

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Over the past few decades, cardiovascular diseases (CVDs), including coronary artery disease (CAD), stroke, heart failure, and hypertension, have been recognized as a global leading cause of death with the highest morbidity and mortality rates all around the world.1) According to World Health Organization, approximately 17.9 million deaths were attributable to CVD in 2019, representing 32% of total mortality, and the number is predicted to increase over time due to the alterations in lifestyles as well as the increasing life expectancy in modern days.2)3)4) Moreover, CVD is a costly disease that imposes a heavy financial burden on the healthcare systems, with an estimated 237 billion USD per year on the costs comprising healthcare services, medications, disability, and premature deaths, and it is expected to grow to 368 billion USD by 2035.5) The development of CVD begins with the occurrence of cardiovascular risk factors, which are known to include hypertension, obesity, diabetes, high low-density lipoprotein (LDL) cholesterol, smoking, physical inactivity, and unhealthy diet.6)7) Several risk factors often occur together, and they interact with each other to form vascular diseases that eventually lead to organ damage, terminal-organ failure, and death.7) On top of this, many recent findings have revealed the associations of the gut microbiome and its metabolites on the development of CVD and the well-established risk factors, and much evidence has suggested the possibility that the gut microbiome itself might be a risk factor for CVD.
A microbiome is a collection of symbiotic microorganisms and their associated genomes.8) There are roughly 1013 to 1014 microbial cells in each human body, which is far more than the total number of human cells. The majority of the microbiome, approximately 10–100 trillion microbial cells, are located in the gastrointestinal (GI) tract, specifically in the gut,9) where they carry out various functions that aid host health, for example, fermentation of indigestible dietary fibers, regulation of immune systems, synthesis of certain vitamins, energy metabolism and maintenance of intestinal epithelial mucosal barrier.10) The recent development in large-scale metagenomic sequencing techniques has expanded our understanding of the structural and functional capacity of the microbiome.11) It has been revealed that the gut microbiome consists of 2,172 species within 12 different phyla, most of which belong to phyla Proteobacteria, Firmicutes, Actinobacteria, Bacteroidetes, and Verrucomicrob, particularly, Firmicutes and Bacteroidetes are the 2 dominating microbes that account for more than 90% of the population.12) Through the accumulated knowledge of the structural and functional capacity of the gut microbiome, many studies have identified its association with host physiology and metabolism, both in health and disease.13) Several studies have reported that gut dysbiosis, a chronic alteration in gut microbiome composition, has a significant correlation with the pathogenesis of both intestinal disorders, including inflammatory bowel disease (IBD), irritable bowel syndrome, and coeliac disease, as well as extra-intestinal disorders including allergy, asthma, type 1 diabetes, obesity, and CVD.13)14) Among them, significant amounts of attention and research efforts were focused on the relationship between the gut microbiome and CVD, along with its related risk factors.
The advances in metagenomics and metabolomics analysis allowed the characterization of gut microbiomes and microbial-derived metabolites involved in the development of various types of CVD.15) Various research groups have demonstrated that changes in the ratio of Firmicutes to Bacteroidetes and the imbalanced levels of gut microbial metabolites, short-chain fatty acids (SCFAs), and trimethylamine N-oxide (TMAO), have been discovered to have a strong correlation with the pathogenesis of CVD.16)17)18) Alteration in the ratio of Firmicutes to Bacteroidetes representing gut dysbiosis gives rise to pathogenic species and reduces the abundance of beneficial microbes, chronically affecting the host physiology mainly through changes in the production of major microbial metabolites SCFAs and TMAO.19)20) These metabolites enter the blood circulation to influence the function of various host proteins at different sites, and they have been reported to each have a positive and negative effect on the cardiovascular system, respectively.20)
Despite continued research and effort on the relationship between CVD and gut microbiome, the exact mechanism by which the gut microbiome and its metabolites act on the pathogenesis of CVD remains undefined. Therefore, further interventions are required to establish new therapeutic and preventive options for CVDs to reduce the mortality and global burden of CVD. In this review, we focus on the compositional changes in the gut microbiome and its associated metabolites in various CVDs. Moreover, the potential treatment and preventive strategies targeting the gut microbiome and its metabolites are intensively discussed.
