < 0.05, and **P < 0.01.

Inhibitory action of FIR irradiation on VSMC proliferation, cellular ATP levels, and AMPK/mTOR/p70S6K signaling axis is FIR’s peculiar effect, not the hyperthermal effect
Several studies showed that FIR irradiation increases the temperature of the culture medium to 38 ± 1°C.2732 This hyperthermal effect of FIR irradiation was suggested to mediate the biological effects of FIR irradiation, such as inhibition of tumor growth.3334 Based on these reports, this study explored whether the hyperthermal effect of FIR irradiation causes the observed results. The temperature of the culture medium was measured under the present experimental conditions. The temperature increased to 38–39°C at 15 minutes after FIR irradiation and maintained for 45 minutes (Supplementary Fig. 3). Therefore, the hyperthermal control was set to 39°C by placing culture dishes on the heat block for 30 minutes and conducted further experiments. Only FIR irradiation, not hyperthermal condition (39°C), enhanced p-AMPK-Thr172 and downregulated p-mTOR-S2448 and p-p70S6k-Thr389 (Fig. 4A and B). Furthermore, cellular ATP levels were reduced in only the FIR irradiation group, not the hyperthermal group (Fig. 4C). Decreased VSMC proliferation was observed in only FIR-irradiated cells (Fig. 4D). These results showed that the inhibitory actions of FIR irradiation on VSMC proliferation, cellular ATP levels, and AMPK/mTOR/p70S6K signaling axis were medicated by the peculiar effects of FIR ray, not hyperthermal effects.
Fig. 4
Inhibitory action of FIR irradiation on VSMC proliferation, cellular ATP levels, and AMPK/mTOR/p70S6K signaling axis is FIR ray’s peculiar effect, not the hyperthermal effect. (A, B) Rat VSMCs were incubated for 30 minutes at 39°C using the heat block, at 37°C in the culture incubator, or at 25°C (RT), or exposed to FIR radiation for 30 minutes at 25°C. Levels of p-mTOR-Ser2448, p-p70S6K-Thr389, and p-AMPK-Thr172 were detected by western blotting. (C) The cells were treated as described above, and cellular ATP levels were assessed using a luminescent ATP detection assay kit as described in the Methods. (D) Cells were treated as described above, and cell proliferation was measured using an MTT assay as described in the Methods. All experiments were independently performed at least four times, and blots shown are representative of at least four experiments (n = 4). Bar graphs depict the mean fold alterations above/below the controls (± standard deviation).
FIR = Far-infrared, VSMC = vascular smooth muscle cell, ATP = adenosine 5′-triphosphate, AMPK = AMP-activated protein kinase, mTOR = mammalian target of rapamycin, p70S6K = p70 S6 kinase, RT = room temperature, MTT = 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
Differences were considered statistically significant at **P < 0.01, ***P < 0.001, and ###P < 0.001.
FIR irradiation attenuates PDGF-stimulated VSMC proliferation by inhibiting AMPK/mTOR/p70S6K signaling axis
PDGF, a mitogen and chemoattractant for VSMCs, has been reported to play a major role in VSMC migration, proliferation, and plaque formation during atherosclerosis.35 Hence, treatment with PDGF in cells is commonly used to mimic atherosclerosis in vitro. Therefore, an in vitro atherosclerosis model was established by treating with 10 ng/mL PDGF for 24 hours, as done previously in the laboratory,36 and conducting further experiments. As shown in Fig. 5A, VSMC proliferation was increased 1.3-fold in PDGF-stimulated cells compared to that in basal VSMCs. Exposure to FIR radiation for 30 minutes significantly attenuated the PDGF-stimulated VSMC proliferation. In parallel with the results observed in basal condition (Fig. 3B and C), overexpression of the dn-AMPKα1 gene significantly alleviated FIR irradiation-induced p-AMPK-Thr172. It reversed p-mTOR-Ser2448 and p-p70S6K-Thr389 decreased by FIR irradiation (Fig. 5B and C). Furthermore, the ectopic expression of the dn-AMPKα1 construct restored FIR irradiation-inhibited VSMC proliferation significantly, as shown in Fig. 5D. These results clearly showed that FIR irradiation decreased VSMC proliferation in PDGF-induced in vitro atherosclerosis model as well as basal VSMCs through AMPK-mediated inhibition of mTOR/p70S6K signaling pathway (Fig. 6).
