

 PDF
PDF Citation
Citation Print
Print



Introduction
Back pain is the most common, chronic, and discomforting condition associated with aging and consequent progressive joint dysfunction [1, 2]. Back pain arises from functionally impaired intervertebral discs [3], which are load-bearing viscocartilaginous tissue lying between the consecutive vertebral bodies allowing moderate spinal movements [4]. Anatomically, each intervertebral discs has three integrated parts: the centrally placed gelatinous nucleus pulposus (NP), the laminated annulus fibrosus (AF), and the hyaline cartilage endplate regions [5-7]. The NP, IAF and outer annulus fibrous (OAF) regions contain two distinct cell types which are described as rounded cells of the nucleus pulposus and the inner annulus regions, and the highly flattened cells of the outer annulus [8, 9]. Recent and past evidence suggests that the nucleus pulposus cells proliferate and may have progenitor capabilities with tissue repair functions [10]. However, cell clusters are also a conventional hallmark of disc degeneration and catabolism, as the disc cells, particularly those in clusters produce cytokines that can trigger catabolic activity without the involvement of immune cells [11, 12]. Cell clusters undergo stress induced premature replicative senescence involving a brief phase of proliferation followed by growth arrest while remaining metabolically active during disc degeneration [13, 14]. Therefore, the activity of nucleus pulposus and the inner annulus cell phenotypes is uncertain, and a clearer distinction between proliferating reparative disc cells, non-proliferating, or proliferating degenerative cells still lacks, and a thorough investigations could assist in development of strategies that can promote endogenous cell-based repair techniques.
In young and mature non-degenerated intervertebral discs, nucleus pulposus cells appear singly, in pairs, or in small cell clusters, whereas with age and degeneration single cells or small cell cluster often progress to form medium and then large sized clusters [15] which is considered as the most common histological sign of disc degeneration. These large cell clusters are confined to the nucleus pulposus and inner annulus regions [9], and may contain as many as 60–100 individual cells in a single large sized cluster [8] presumed to arise from proliferation [9, 16], although a few studies suggest that cell migration may be another process involved with formation of clusters [17, 18], furthermore, proliferating or migrating cells, are influenced by cytokines and chemokines overexpressed by the cell themselves during degeneration to initiate catabolism [19].
Animal models of osteoarthritis shows, chondrocyte cell proliferation is also associated with increased proteoglycan synthesis representing a repair response to tissue damage [20, 21]. Similarly, cyclic mechanical compression increases proteoglycan and collagen production by the nucleus pulposus cells [20] allowing cell clustering [22, 23]. There is also evidence that disc cell proliferation is initatied to faciliate repair due to stimulation by diffusible growth factors [24], which is in contrast to stimulation through neuropeptide such as Substance-P that acclerates the release of cytokines [25, 26].
Spatial associations studies between disc cell clusters and histological degenerative features indicates [27] that highly crosslinked disc matrix is beyond the repair capacities by the cells, as the main anabolic mediator produced by these cells are proteoglycan molecules and fibrillar collagen which are structurally weak. This problem could be exacerbated by the relative avascularity, accelerated nutritional depletion, and accumulation of toxic metabolites such as lactate [28, 29] promoting premature replicative senescence and apoptosis, of the otherwise viable cells. Large clusters of senescent cells can also possibly synthesize sufficient proteolytic enzymes to cause focal matrix breakdown and allow fissure formation [11].
Cells of the nucleus pulposus, in contrast, has also been shown to form colonies ex-vivo and are immunopositive to stem and progenitor cell markers such as GD 2, a disialoganglioside, identified as a marker for bone marrow and umbilical cord mesenchymal stem cells. Tie-2, a tyrosine kinase receptor is localised in the disc cell membrane, that forms ligand with angiopoietin-1 to maintain the progenitor potential of the cell and protects it from apoptosis [30, 31]. Such progenitor nucleus pulposus cells differentiate into adipocytes, osteocytes, and chondrocytes in cell culture [32], and can be extracted from the intact discs [33]. This aforesaid evidence indicate that phenotypic heterogeneity exists within the nucleus pulpous cells, which may differ during early, intermediate, and advanced stages of disc degeneration. These undefined parameters may influence the ontogeny and function of the nucleus pulposus cells. Therefore, cell proliferation markers such as proliferating cell nuclear antigen (PCNA) and Ki-67 can identify proliferating cells clusters and define the differences in cluster sizes seen during the various grades of disc degeneration. Thus, we objectively performed an immunohistochemical and protein quantification sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) investigation to analyse such differences.