ALTERATION IN GUT MICROBIOME COMPOSITION IN CARDIOVASCULAR DISEASE
Coronary artery disease
Recently, multiple studies have been conducted to identify the presence of a significant correlation between gut microbiome composition and CAD. Among them, Toya et al.21) analyzed fecal samples from advanced CAD patients and controls matched for age, gender, race, and Body Mass Index using the 16s ribosomal RNA (rRNA) sequencing at the V3–V5 region. According to their findings, the ratio of Firmicutes to Bacteroidetes did not significantly differ between the 2 groups; however, there was a decrease in microbial richness and evenness as well as alterations in microbiome composition. Ruminococcus gnavus, which produces inflammatory polysaccharides and is associated with IBD, increased in relative abundance in CAD patients, while that of butyrate producers, including Lachnospiraceae NK4B4 group, Lachnospiraceae UCG-004, and Ruminococcus gauvreauii decreased compared to control.21)22)23) In contrast with R. gnavus, which has pro-inflammatory traits, the butyrate producers Lachnospiraceae NK4B4 group, Lachnospiraceae UCG-004, and R. gauvreauii possess anti-inflammatory properties that act by inhibiting pro-inflammatory immune cells while activating anti-inflammatory immune cells.24) Therefore, a decrease in butyrate producers results in a decrease in butyrate concentration, which in turn causes a variety of inflammatory disorders like hypertension, obesity, and diabetes, all of which are risk factors for CAD.25) Similarly, Emoto et al.26) carried out a case-control study to investigate the relationship between gut dysbiosis and CAD. They compared the variations in the gut microbial composition between fecal samples from patients with CAD and age- and sex-matched controls. The analyses of 16s rRNA amplicons revealed that the microbiome in the phylum Firmicutes, specifically the order Lactobacillales, including genera Lactobacillus, Streptococcus, and Enterococcus, were significantly enriched. Contrastively, the abundance of microbiomes that belong to the phylum Bacteroidetes with genera Bacteroides and Prevotella declined in patients with CAD, and they have been recognized to produce SCFAs in individuals with a high-fiber diet.27) Previously, the increased Firmicutes to Bacteroidetes ratio has been discussed to have a significant correlation with obesity, and Emoto et al.26) further discovered its relationship with other CAD risk factors, including type 2 diabetes, dyslipidemia, and hypertension. In support of the other findings, Zhu et al.28) also reported an increase in TMAO production but a decrease in SCFA production in CAD patients and highlighted their importance as potential markers for predicting the prognosis of CAD. Subsequently, they sequenced 16s rRNA obtained from stool samples of CAD patients and healthy controls to evaluate their gut microbiome composition. As a result, they discovered that Escherichia-Shigella and Enterococcus, both prospective pathogens, increased their abundance, whereas Faecalibacterium, Subdoligranulum, Roseburia, and Eubacterium rectale, which are known as butyrate producers, decreased in level in CAD patients compared to healthy controls. Furthermore, Zheng et al.29) identified an association between gut microbial dysbiosis and CAD by sequencing 16s rRNA of the V3–V4 region of fecal samples from CAD patients and healthy controls. They reported that phylum Bacteroidetes, specifically genus Butyricimonas, Chryseobacterium, Proteus, Collinsella, Megasphaera, Serratia, and Shewanella, were frequently distributed in CAD patients and dominated the gut microbial population. However, the genera of Anaerostipes and Blautia, both SCFA producers, were less abundant in CAD patients compared to healthy controls. Although the above 4 studies identified slightly different species of gut microbes that were altered in patients with CAD, they had one common finding: SCFA producers, especially butyrate producers significantly decreased in their abundance in CAD patients. This greatly supports the idea that SCFAs have a detrimental effect on cardiovascular health and are implicated in the pathogenesis of CVD.
