

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Ankylosing spondylitis (AS) is a common autoinflammatory disease that affects the axial skeleton and is characterized by excessive osteoblast activity and new bone formation at local sites of entheses [1,2]. Clinically, anti-tumor necrosis factor (TNF) and anti-interleukin-17A therapies reduce inflammation in patients with AS, but they do not inhibit radiographic progression [3]. Thus, an understanding of pathological osteoblasts is needed to develop a specific therapeutic target for bony ankylosis in AS patients.
Bone morphogenic proteins (BMPs) are originally identified as growth factor morphogens that are tightly involved in the cascade of skeletal development and bone formation. BMPs can induce diverse signaling pathways in cells. Among these pathways, the BMP-SMAD protein signal is well-known. Upon BMP binding, the serine–threonine kinase activity of type I BMP receptors (BMPRs) fully activates the phosphorylation of smad1/5/8 protein to translocate it to the cell nucleus. Subsequently, the phosphorylated smad1/5/8 protein accumulates to upregulate the transcription level of BMP-response genes. BMPRs forms of type I and 2 type II kinase receptors oligomerize [4]. Many studies suggest that type I BMPRs are a general component of BMP signaling, yet the functional role of type II BMPRs remains largely unknown.
BMP2 is strongly associated with initiation and progression in AS [5-8], and an association between BMP2 variants and susceptibility to AS was reported [9]. Shen et al. put forward the notion that BMP2 has significant effects on the proliferation and differentiation of human mesenchymal stem cells of AS, and upregulated BMPR1A expression triggers fat metaplasia to form new bone in AS [10,11]. Intriguingly, a genetic study revealed that BMPR2 variants are strongly associated with radiographic changes in AS [12]. Additionally, it appears that high expression of BMPR2 in adipocytes has strong osteogenic potency [13]. However, it remains unclear whether extracellular BMP2 agonists discriminate BMPR2 and which smad1/5/8 protein signaling pathways are activated by BMP2 in AS.
MATERIALS AND METHODS
Patients
This study was carried out in accordance with institutional guidelines and approval from the Ethics Committee of Hanyang University Seoul Hospital with written informed consent provided by participants (IRB 2014-05-002). Facet joints were obtained from 14 patients (all male; mean age, 42.4±8.0 years) diagnosed according to the modified New York criteria [14] and 10 patients (all male; mean age, 56.3±11.2 years) with non-inflammatory spinal diseases as healthy control (HC). The clinical characteristics of human facet joint samples are shown in Table 1.
Isolation of human osteoprogenitors (OPs) from facet joints
Facet joints were cut into ≤1 cm bone chips with scissors. The bone chips were washed in serum-free Dulbecco’s modified Eagle medium (DMEM) (SH30243.01; Hyclone Laboratories, Logan, UT, USA) containing 1% penicillin–streptomycin (15140122; Gibco Laboratories, Gaithersburg, MD, USA) antibiotics and incubated in DMEM growth medium containing penicillin–streptomycin and 10% fetal bovine serum (15140122; Gibco Laboratories) at 37°C for 2 weeks to isolate primary OPs. Cell suspensions were filtered through a nylon mesh, washed in serum-free DMEM several times, and seeded for culture. All isolated OPs were assessed with mycoplasma negative using a polymerase chain reaction (PCR)-based method (Takara, 6601).
RNA sequencing
Two HC-OPs and two AS-OPs were involved in total RNA extraction and RNA sequencing. Data analysis was carried out by EBIOGEN Inc. (Seoul, Korea). Briefly, RNA library construction was performed using the QuantSeq 3’ mRNA-Seq Library Prep Kit (Lexogen, Inc., Greenland, NH, USA) according to the manufacturer's protocol. Analysis of differentially expressed genes andgene ontologieswas carried out using the Microsoft Excel–based Differentially Expressed Gene Analysis (ExDEGA) software package provided by EBIOGEN Inc. Heatmap generation and clustering analysis were performed using R (version 3.6.0; R Foundation for Statistical Computing, Vienna, Austria) and MeV (version 4.9.0) for selected genes.
Real-time quantitative PCR (RT-qPCR) and immunoblotting
RNA and protein extractions were completed as previously described [15,16]. RNA and proteins were extracted from stimulated cells with Trizol (15596026; Thermo Fisher Scientific, Waltham, MA, USA) and 1× radioimmunoprecipitation assay buffer, respectively. Complementary DNA was generated from 1 μg of total RNA with reverse transcriptase (#EP0442; Thermo Fisher Scientific). The cells were lysed with 1× radioimmunoprecipitation assay buffer containing phosphatase (5870S; Cell Signaling Technology, Danvers, MA, USA) and protease (#535140; Calbiochem, San Diego, CA, USA) inhibitors. Proteins were quantified with a Bradford assay. A total of 30 to 50 μg of protein was subjected to immunoblotting.