Fig. 5
FIR irradiation attenuates PDGF-stimulated VSMC proliferation by inhibiting AMPK/mTOR/p70S6K signaling axis. (A) Rat VSMCs were irradiated with FIR ray for 30 minutes, followed by incubation for 24 hours in the absence or presence of 10 ng/mL PDGF, and then cell proliferation was measured by MTT assay, as described in the Methods. (B, C) After rat VSMCs were transfected with rat dn-AMPKα1 (D157A) gene or empty vector, cells were irradiated with FIR ray for 30 minutes, followed by incubation for 24 hours in the absence or presence of 10 ng/mL PDGF. The levels of p-AMPK-Thr172, p-mTOR-Ser2448, and p-p70S6K-Thr389 were detected by western blotting. (D) Cells were treated as described above, and cell proliferation was assessed by MTT assay as described in the Methods. All experiments were independently performed at least four times, and the blots shown are representative of at least four experiments (n = 4). The bar graphs depict mean fold alterations above/below the controls (± standard deviation).
FIR = Far-infrared, PDGF = platelet-derived growth factor, VSMC = vascular smooth muscle cell, AMPK = AMP-activated protein kinase, mTOR = mammalian target of rapamycin, p70S6K = p70 S6 kinase, MTT = 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, dn = dominant negative, RT = room temperature, GAPDH = glyceraldehyde-3-phosphate dehydrogenase.
Differences were considered statistically significant at *P < 0.05, #P < 0.05, **P < 0.01, and ##P < 0.01.
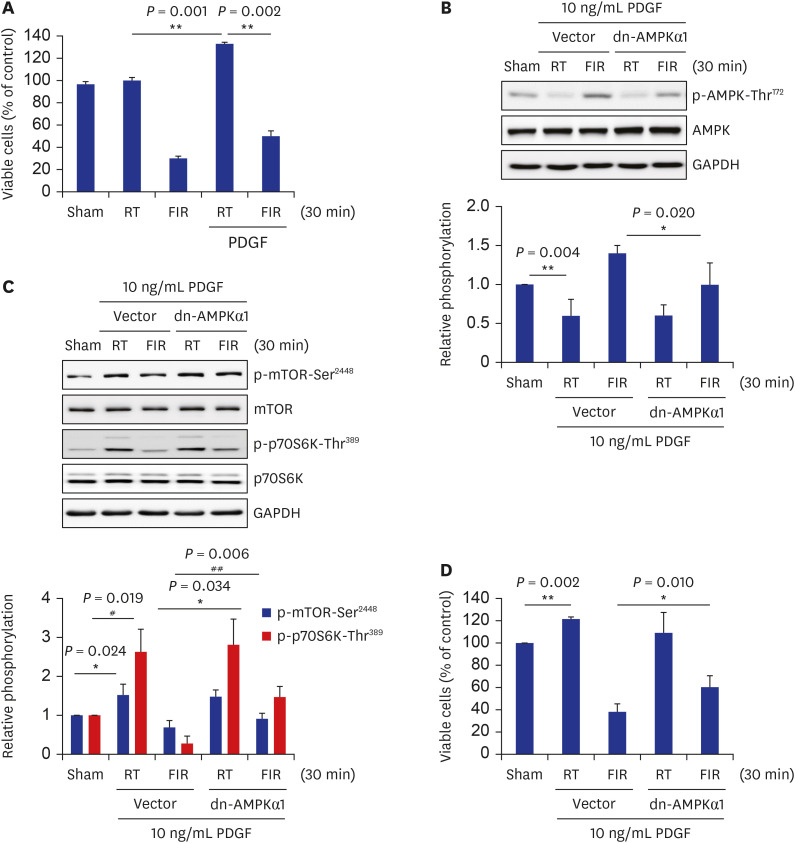
Fig. 6
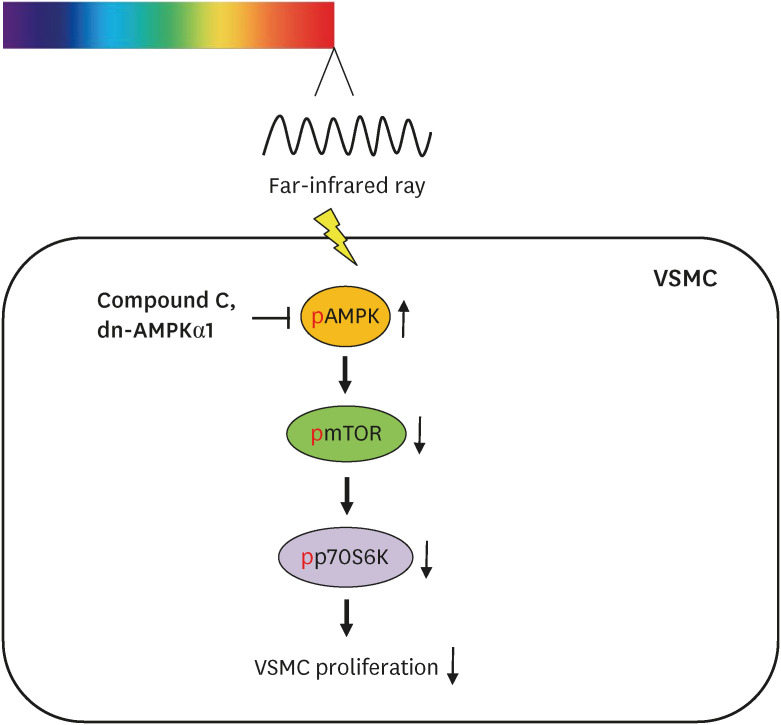
A schematic illustration of FIR irradiation-inhibited VSMC proliferation. FIR irradiation reduces the cellular ATP level, which increases p-AMPK-Thr172. Increased AMPK activity decreases p-mTOR-Ser2448 and p-p70S6K-Thr389. Finally, FIR irradiation-activated AMPK inhibits basal and PDGF-stimulated VSMC proliferation by decreasing mTOR/p70S6K signaling pathway.