Peripheral artery disease
Direct research on gut microbiota effects on peripheral artery disease (PAD) is still in its early stages since the relationships between microbe-derived metabolites and lower extremity PAD with major adverse cardiac events (MACEs) have been recently reported.30) It tested the concentrations of 9 metabolomic products in the plasma, including kynurenine (KYN), hippuric acid (HA), indole-3-propionic acid (IPA), indol-3-aldehyde (I3A), indoxyl sulfate (IS), 3-hydroxyanthranilic acid (HAA), p-cresyl sulfate (PCS), tryptophan, and serotonin. The result showed that PAD patients with severe complications had higher IS, HAA, and PCS and lower serotonin, KYN, IPA, I3A, and HA than the control group. From the findings, new research can be established to employ gut microbiota-derived metabolites as biomarkers of PAD. Moreover, supplementation of probiotic strains that highly produce IPA, I3A, and KYN could be successor studies to prevent PAD.30)31) New biomarkers can also be employed to predict the severity of the disease. Meanwhile, a large number of researches on gut flora impacts indicate the potential roles of the gut microbiome on PAD.32) Firstly, the mechanisms of anti-inflammation using SCFAs and reducing TMAO levels in the plasma stated above can also be applied to PAD prevention and treatment.32) Secondly, the intact epithelial layers of the gut can help to prevent atherosclerosis.32) Therefore, the administration of probiotics that support the gut barrier can also be a promising strategy to prevent PAD. Last but not least, the development of markers based on metabolites of microbiota to predict and prevent PAD can be employed to reduce the morbidity of the disease.33) To sum up, direct research on microbiomes and PAD should be intensively investigated to provide sufficient evidence for the field. Especially, gut microbiota-derived metabolites can be promising strategies to target PAD diagnosis, prevention, and treatment.
Cerebrovascular disease
Gut-brain axis has been named and well-established for more than a decade, providing deep insight into gut microbiota and brain crosstalk.34) Indeed, bacterial colonization of the gut is critical to the development and maturation of the central nervous system.35) Microbiota also affects stress reactivity and anxiety-like behavior.36) Without gut flora, memory function can be impaired because brain-derived neurotrophic factor expression is shifted.37) Multiple mechanisms have been raised.38) The gut microbiota affects the production and turnover of neurotransmitters and neurotrophic factors, and it directly influences the intestinal barrier and tight junction integrity. In addition, enteric sensory afferents are also modulated by gut microbiota. Importantly, through the immune system, the microorganism in the gut can strongly impact the brain and the vascular system. Preclinical and clinical research employing germ-free animals, probiotics, antibiotics, and infection have clarified the above mechanisms and are well documented in previous publications.38) Therefore, in this review, we only focus on the roles of gut microbiota in cerebrovascular disease.
Probiotic consumption demonstrated promising outcomes in preclinical stroke models using animals. Bifidobacterium breve, Lactobacillus casei, Lactobacillus bulgaricus, and Lactobacillus acidophilus were tested on stroke mice. The bacteria administration helped to reduce the infarct size and decrease the tumor necrosis factor-alpha (TNF-α) level.39) Another study applying Clostridium butyricum enhanced spatial learning ability, protected the hippocampus morphology of the stroke mouse model, and increased butyrate levels in the brain.40)
C. butyricum was also proven to have the same effects on diabetic mice. It improved cognitive impairment and hippocampus by upregulating p-Akt expression and downregulating caspase-3-expression, inhibiting neuronal apoptosis.41) A commercial product containing 7 bacteria strains has been examined on a mouse stroke model. The physiological outcomes were significant with spatial memory impairment and neurological dysfunction reduced. However, malondialdehyde (MDA) and brain-derived neurotrophic factor were not significantly changed.42) Inactivated lactobacillus was also employed in a rat model. Protective effects on the cerebral infarction area and neural cells were obtained. Moreover, MDA increased remarkably, and oxidative stress subsided.43)
There have been multiple studies that highlighted the vital role of SCFA-producing microorganisms in stroke patients as well. Wang et al.44) evaluated the differences in the gut microbiome composition in patients with ischemic stroke compared to their age- and sex-matched healthy controls. The result showed that Megasphaera increased in its relative abundance in stroke patients compared to the healthy controls, while that of symbiotic bacteria Bifidobacterium decreased in relative abundance. Megasphaera is a lactate-utilizing bacteria that spontaneously metabolize lactate into SCFAs such as acetate, propionate, and butyrate.45) The fact that Megasphaera serves as an SCFA producer and its level is elevated in this study is incompatible with other findings, which showed a negative correlation between CVD and SCFA levels. However, some other studies have also suggested that Megasphaera acts as an opportunistic pathogen that adversely affects an individual’s health.46) Meanwhile, Bifidobacterium, known to have health benefits in the GI tract has been recognized and used as a probiotic agent.47) Another study by Yin et al.46) on large-artery atherosclerotic ischemic stroke patients and transient ischemic attack patients reported that levels of opportunistic pathogens, including Enterobacter, Megasphaera, and Desulfovibrio increased, while the levels of beneficial bacteria such as Bacteroides, Prevotella, and Faecalibacterium decreased. Particularly, Faecalibacterium is known to produce butyrate through the fermentation of dietary fiber, and it is involved in the pathogenesis of many other diseases, including Crohn’s disease and IBD.48)49) In agreement with the 2 studies, Tan et al.50) proposed that Lactobacillaceae, Akkermansia, Enterobacteriaceae, and Porphyromonadaceae enriched while SCFA-producing bacteria including Roseburia, Bacteroides, Lachnospiraceae, Faecalibacterium, Blautia, and Anaerostipes depleted in stroke patients. Taken together, patients with cerebrovascular disease, specifically stroke, generally have increased levels of opportunistic pathogens and decreased levels of SCFA-producing beneficial bacteria, which exert anti-inflammatory influences and benefit cardiovascular health.