Go to : 
Materials and Methods
Human tissue
Ethical clearances were obtained from National Research Ethics Service (NRES17/SW/0112), Frenchey Hospital, North Bristol NHS Trust, UK, and Sikkim Manipal Institutional Ethics committee (IEC/SMIMS/2018/174-24), Ministry of Health and Research (F.NP.L.19015/53/97-IH) Govt., of India, prior to the human tissue sample collection process. Experimental protocol/methods were approved by Institutional Research Committee of the University of Bristol, UK and Sikkim Manipal University, India. The method described is in accordance with the guidelines mentioned by the World Medical Association, Declaration of Helsinki for medical research. Written informed consents were obtained from the patients for collecting the waste disc tissue. Fifty-five human disc tissues were collected from patients aged 37–72 years undergoing spinal surgeries. 19 patients had slightly degenerated non-herniated discs, or degenerative scoliosis disc with <45° deformity; 21 patients had severe degenerated discs with herniation; and 15 were non-degenerated cadaveric control discs. An anonymous clinical data sheet was obtained for each patient, and magnetic resonance imaging (MRI) scans were graded 1–5 as according to the clinically relevant Pfirrmann (pf) grade of disc degeneration [34]. Data are summarized in Table 1.
Table 1
Summary data for human discs used in this study
![]()
Semi-quantitative histological analysis of human discs
Each disc was divided into two halves in the sagittal plane. One-half was snap-frozen in isobutane, embedded in optimal cutting tissue medium (OCT), five-micron thin sequential sections were obtained on a Leica S-320 cryostat, and fixed in 10% neutral buffered formalin. Ehrlich’s Haematoxylin & Eosin stain was used for the assessment of cells and the matrix. Consecutive sections were stained with toluidine blue to assess proteoglycans loss [35]. Outer annulus, inner annulus, and nucleus pulposus tissue was distinguished from each other following a published protocol [8]. Using the modified criteria of Boos et al. [36] histological features were graded on an ordinal scales: cell clustering (0–5), inflammatory cells (0–3), proteoglycan staining score (0–3), and presence of tears/fissures in the matrix (0–3). Inflammatory cells were identified by observing its characteristic shape, organization, and localization around blood vessels. In each case, ‘0’ and ‘3’ (or 5) were referred to as ‘absence’ or ‘abundance’ of the scored variables as analyzed with Table 2. Scores were averaged across three fields of view, and then across specimen groups.
Table 2
Variables for histological assessment
Based on modified Boos et al. [36].
![]()
Quantitative immunohistochemical analysis
Five-micron thin sections were fixed in acetone for 10 minutes at –20°C, washed in phosphate buffer saline (PBS) and all non-specific binding sites were blocked by application of 5% of rabbit or donkey serum at 4°C for one hour (1:5 dilutions in PBS). Sections were washed in PBS and applied with monoclonal primary antibodies to matrix metalloproteinase-1 (MMP-1) (Ab52631; Abcam), Caspase-3 (31A1067; Novus Biologicals), Ki-67 (Novus biologicals, MK167-8D5), and PCNA (PC10; Novus biologicals) at 1:40 dilutions in PBS, and PBS-immunogloblin G (IgG) alone as negative controls. Sections were incubated overnight to allows maximum interaction between the antigen of interest and primary antibodies. The next day, sections were washed in PBS and incubated with appropriate Alexa fluorophore secondary antibody at 1:100 dilutions in PBS for one hour, after PBS washes auto-fluorescence was quenched using 0.1% Sudan black B for 12 minutes. Sections were counterstained with Vecta Shield Dapi (H-1200; Vector), and secured with a cover slip [37].
Quantitative analysis was done with Leica-Olympus DP72 (Olympus) camera (12.8 megapixels) with a fixed frame view of 650 µm×500 µm. Positively stained cells in clusters for MMP-1, Caspase-3, Ki-67, and PCNA were counted across the entire section at a magnification of 20×, with an average of 8 fixed frames per section. Ten 5-µm sections were analyzed for each disc sample. Immuno-positive images were counted with the cell sense software package.