FIR = Far-infrared, ATP = adenosine 5′-triphosphate, AMPK = AMP-activated protein kinase, mTOR = mammalian target of rapamycin, p70S6K = p70 S6 kinase, PDGF = platelet-derived growth factor, VSMC = vascular smooth muscle cell.
DISCUSSION
The functional dysregulation of VSMCs plays a critical role in the development of diverse vascular diseases, such as atherosclerosis.37 During early atherosclerosis, VSMCs increase not only the expression of inflammatory mediators, including tumor necrosis factor α, monocyte chemoattractant protein-1, and vascular cell adhesion molecule-1, but also the synthesis of extracellular matrix proteins, leading to the atheroma formation.3839 In particular, excessive and abnormal proliferation of VSMCs in arterial vessels has been pointed out as a paramount pathophysiological factor in the development of atherosclerosis and in-stent restenosis in patients who had undergone angioplasty.404142 Therefore, pharmacological or non-pharmacological interventions that can alleviate aberrant VSMC proliferation are considered effective strategies for preventing and treating atherosclerosis and in-stent restenosis. Therefore, these results showing that FIR irradiation inhibits cell proliferation in both basal and PDGF-stimulated VSMCs via the AMPK-mediated suppression of mTOR/p70S6K signaling axis (Fig. 6) provide scientific evidence that FIR therapy as a non-pharmacologic intervention may be useful preventing and treating arterial narrowing diseases, such as atherosclerosis and in-stent restenosis.
One notable finding in this study is that exposure of VSMCs to FIR radiation decreased the cellular ATP levels and activated AMPK (Fig. 2B and C). On the other hand, a recent study reported that FIR irradiation increases ATP production via the improved enzymatic activity of complex I in the mitochondrial oxidative phosphorylation pathway.43 Furthermore, the oxygen consumption rate, which reflects the mitochondrial respiration in cells, was increased by FIR irradiation in pancreatic insulinoma cells and neuroblastoma cells.4445 The discrepancies between the present results and previous studies are difficult to explain. On the other hand, the difference in FIR irradiation conditions may cause this discrepancy. First, the difference in peak wavelength of the FIR irradiation used in each study can result in the discrepancy. In this regard, the 7- to 12-µm FIR source, not the 2- to 5.25-µm source, promotes cell migration in the rat renal tubular epithelial cell line. Rather, the 2- to 5.25-µm FIR source decreases cell migration.46 Furthermore, the 2- to 5.25-µm FIR source does not affect the mitochondrial function, even though the 7- to 12-µm FIR source increases the basal and maximal levels of mitochondrial function in the epithelial cells.46 In this respect, Li et al.43 used a ceramic FIR radiator that can emit FIR radiation with a wavelength range from 3 to 25 µm with no specified peak wavelength. Hsu et al.45 used the 8 to 10 µm wavelength range by applying an IR bandpass filter, and Chang et al.44 used the 3 to 25 µm wavelength range with a peak value of 5–7 µm. In contrast to these groups, an FIR emitter was used with an emission wavelength range from 1 to 20 µm with a 4 µm peak wavelength. In other words, the present study used a shorter FIR wavelength than in other studies, which may have caused the discrepancy between the current results and those from other groups. Second, the difference in the intensity of FIR irradiation used in each study can lead to the discrepancy. The FIR irradiation intensity used in the above reports was 0.13 mW/cm2,434445 but the FIR intensity of 65 mW/cm2 was used in the experimental condition, indicating that the intensity of FIR irradiation in the current experiments was 500 times stronger than that in these previous studies. This difference in the intensity of FIR irradiation used among the investigations may also cause the discrepancy. Nevertheless, the intensity of FIR irradiation for 30 minutes used in our recent report26 and the current study did not induce the cell death (Fig. 1B).