Heart failure
Several recent studies have identified the gut microbiome compositions and discovered their association with heart failure. Hayashi et al.51) examined the gut microbial composition of heart failure patients and age-, sex- and comorbidity-matched controls to evaluate the differences between the 2 groups. They carried out 16s rRNA gene amplicon sequencing and discovered that the patients with heart failure had significantly increased relative abundance of genus Bifidobacterium but decreased relative abundance in genus Megamonas compared to controls. Bifidobacterium is a beneficial microbiome with many physiological advantages, such as reducing the number of harmful bacteria, altering the host immune system to prevent infection, and lowering the pH and ammonia concentration in feces to enhance the intestinal environment. Megamonas, depleted in heart failure patients, are known to produce SCFAs, propionate, and acetate, through glucose fermentation. Cui et al.52) assessed and compared the fecal gut microbiome of chronic heart failure patients and healthy controls using metagenomic analyses and observed a decrease in Faecalibacterium prausnitzii and an increase in R. gnavus abundance. This finding was further supported by metabolomic analyses, which disclosed an imbalance of gut microbe metabolites with decreased butyrate concentration but increased TMAO concentration, which is analogous to Zhu et al.’s finding28) on CAD patients. Moreover, Kamo et al.53) conducted 16s rRNA gene sequencing from the fecal samples and evaluated the differences in gut microbiome composition between heart failure patients and healthy controls. They found that the relative abundance of E. rectale and Dorea longicatena were lower in heart failure patients than in healthy controls. E. rectale and D. longicatena directly or indirectly produce butyrate, which is a major source of energy for intestinal epithelial cells, and they contribute to local anti-inflammatory effects as well as epithelial barrier integrity. In summary, metagenomics studies on the gut microbial composition of heart failure patients discovered that the patients had a lower level of anti-inflammatory SCFAs and a higher level of pro-inflammatory TMAO that result from the alterations in the composition of the gut microbiome, and these changes are predicted to be involved in the development of inflammation which considerably increases the risk of CVD.
Hypertension
There have been extensive studies on the effect of alterations in gut microbiome composition on hypertension. For example, Li et al.54) discovered the correlation between dysbiosis of the gut microbiome and hypertension by transplanting the fecal microbiome from hypertensive patients into germ-free mice and analyzing its composition using metagenomic and metabolomic analyses. Compared to healthy controls, the hypertensive patients significantly decreased in microbial richness and diversity but increased in the abundance of certain pathogenic bacteria, including Prevotella and Klebsiella. Additionally, transplantation of fecal microbiome from hypertensive patients consequentially increased blood pressure in germ-free mice. Also, Yan et al.55) performed metagenomics sequencing of fecal samples from primary hypertension patients along with age-, sex-, and body-weight-matched healthy controls to analyze the differences in gut microbiome composition. Like many other studies, an increase in opportunistic pathogens, including Klebsiella, Streptococcus, and Parabacteroides merdae, and a decrease in SCFA-producing microbes such as Roseburia and F. prausnitzii were the distinctive characteristics of patients with hypertension that differ from healthy controls. Similarly, metagenomics analyses of pulmonary arterial hypertension patients conducted by Kim et al.56) showed enrichment of microbes that are associated with TMAO production but depletion of butyrate and propionate producers, including Coprococcus, Butyrivibrio, Lachnospiraceae, Eubacterium, Akkermansia, and Bacteroides. Once again, the analyses of the gut microbiome components of hypertensive patients also showed an increase in pathogenic species and TMAO producers, but a decrease in SCFA-producing microbes. These results emphasize that the concentration and balance of TMAO and SCFAs are critical in controlling the occurrence of CVD.