Protein extraction and SDS-PAGE analysis
The remaining half of each human disc was rinsed in phosphate buffered saline, finely diced and proteins were extracted at 4°C under continuous agitation for 72 hours using 15 volumes of 4 M guanidine hydrochloride (Sigma Aldrich), 50 mM sodium acetate and 10 mM EDTA (Sigma Aldrich). The solution was adjusted to 5.8 pH, and protease inhibitor was added before introducing the diced disc samples into the extraction buffer. The extract was separated from the tissue by centrifugation in 13,000 g, and aliquots of 8 µl were prepared for SDS-PAGE. This solution was digested with Keratanase (Ambsio LLC) at 1 mU/25 µl for 6 hours, further treated with 100 mM Tris-HCL, ph 7.4 and digested overnight with chondroitinase ABC (Ambsio LLC) at 50 mU/25 µl of the extract.
SDS-PAGE
Lamelli buffer containing 2× sodium dodecyl sulfate, 2 mercaptoethanol, and milliQ double filtered water was added to the sample and heated in a water bath at 95°C for 10 minutes before being fractionated on 10% polyacrylamide gels at 94 V (constant voltage) for 1 hour 40 minutes. Proteins of interest were transferred to nitrocellulose membranes (0.45 µm) using transfer buffer with 20% methanol at 100 V (constant voltage) for 1 hour. Membrane was blocked with 5% skim milk powder in 0.01M Tris-HCL, 0.15 M NaCl, 0.1% Tween20 (TBST), ph 7.6. Monoclonal antibodies to MMP-1, PCNA, Ki-67, and Caspase-3 diluted in 1: 1,000 TBST with 1% BSA, were applied to the blots for 2 hours, followed by TBST washes. Appropriate secondary conjugated antibody to horseradish peroxidase (Thermo Fisher Scientific) was applied at 1:1,000 dilutions, followed by TBST washes. Protein bands were developed by chemiluminescence with an ECL mixture (GE Healthcare). Blots were also analyzed omitting primary antibody to check for IgG species in the tissue extracts.
Quantification of western blot bands with Image J
Protein blot images were saved at 600 dpi in gray scale, each protein lane (row-wise) was then measured by selecting a single region of interest with a rectangle tool from the Image J measurement tool bar (https://imagej.nih.gov/ij/download.html) to define a frame around the largest band of that row. The ratio of each protein band to the loading control was analysed by inverting the pixel density of the band as 255-X, where X was the value recorded for each protein band and the loading control.
Statistical analysis
Spearman’s rank correlation was used to examine associations between histological variables. One and two-way ANOVA was used to compare mean values of cell clusters within the NP, IAF, and OAF regions, and to compare cell counts of positively stained Ki-67, PCNA, MMP-1, and Capase-3 in the patient and control groups. Statistical tests were performed using SPSS software version 19 package (IBM Co.). P<0.05 was considered as statistically significant.
Go to :
Results
Frequency of cell clustering in NP, IAF, and OAF regions
The OAF region of the control discs comprised of parallel and crimped collagen fibers along with elongated and extremely flattened fibroblast-like cells, with almost indistinct cytoplasm (Fig. 1A). Cell clusters were always absent in the outer annulus of the control tissue but were a prominent feature of severely degenerated and herniated discs with structural defects, inflammatory cells, and disrupted collagen lamellae (Figs. 1B, C, 2B, D). Infiltrating inflammatory cells and blood vessels were localized around the vicinity of fissured areas in herniated mixture of nucleus and annulus tissue (Fig. 1B). Nucleus pulposus and inner annulus regions showed single cells, cells in pairs, in small size clusters (3–7 cells), medium size clusters (8–15 cells) in non-degenerated control cadaveric disc (Fig. 2A, C). Large-sized clusters appeared in a meshwork of loosely organized collagen fibres without any crimp, and these large clusters (>15 cells) were a feature of herniated, severely degenerated discs, localised in the NP and IAF junction (Fig. 2B, D). Clustering cells identified through histological scoring increased with pf grade of disc degeneration (Fig. 2F) in the examined tissue sections (rs=0.316, P<0.04).