We used the following RT-qPCR primers: RUNX2 forward, 5’-TGAGCTGAGAGGACATATGGCC-3’; RUNX2 reverse, 5’-TAGACACCAAACTCCACAGCCC-3’; OCN forward, 5’-ATGAGAGCCCTCACACTCCT-3’; OCN reverse, 5’-CTTGGACACAAAGGCTGCAC-3’; BMPR1A forward, 5’-CAGCATTCGATGGCTGGTTT-3’; BMPR1A reverse, 5’-TCTAAGGACAACAGGCACGC-3’; BMPR1B forward, 5’-GCCCAGTGACCCCTCTTATG -3’; BMPR1B reverse, 5’-GTTTGGGAATGAGGGGCGTA -3’; BMPR2 forward, 5’-CCACTAGAAGGTGGCCGAAC -3’; and BMPR2 reverse, 5’-TCACCTATCTGTATACTGCTGCC -3’; ALK1 forward, 5’-CTCTGCCTACCACCTCCTCT-3’; ALK1 reverse, 5’-CAAGCTGGTGGGCTTGTTTC-3’; ALK2 forward, 5’-AATCCCCGAGACGTGGAGTA-3’; ALK2 reverse, 5’-TTCCCGACACACTCCAACAG-3’.
The antibodies used for immunoblotting and immunofluorescence were as follows: RUNX2 (12556; Cell Signaling Technology), BMPR2 (sc-393304; Santa Cruz Biotechnology, Dallas, TX, USA), ALK1 (AF370; R&D Systems, Minneapolis, MN, USA), phos-smad1/5/8 (sc-12353; Santa Cruz Biotechnology), total-smad1/5/8 (sc-6031-R; Santa Cruz Biotechnology), OPG (sc-390518; Santa Cruz Biotechnology), phos-ERK (5683; Cell Signaling Technology), phos-p38 (9215; Cell Signaling Technology), and GAPDH (2118; Cell Signaling Technology).
Immunohistochemistry
Immunohistochemistry was performed as previously described [15,16]. The facet joint samples were fixed in 10% formalin for 1 week, dehydrated, and embedded in paraffin. Tissues were sectioned into 5-μm-thick slices and used for immunohistochemistry for BMPR2. The images were obtained using an Eclipse microscope (Nikon, Tokyo, Japan) with transmitted light at 4× and 20× magnifications.
Osteogenic differentiation and quantitative methods
Osteogenic differentiation and its quantitative methods were previously reported [16-20]. Briefly, primary OPs from the facet joints were seeded in DMEM growth medium (SH30243.01; Hyclone Laboratories) and then differentiated with osteogenic medium (ascorbic acid, β-glycerophosphate, and dexamethasone) into mature osteoblasts for indicated days. Matrix maturation of differentiation was assessed by alkaline phosphatase (ALP) staining and activity. Matrix mineralization were assessed by alizarin red (ARS), von Kossa (VON), and hydroxyapatite (HA) staining. Differentiation medium was changed every 3 days.
Statistical analysis
Data were generated and analyzed by GraphPad Prism version 7.0 (GraphPad Software, San Diego, CA, USA). Statistical analysis was performed using the Mann–Whitney U test with unpaired tests. Values are shown as mean±standard error of the mean from ≥3 independent experiments. The asterisks represent the level of statistical significance (p<0.05, p<0.01, p<0.001).
RESULTS
To distinguish the basal level of BMP-related genes, we collected 2 HC-OPs and 2 AS-OPs and conducted RNA sequencing. As shown in Figure 1A, the messenger RNA (mRNA) expression of BMP3 and BMP6 was upregulated and downregulated in AS-OPs compared to HC-OPs, respectively. BMPR1A and BMPR1B expression was not different between the OP groups, whereas BMPR2 and ALK1 expressions were obviously elevated in AS-OPs (Figure 1A). We then validated BMPRs using RT-qPCR and revealed that BMPR2 expression, but not BMPR1A, BMPR1B, ALK1, or ALK2 expression, was significantly increased in AS-OPs (Figure 1B). ALK1 protein expression is statically different between HC-OPs and AS-OPs, but an increase in BMPR2 protein expression in AS-OPs was confirmed (Figure 1C). Moreover, BMPR2-expressing bone-lining cells were more abundant and statistically elevated in facet joints with AS (Figure 1D). Collectively, we found that BMPR2 expression is significantly higher in patients with AS compared to HCs.