MECHANISM OF ACTION OF MAJOR GUT MICROBIOME METABOLITES THAT CONTRIBUTES TO CARDIOVASCULAR DISEASE
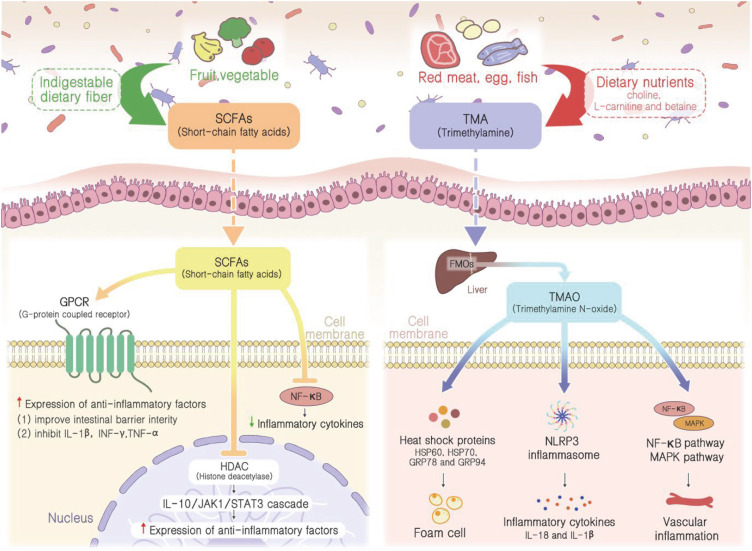
As aforementioned, numerous studies have agreed that the microorganisms in the gut can affect human health through their metabolites, including SCFAs and TMAO. These metabolites interact with various cell types to trigger specific pathways and impact the cardiovascular system (Figure 1).
Figure 1
Major signaling pathways of gut microbiome metabolites contributing to CVDs. SCFAs released by the gut microbiome can enter various cell types to inhibit HDAC and NF-κB, suppressing pro-inflammatory and enhancing anti-inflammation gene expression. In contrast, TMAO which is produced from trimethylamine triggers inflammation reactions, exacerbating CVDs.
CVD = cardiovascular disease; FMO = flavin-containing monooxygenase; GPCR = G-protein coupled receptor; GRP = glucose regulated protein; HDAC = histone deacetylase; HSP = heat shock protein; IL = interleukin; INF = interferon; MAPK = mitogen-activated protein kinase; NF-κB = nuclear factor kappa B; NLRP3 = NLR family pyrin domain containing 3; SCFA = short-chain fatty acid; TMA = trimethylamine; TMAO = trimethylamine N-oxide; TNF = tumor necrosis factor.