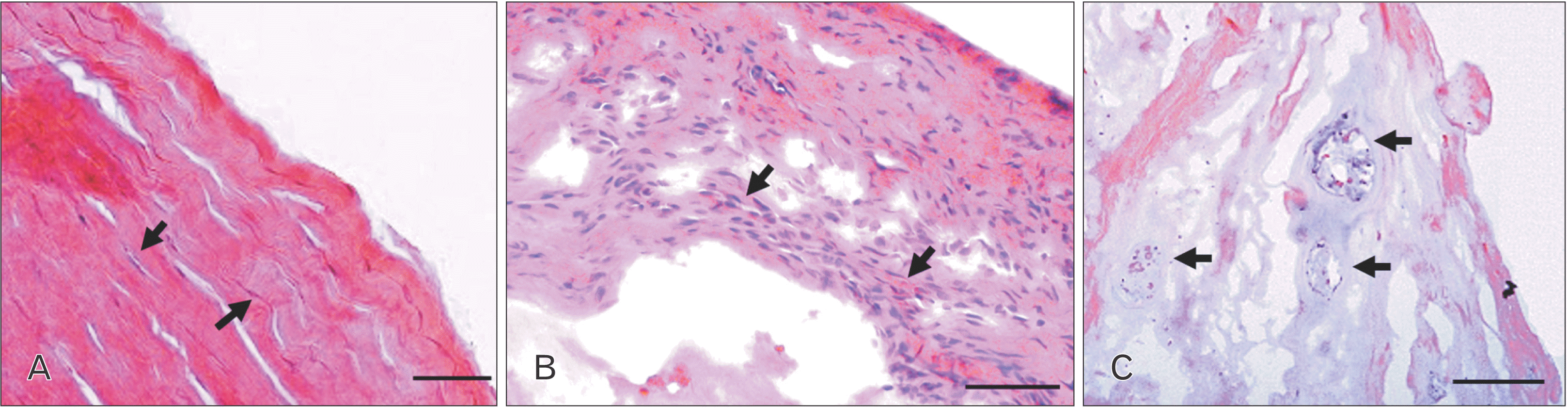 | Fig. 1Comparisons between (A) control non-degenerated cadaveric human disc with extremely flattened annulus fibrosus (AF) cells (arrows) and (B) herniated disc, arrows shows inflammatory cells, and (C) degenerated disc with displaced lamellae and large cell clusters (arrows). Scale bar: 100 μm.
|
 | Fig. 2Cell clusters comparisons between (A) control non-degenerated human cadaveric disc with small size cell clusters (2, 3, 6 cells) and (B) herniated disc with large sized cell clusters (50–60 cells, arrow). Scale bar: 100 μm (A) and 50 μm (B). (C) Toluidine blue stained control NP/IAF region without clusters and (D) herniated disc tissue with large cluster (arrow) stained with toluidine blue. Scale bar: 50 μm. (E) Cell cluster scores (0–5) and increasing pf grade of disc degeneration. (F) Proteoglycan staining score (0–3) and increasing pf grade of disc degeneration. NP, nucleus pulposus; IAF, inner annulus fibrosus; pf, Pfirrmann grade of degeneration; PG, proteoglycans.
|
Toluidine blue staining to assess proteoglycans loss
Proteoglycan staining score was greatest in control discs, the NP and IAF regions, and decreased with severity of degeneration, Table 3. Staining intensity reduced around invading inflammatory cells, large clusters, especially in pf grade 4, 5 degenerated discs (Fig. 2D, F). Proteoglycans loss scores correlated with the presence of fissures in the OAF (rs=0.85, P<0.001), IAF (rs=0.53, P<0.001) and NP regions (rs=0.74, P<0.001). Large clusters showed little or no purple-stained proteoglycan matrix, and often exhibited nuclear shrinkage or ‘pyknosis’ (Fig. 2B), a characteristic feature of cells that had lost its ability to divide. Herniated discs showed greatest proteoglycans loss in comparison to moderately degenerated and control discs (Fig. 2C, D, Table 3).
Table 3
Summary of results for human discs, comparing three tissue regions NP, IAF, and OAF in three groups
NP, nucleus pulposus; IAF, inner annulus fibrosus; OAF, outer annulus fibrosus; PG, proteolgycan; MMP-1, matrix metalloproteinase-1; PCNA, proliferating cell nuclear antigen.
![]()
Immunohistochemical localization and quantification of MMP-1, Ki-67, PCNA, and Caspase-3
Patient discs showed selective cells in clusters that were positive to MMP-1, Caspase-3, Ki-67, and PCNA, Table 3.
MMP-1 positive cells were selectively stained in clusters (Figs. 3A, 4B) and localized around the vicinity of disrupted tissue region in the NP-IAF junction as determined by phase-contrast (Fig. 4A, B) in severely degenerated and herniated discs. MMP-1 was also found to be localized around infiltrating inflammatory cells (Fig. 4B). Control discs showed only a few MMP-1 positive cells (Fig. 3A).