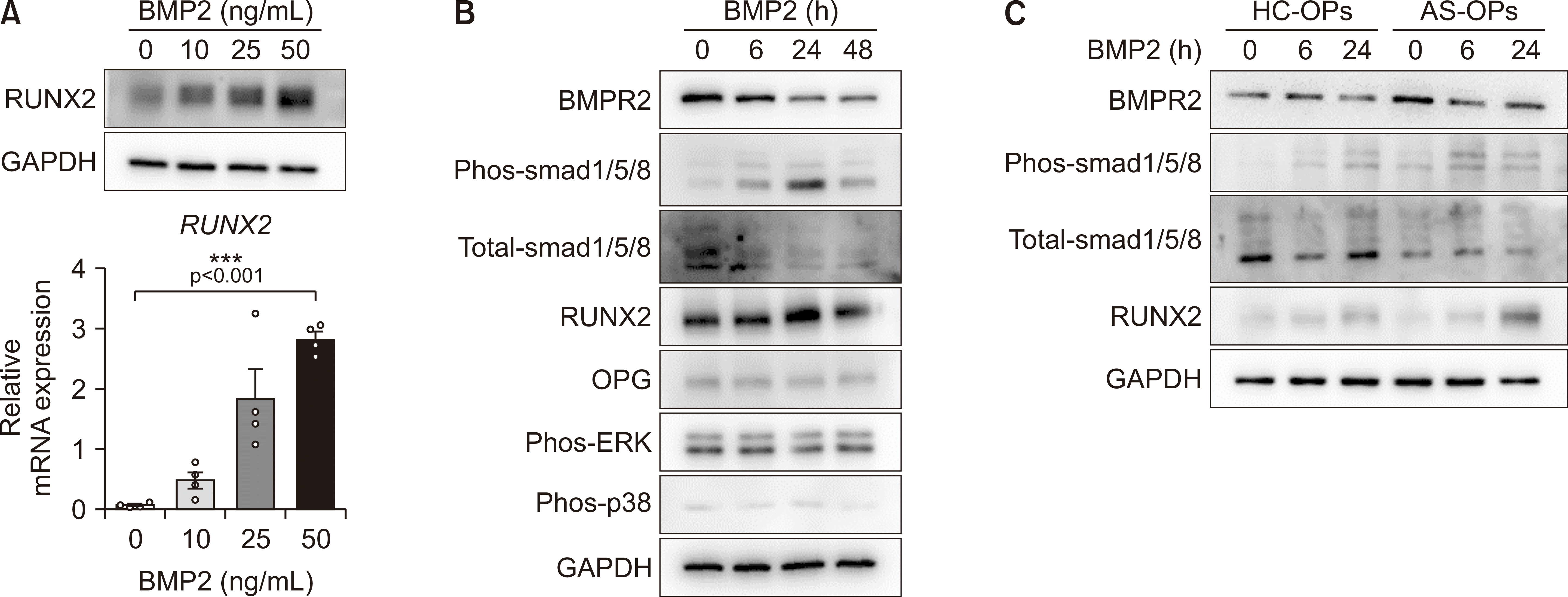
Since BMP2 is well-known as a trigger for excessive osteoblastic activity in AS, we treated AS-OPs with various BMP2 doses. BMP2 treatment dramatically upregulated the mRNA and protein expression of RUNX2 in AS-OPs (Figure 2A). There were no substantial changes in OPG, phos-ERK, and phos-p38 protein levels, but BMP2 treatment led to an increase in the phos-smad1/5/8 and RUNX2 protein concentrations of AS-OPs in a time-dependent manner as well as a decrease in total-smad1/5/8 and BMPR2 protein concentrations (Figure 2B). Consistent with a previous report [10,21], AS-OPs exhibited greater phosphorylation of smad1/5/8 and sustained BMPR2 and induction of RUNX2 protein in BMP2 treatment compared to HC-OPs (Figure 2C). These results indicate that AS-OPs have a greater capacity for induction of RUNX2 by BMP2 than do HC-OPs.