Short-chain fatty acids
SCFAs, including butyrate, acetate, and propionate, are gut bacterial metabolites that exhibit anti-inflammatory effects against various IBDs such as Crohn’s disease and ulcerative colitis.57) The major functions of SCFAs include the maintenance of intestinal barrier integrity, glucose, and lipid metabolism, blood pressure regulation, immune system modulation, and anti-inflammatory responses.58) SCFAs have been recently recognized to have a significant effect against various inflammatory diseases, including CAD and hypertension, suggesting their importance in maintaining health in the cardiovascular system.57) The amount of SCFAs produced is determined by the amount of dietary fiber intake along with the composition of SCFA-producing microbes in the gut flora. In this regard, alteration in gut microbiome compositions is an important factor that affects the concentration of SCFAs, therefore, contributing to the development of inflammation-related CVD.59)
There are 2 major pathways through which SCFAs can exhibit their anti-inflammatory potential. First, SCFAs are well-known histone deacetylase (HDAC) inhibitors. The histone acetylation pathway, inhibited by HDAC, enhances gene expression by decondensation of the chromatin structure and stimulates the differentiation of regulatory T cells to produce anti-inflammatory cytokines such as interleukin 10 (IL-10).57) The resulting IL-10 will bind to its cognate receptor and activate the IL-10/JAK1/STAT3 cascade to increase the expression of anti-inflammatory factors.60) Therefore, SCFAs function to enhance the production of anti-inflammatory factors by inhibiting the activity of HDAC. Second, the activation of certain G-protein coupled receptors (GPCRs) is also discovered to be involved in anti-inflammatory responses using SCFAs. SCFAs serve as ligands for GPCRs, including GPR41, GPR43, and GPR109A, which are mainly expressed in intestinal epithelial cells and nearly all types of immune cells. GPR41 and GPR43 can be activated by all 3 major SCFAs, butyrate, acetate, and propionate, whereas GPR109A is most effectively activated by butyrate.61) The binding of SCFAs to GPCRs protects the intestine from inflammation by promoting differentiation of regulatory T cells to produce IL-10 and improving intestinal barrier integrity via NLR family pyrin domain containing 3 (NLRP3) inflammasomes, which produce IL-19 involved in the maintenance of epithelial integrity by improving the function of tight junction proteins.61) Earlier research demonstrated the development of atherosclerosis and chronic heart failure due to the impaired function of the intestinal barrier, which subsequently leads to bacterial translocation and the circulation of pro-inflammatory bacterial metabolites, highlighting the importance of epithelial integrity.62) Additionally, the SCFAs are also involved in the production of anti-inflammatory gut hormones such as glucagon-like peptide 1, which promote insulin secretion and inhibit glucagon secretion, providing antagonistic effects against pro-inflammatory cytokines like IL-1β, interferon-gamma, and TNF-α that inhibits the secretion of insulin.63)64)65)
SCFAs have been shown to inhibit nuclear factor kappa B (NF-κB) activation in immune cells through histone deacetylation and lipoxygenase activation.66) Due to the inhibition, NF-κB cannot form a complete complex to translocate to the nucleus, leading to the inactivation of inflammatory genes, such as TNF-α, IL-6, and IL-8.66) Research has also shown that SCFA treatment increased anti-inflammation gene production, such as IL-10.67) Collectively, these evidences strengthen the anti-inflammation effects of SCFAs through NF-κB inhibition.
Trimethylamine N-oxide
Unlike SCFAs, TMAO is a pro-inflammatory bacterial metabolite that is also discovered to be involved in the pathogenesis of CVD. It acts by activating the immune and inflammation responses, increasing the expression of inflammatory cytokines, and inhibiting the synthesis of bile acid, eventually resulting in thrombus formation. TMAO is produced upon dependence on gut microbiomes through the metabolism of dietary nutrients, including choline, L-carnitine, and betaine, into TMA, which is further oxidized by an enzyme called hepatic flavin monooxygenase in the liver.68) The amount of TMAO generated varies between individuals depending on the degree of dietary precursor consumption and the composition of individual gut microbial flora.44) Recent metabolomics studies have evaluated the correlation between increased TMAO plasma levels and CVD as well as its linkage with MACEs such as myocardial infarction, stroke, heart failure, and cardiovascular death.18) Wang et al.69) performed a liquid chromatography-mass spectrometry analysis of plasma after supplying mice with dietary precursors of TMAO, and they discovered that the supplementation resulted in increased expression of macrophage scavenger receptors, which stimulates the accumulation of cholesterol in arterial wall upon activation.70) And the following human clinical study was able to obtain an equivalent conclusion and successfully demonstrated the association between plasma TMAO levels and the risk of CVD and MACE.71)
There are several underlying mechanisms by which increased TMAO levels can induce inflammation and atherosclerosis, both of which are the most common cause of CVD. First, TMAO regulates the expression of stress-induced heat shock protein (HSP) 60, HSP70, glucose regulated protein (GRP) 78, and GRP94. HSPs are autoantigens that activate both innate and adaptive autoimmunity and contribute to the development of atherosclerosis.72) The activation of HSPs abnormally increases the expression of pattern recognition receptors SR-A1, and cluster of differentiation 36 on the surface of macrophages stimulates the uptake of ox-LDL and results in foam cell formation, which is known as a hallmark of atherosclerosis.73)74)75) Second, TMAO also functions to produce inflammatory cytokines by activating the NLRP3 inflammasome. Upon activation, NLRP3 inflammasome cleaves inactive pro-caspase-1 into an active form of caspase-1, which automatically stimulates the maturation and secretion of IL-18 and IL-1β.18) These cytokines are then subsequently released into the cytoplasm, exerting pro-inflammatory effects on neighboring cells via paracrine signaling.76) In addition, TMAO injection was demonstrated to dramatically elevate adhesion molecules and inflammatory cytokines such as cyclooxygenase 2, IL-6, E-selectin, and ICAM1 via the MAPK and NF-κB signaling pathways, which ultimately recruit endothelial leukocytes and promote vascular inflammation.77)78) Some researchers reported that the administration of certain microbes, for example, Enterobacter aerogenes ZDY01 or Lactobacillus plantarum ZDY04, have effectively reduced the plasma TMAO concentration in mice, showing the importance of the involvement of gut microbiomes in the metabolism of TMAO.79)80) Despite these findings, probiotic treatments for reducing TMAO levels in humans have not been fully understood and require further investigation.