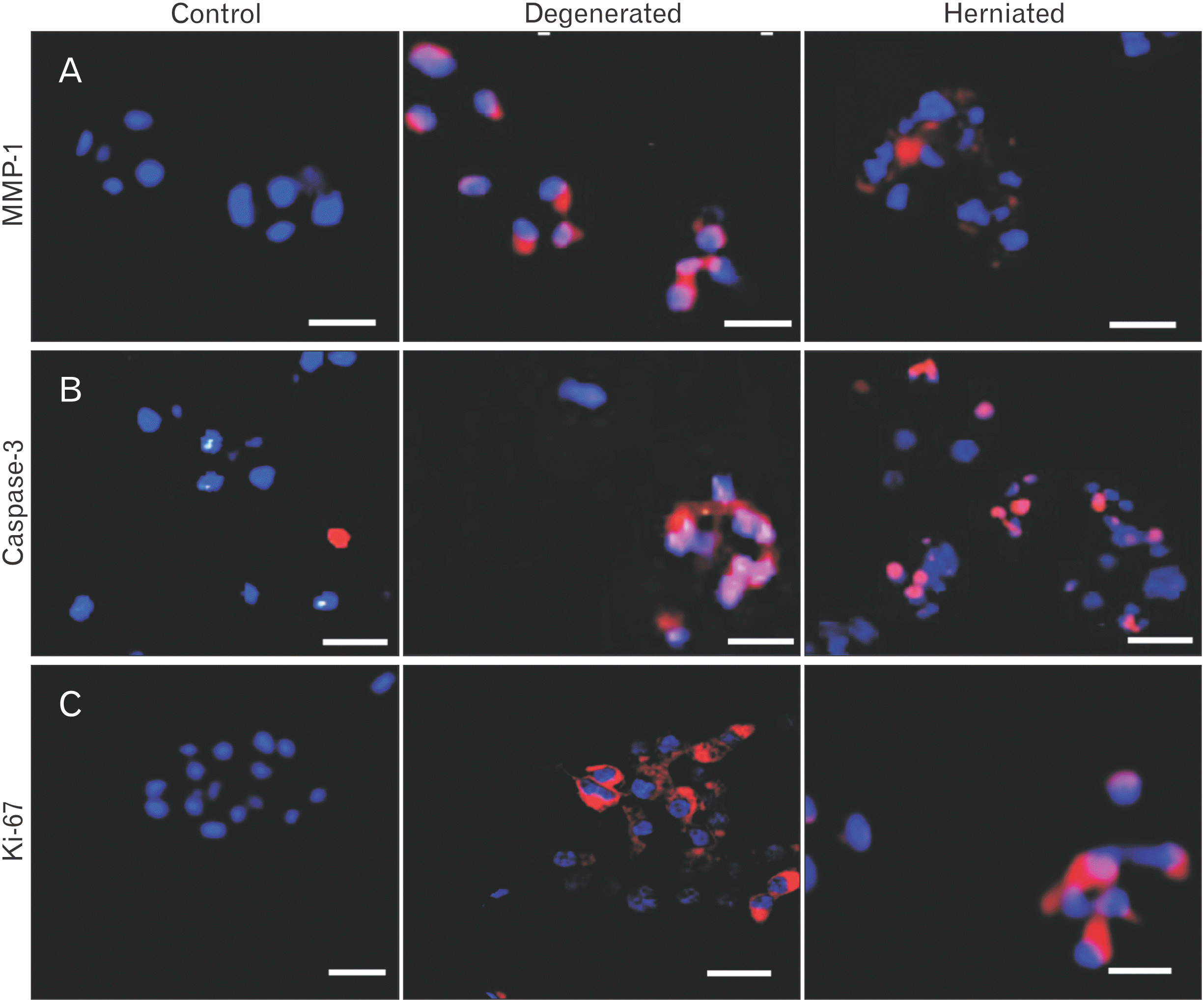 | Fig. 3Small, medium, and large sized cell clusters of the ‘control’ and its comparisons to ‘degenerated’ and ‘herniated’ discs for (A) MMP-1, (B) Caspase-3, and (C) Ki-67. Scale bar: 50 μm. MMP-1, matrix metalloproteinase-1.