Finally, we treated both HC-OPs and AS-OPs with BMP2 (50 ng/mL) during osteogenic differentiation and observed bone matrix maturation and matrix mineralization of osteoblasts on indicated days. For matrix maturation, ALP staining and activity was markedly pronounced in AS-OPs compared to control-OPs (Figure 3A and B). For matrix mineralization, ARS, VON, and HA staining and quantification data revealed that BMP2 treatment accelerated matrix mineralization in AS-OPs compared to HC-OPs (Figure 3C and D). As expected, expression of RUNX2 and OCN mRNA was increased to a greater degree in AS-OPs (Figure 3E). Therefore, BMP2 treatment had an additive effect on both matrix maturation and matrix mineralization of AS-OPs compared to HC-OPs.
DISCUSSION
Long-term follow-up studies covering anti-TNF use have commonly reported reductions in inflammation and alleviation of ankylosis progression [22]. However, this effect cannot fully explain how chronic TNF exposure is related to new bone formation in AS. Thus, uncertainties exist regarding the precise pathogenesis of AS. Surprisingly, Lories et al. [6] highlighted BMP to play a critical role in onset and progression of spinal ankylosis, implicating that clinical application of the BMP axis for new bone formation. In line with the above, our results support the notions that BMPR2 is increased in AS-OPs and is much more sensitive to mediating bone formation activity in AS patients than HCs.
RUNX2 is a master regulator of bone formation and development [23,24]. RUNX2 is known to positively regulate ALP and OCN expression levels [25]. ALP is an early marker and a sign of osteogenesis; OCN is a late osteogenic marker that forms calcified nodule complexes. We previously reported that the ALP basal level is high in AS patients compared to RA and helps promote bone-forming activity [15,16,18,19]. In line with other previous reports, we confirmed that BMP2 is a trigger for RUNX2 expression [25]. BMP2 showed a potent function in accelerating osteoblast differentiation of mesenchymal stem cells derived from AS [10,21]. Our data highlighted that BMPs show substantially low expression in peripheral blood mononuclear cells but relatively high expression in OPs (Supplementary Figure 1).
BMP2 serum level is positively associated with proinflammatory cytokines, high bone-forming activity, and disease progression in AS patients [6,26-29]. Thus, we treated OPs with diverse proinflammatory cytokines such as TNF, interleukin-17A, and interleukin-23 but BMP2 expression in OPs was not altered following these treatments. Additionally, Ding et al. [7] reported that overexpression of BMP2 significantly induced BMPR2 expression in fibroblasts derived from AS, while we revealed that changes in BMPR1A, BMPR1B, and BMP2 mRNA expression did not differ between vehicle and BMP2 treatment groups. As shown in Figure 2, BMP2 treatment reduced BMPR2 protein expression in AS-OPs to induce upregulation of RUNX2 transcript by smad1/5/8 pathway activation. Collectively, AS-OPs showed much higher sensitivity to BMP2 for an increase in RUNX2 by smad1/5/8 signaling, reflecting high bone-forming activity in AS.
It is clear that BMPs and their receptor (ALK2; activin type I receptor) have a distinct role in promoting bone formation in AS [5]. Intriguingly, Breban et al. suggested that genetic HLA-B27 specifically interacts with BMPRs and influences the phosphorylation of SMAD proteins, which leads inflammation and ossification in AS development [30,31]. In addition, the antagonist of BMPRs exerts inhibitory functions on pathological bone features in mouse and Drosophila models of AS [30,32,33]. Less is known about the functional role of BMPR2 in AS. However, our results provide additional insight into BMP-mediated pathological bone features such as new bone formation, promoting ossification, and accelerated bone-forming activity.
This study has several limitations. First, the exact mechanisms behind the induction of the smad1/5/8-RUNX2 pathway are yet to be determined. Second, BMPR is a commonly formed heterodimer, and further molecular characterization and structure complexes of BMPR1A or BMPR1B based on BMPR2 should be determined. Third, it is unclear which stimulant can induce BMP2 expression and BMPR function. Fourth, BMP2K expression was increased in AS-OPs, but functional study remains necessary. Last, the effect of BMPR inhibition on AS in in vitro and in vivo models needs further study.
CONCLUSION
The most important finding of the current study is that BMPR2 is strongly expressed in AS-OPs, albeit with a stronger tendency towards osteoblast differentiation and bone-forming activity through smad1/5/8-RUNX2 regulation in response to BMP2 stimulation. Additionally, cytological observation revealed that BMPR2 is predominantly expressed in bone-lining cells of AS patients compared to HCs.
SUPPLEMENTARY DATA
Supplementary data can be found with this article online at https://doi.org/10.4078/jrd.2023.0024.

 XML Download
XML Download