A previous study using FIR irradiation under the same conditions as the current study showed that intracellular Ca2+ was translocated into the nucleus in breast cancer cells.26 In parallel with this, intracellular Ca2+ localization was changed from the cytosol to the perinucleus when the VSMCs were exposed to FIR radiation (Supplementary Fig. 4). A large amount of ATP is consumed to transport ions against the concentration gradient in cells.47 Indeed the skeletal muscles consume 40–50% of the total cellular ATP to transport Ca2+ from the cytosol to the sarcoplasmic reticulum in the resting status.48 Under normal conditions, the cellular ATP levels are determined by the sum of the ATP generation rate through oxidative phosphorylation in the mitochondria and the ATP consumption rate through various physiological processes, including Ca2+ transport. Considering these facts, the FIR irradiation applied in the current experimental condition alters subcellular Ca2+ localization at the expense of cellular ATP consumption, resulting in decreased ATP levels and AMPK activation. The current results will provide a scientific basis for applying FIR irradiation to prevent and treat metabolic diseases, such as obesity and diabetes, and arterial narrowing diseases.
The results showed that overexpression of the dn-AMPKα1 mutant gene constructs partially restored the basal and PDGF-stimulated VSMC proliferation inhibited by FIR irradiation (Figs. 3D and 5D), suggesting that FIR irradiation-inhibited VSMC proliferation is mediated by at least two independent pathways. In this regard, the decrease in p-p70S6K-Thr389 was more severe than those of p-mTOR-Ser2448 (Fig. 2A) at 30 minutes post-FIR irradiation, suggesting that a signaling pathway may be involved in inhibiting p-p70S6K-Thr389 but not p-mTOR-Ser2448. Peterson et al. reported that protein phosphatase 2A (PP2A) could associate and dephosphorylate p70S6K, and mTOR can phosphorylate and inactivate PP2A, providing two modes of p70S6K inactivation by PP2A.49 PP2A inhibited p70S6K in transforming growth factor-β-treated EpH4 mammary epithelial cells to induce G1 arrest.50 Most recently, valproic acid inhibited p70S6K activity by increasing the association of the PP2A catalytic subunit (PP2Ac) and p70S6K.36 Furthermore, PP2A inhibits p70S6K activity by increasing the binding of PP2Ac and p70S6K independently of mTOR.51 Based on the results and these reports, possible PP2A-mediated p70S6K repression is another signaling pathway that may be involved in the FIR irradiation-inhibited VSMC proliferation. On the other hand, further research is needed to clarify this issue, which is beyond the scope of this study.
It is unclear if the many beneficial effects of FIR irradiation on the body are stemmed from the hyperthermal effect of FIR ray or the peculiar effect of FIR ray independently of hyperthermia because the skin temperature increases steadily, peaking at 38–39°C, when the FIR generator is placed 20–30 cm above the skin surface.632 In accordance with the previous reports, FIR irradiation increased the temperature of the culture medium to 38–39°C under the present experimental conditions (Supplementary Fig. 3). On the other hand, hyperthermal condition (39°C) did not affect VSMC proliferation, cellular ATP level, and AMPK/mTOR/p70S6K signaling axis (Fig. 4), suggesting that the inhibitory actions of FIR irradiation were due to the peculiar effects of FIR ray, irrespective of its hyperthermal effects. In this regard, only two studies reported that whole-body hyperthermia with FIR irradiation inhibits mammary tumor growth in SHN mice and subcutaneous colon cancer growth in a subcutaneous tumor model of colon cancer, respectively.3334 In contrast, Toyokawa et al.52 reported that FIR irradiation promotes full-thickness skin wound healing in rats with no changes in skin blood flow and temperature before or during FIR irradiation. A recent study also reported that FIR therapy reduces the vascular endothelial growth factor-induced cell proliferation independent of the thermal effects in human umbilical vein endothelial cells (HUVECs).2 Furthermore, FIR irradiation inhibits the proliferation, migration, and angiogenesis of HUVECs independent of hyperthermia.27 In addition, it increases skin microcirculation through non-thermal biological effects in SD rats.32 Most recently, FIR irradiation inhibits the proliferation of human breast cancer cells independently of the hyperthermal effect.26 Considering the current results and many previous reports, FIR irradiation mainly exerts its biological effects through a non-thermal pathway. Hence, identifying the cellular receptor that mediates the biological effects of FIR irradiation in the body would warrant advances in biophysics and biomedical research.
In conclusion, these results showed that FIR irradiation-activated AMPK inhibited VSMC proliferation, at least in part, by downregulating the mTOR/p70S6K signaling axis, irrespective of the hyperthermal effect of FIR. Understanding the detailed mechanism of the inhibitory effects of FIR ray on VSMC proliferation may provide scientific evidence for its utility in preventing and treating arterial narrowing diseases, including atherosclerosis and post-intervention restenosis.
 XML Download
XML Download