|
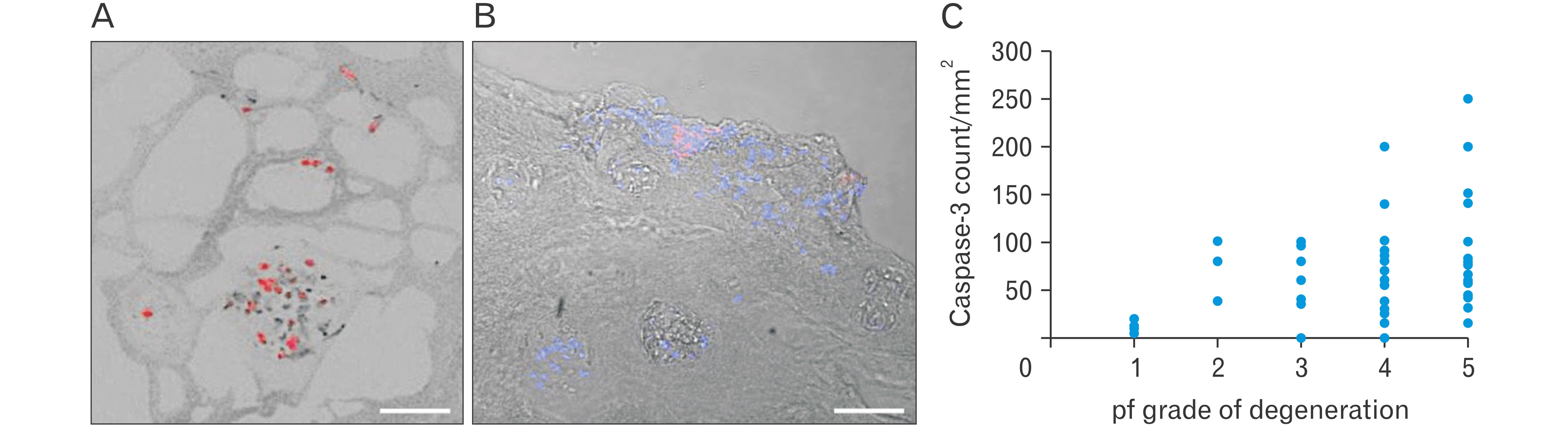 | Fig. 4Medium and large sized cell clusters stained with (A) Caspase-3 (red stain, blue indicates DAPI stained cell nuclei), (B) MMP-1 (red stain, blue indicates DAPI stain cell nuclei) with phase contrast background. Scale bar: 100 μm. (C) Caspase-3 positive cell clusters count and increasing pf grade of disc degeneration. MMP-1, matrix metalloproteinase-1; DAPI, 4’,6-diamidino-2-phenylindole; pf, Pfirrmann.
|
Caspase-3 positive clusters, of all sizes, increased with pf grade of degeneration, were most common in herniated and degenerated discs (Figs. 3B, 4A), frequently located in disrupted inner annulus region (Fig. 4C). Very few clusters (4%) were positive in control discs.
Ki-67 positive clusters were a feature of moderately degenerated discs and controls. Herniated and severely degenerated discs showed fewer Ki-67 and PCNA positive cells (Fig. 3C). Ki-67 and PCNA positive clusters did not increase with pf grade of disc degeneration.
Ki-67 staining was visualized as non-nuclear, which is in contrast to the general acceptance that Ki-67 and PCNA proliferative markers must always be a nuclear stain, however, recent evidence indicates non-nuclear cytoplasmic staining of Ki-67 and PCNA is common in mature neutrophils [38], breast carcinoma cells [39], and hyalinizing trabecular adenomas of the thyroid and sclerosing haemangiomas of the lung [40]. Such non-nuclear localization can be explained as cells in discs tissue are predominantly non-mitotic and don’t proliferate, but during degeneration to compensate for proteoglycans loss, cells begins to form clusters of various sizes and relatively non-dividing cells proliferate to as many as 20–100 cells within a large clusters which can translocate Ki-67 and PCNA to subcellular, cytoplasmic, and membrane bound locations either due to point mutation or deletions of the nuclear export signals resulting in molecular remodeling of its cellular and subcellular components [41].
All patient discs additionally also contained fewer numbers of single, paired cells, and flattened annulus cells, that were positive to MMP-1, Caspase-3, Ki-67, and PCNA.
SDS-PAGE Quantitative analysis for MMP-1, Ki-67, PCNA, and Caspase-3
Protein extracts from human disc tissues along with non-degenerated control and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as loading control were analyzed on a weight per volume basis by gel electrophoresis. Image J analysis software were used to quantify the bands.
MMP-1 was expressed as a weak band in control discs, while degenerated and herniated discs showed prominent bands at 54 kDa (Fig. 5A). MMP-1 expressions gradually increased with pf grades of degeneration and were most expressed in grade 3 discs (Fig. 5B), followed by decline in grade 4 and grade 5 discs. Control, grade 1 and grade 2 disc also had similar low MMP-1 expression profile perhaps to prevent the tissue from self-proteolysis.
 | Fig. 5Western blot analysis on a 10% gel. Lane were loaded as C, ‘non-degenerated cadaveric disc’ followed by disc specimens categorised according to pf grades degeneration in the subsequent lanes as pf grade 1, 2, 3, 4, and 5 discs. (A) MMP-1 core protein at 54 kDa. (B) Expression level of control ‘c’, and grade ‘1–5 discs’ shown with grade 3 discs at the highest expression level. (C) Ki-67 core protein with a molecular weight of 250 kDa. (D) Weak expression level for degenerated grade 3–5 discs. (E) PCNA core protein with a molecular weight of 36 kDa. (F) Bands expressed higher values for grade 2 and 3 discs in comparisons to other grades. (G) Caspase-3 core protein at 22 kDa. (H) Strongly expressed for the severely degenerated grade 4 and 5 discs. GAPDH as loading control is at 36 kDa. C, control; pf, Pfirrmann; MMP-1, matrix metalloproteinase-1; PCNA, proliferating cell nuclear antigen; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
|
Ki-67, PCNA, and Caspase-3 were expressed moderately in control discs (Fig. 5 C–H). Ki-67 appeared as a 350 kDa band and were prominently expressed in grade 1 and 2 discs followed by grade 3, 4, 5 discs (Fig. 5C, D). PCNA, appeared at 36 kDa and followed the same expression pattern as Ki-67 (Fig. 5E, F) suggesting that grade 2, 3 degenerated disc tissue had greater quantity of viable cells. Caspase-3, appeared as a 22 kDa band, and were minimally expressed in controls, grade 1–3 discs, followed by increased expression in severely degenerated grade 4, 5 discs (Fig. 5G, H).
Go to :
Discussion
Summary of results and gap analysis
The present study shows small and medium-sized clusters in control and slightly degenerated discs express Ki-67 and PCNA markers whereas large clusters localised around disrupted proteoglycan-depleted tissue in the NP/IAF junction of severely degenerated, herniated discs are positive to MMP-1 and Caspase-3. These findings were confirmed with SDS-PAGE that shows these target markers can be extracted from the relatively dense collagenous matrix. SDS-PAGE investigation allowed segregation and identification of these protein of interest through its molecular weight, thereby eliminating any interferences which may be caused due to non-specific binding charges introduced by immuno-histochemical methodologies. Small and medium-sized cell clusters had higher proportions of cells positive to proliferating markers, while caspase-3 activity in large clusters were common in end-stages of disc degeneration. Control disc showed least number of cell clusters, proteoglycan loss score, and immunopositivity towards MMP-1 and Caspase-3. Large clusters, in contrast, were a characteristic feature of severely degenerated and/or herniated disc with increased Caspase-3 activity [42].
This study compared pathological human discs with age-matched cadaveric controls. Ki-67 and PCNA antigen were the operational markers used for determining cellular proliferation. Ki-67 is not expressed by quiescent or resting cells in the G0-phase [43] and PCNA markers are expressed only during the S phase of the cell cycle [44], hence a combination of these two markers facilitated the assessment of cell proliferation phase in the clusters of various sizes. MMP-1, is the main enzymatic marker involved in initiation of extracellular matrix degradation and is suggested to play a vital role in increased proteolysis of the disc matrix [42] and leukocyte migration. MMP-1 cleaves collagen types I, II, III and proteoglycans which forms the main microstructural framework of the intervertebral discs [45]. Similarly, other ubiquitous matrix metalloproteinases such as MMP-2, 3, 5, 7, 9, 13 [46] are also shown to be involved in diverse functions associated with tissue injury, repair, inflammation, blood vessels and nerve ingrowth [8, 47].
Caspase-3, is involved in the activation cascade of apoptotic cell death pathway in human intervertebral discs [43]. At the onset of apoptosis, it proteolytically cleaves poly (ADP-ribose) polymerase at ‘216-Asp-Gly-217’ bond which shows caspase 3 is a key cysteine protease that executes programme cell death. Caspase 3 can also activate caspases 6, 7, and 9 that overlap with the necrotic mode of cell death [47, 48], a prominent feature of inflammation and degeneration. Caspase-3 antibody used in this study identified both the pro-caspase form (32 kDa) and the active caspase form (14–22 kDa).
This study made comparisons only between human tissue, and even though the results are applicable to the human population, a controlled animal tissue study along with human discs samples would perhaps be a more insightful prospect, however animal are quadrupled species and unlike humans their discs are more complex to load and would involve ethical concerns for in vivo loading.
Relationship to other studies
With age chondrocyte-like cells of the nucleus and inner annulus regions become more and more rounded or even irregular in shape, develop a dense pericellular matrix, and frequently proliferate to form cell clusters [15, 49]. Each small clusters consists of 3–4 cells within a lacunae that isolate them from the rest of the matrix [24] large cell clusters with <8–15 cells are considered to be a histological hallmark feature of cartilage tissue degeneration [9], but cell cluster are also hypothesized to represent an attempted repair response to tissue damage, where remodeling of matrix occur via chondrocyte division or proliferation of cells, swelling of the chondrons, and distension of the surrounding microenvironment [50]. Since cell clusters are abundantly located in regions that show loss of fibrillar orientation [51], or are located mostly near lesions in the experimental animal model [52] they could be indicative of a repair response. Cell proliferation are considered to be an essential process for maintaining hydration of tissue, and organ culture studies of human discs show that native disc cells produce large proteoglycan aggregates [53] that play a crucial role in maintaining swelling pressure of the disc tissue [54], but with aging, accelerated degeneration, and pain, changes in the morphological and biochemical structure of the collagen lamellae, and vital cellular or non-cellular protein components increases cell clustering. Anabolic factor such as the proteoglycans molecules produced by the cells and clusters, as a repair response, can easily leach out of the disc due to excessive tissue swelling and loss in restrain especially in grade 4 and 5 discs [11]. Other significant problem associated with tissue repair is decline in the number of viable disc cells due to poor metabolite transport [55], subsequently increasing the proportions of cells that undergoes senescence [13], followed by apoptosis [56]. Thus, large, and medium sized cell clusters formed during degeneration, and in aged individuals, may contribute towards increased release of matrix-degrading enzymes [37], capable of altering the mechano-transduction pathways [29]. Moreover, unavoidable daily loading stress, age-related changes of the intrinsic and extrinsic cell-matrix environment, and genetic factors may progressively weaken the disc matrix, and structural changes, such as fissure formed, may not be easily repaired by the temporarily proliferating cell. Thus, with age and injury, cell clustering is perhaps unavoidable, especially near the nucleus pulposus and inner annulus junction where clusters were abundant [8, 11], and immunoreactivity towards MMP-1 and Caspase-3 are located within the same region. This finding are additionally in agreement with mechanical studies that show the inner annulus is weak and prone to structural disruptions due to repetitive loading and ageing [3]. Current and past studies on humans, porcine and ovine discs confirm structural failure always leads to progressive degenerative biochemical changes [52], but are not seen in non-degenerated disc [57]. Thus, findings from this study suggest cell clustering is initiated as tissue repair or remodeling process that fails due to unsupportive metabolic, biomechanical, and biochemical responses, and these cell cluster eventually undergoes apoptosis.
Explanation of results and conclusion
Proteoglycan-depleted tissue regions with large cell clusters indicates phenotypic alteration perhaps due to abnormal mechano-transduction pathways [58], and degenerated, painful discs exhibit negative regulation to nuclear factor-κB pathway [59] influencing the proliferation process. Increased MMP expression contributes towards focal tissue disruptions by allowing enzymatic fragmentation of matrix that may activate TLR receptors [60, 61], and these fragmented molecules can acts as damaged associated molecular pattern (DAMPS) which acts in a similar ways as the pathogen associated molecular pattern (PAMPS) to initiate a sterile inflammatory catabolic cascade. Therefore, to conclude, distinctive microscopic changes and mechanical loading can cause the discs to enter ‘vicious cycle’ of interconnected mechanisms such as focal swelling that accelerates proteoglycan loss and increase cell clustering in an attempt to compensate proteoglycan loss, but as this attempt fails, clusters increase the expression of MMP-1 and caspase 3. Therefore, clusters appearing during late stages degeneration can’t remodel the matrix and ‘senolytic drugs’ proposed to clear out non-functioning cells could possibly benefit the end stages of disc degeneration.
Go to :
 XML Download
XML Download