NEW STRATEGIES APPLYING GUT MICROBIOME IN CARDIOVASCULAR DISEASE TREATMENT AND PREVENTION
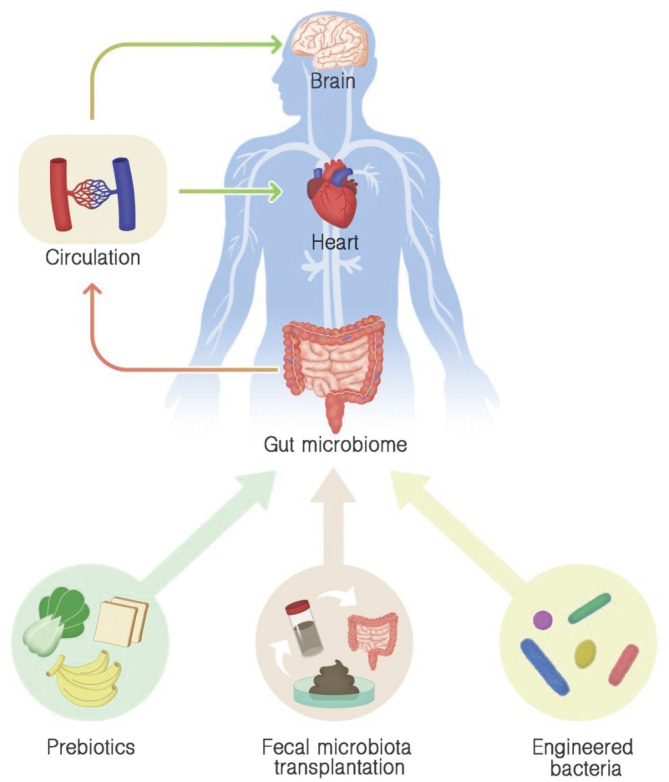
During the past few decades, various CVD treatment and prevention measures have been extensively developed, yet CVD remains the leading global cause of death; thus, more effective therapeutic strategies must be established.65)81) Accumulating evidence on the relationship between the alteration of gut microbiome composition and CVD emphasized the gut microbiome and its associated metabolites as a novel regulator of CVD, suggesting their potential as new therapeutic targets (Figure 2).
Figure 2
Modulation gut microbiome approaches in cardiovascular disease treatment and prevention. Prebiotic consumption, fecal microbiota transplantation, and engineered bacteria administration can be employed to maintain healthy gut microbiome or to improve unbalanced microbiome, leading to systemic effects on the cardiovascular system and improving cardiovascular health.
Fecal microbiota transplantation
Fecal microbiota transplantation (FMT) is the transplantation of a fecal solution from a healthy donor to the recipient’s intestinal tract, which has been used to cure Clostridium difficile infection.82) A meta-analysis research shows that FMT results in a 92% clinical resolution of C. difficile infection symptoms.83) It is believed to restore healthy gut microbiota, compete with the pathogen, and support good microorganisms in the gut, resulting in a balance gut flora. Since gut microbiome and dysbiosis link to CVD, restoring its good condition could benefit the disease treatment. Indeed, FMT was proven to alleviate myocarditis in experimental autoimmune myocarditis mouse models.84) The suggested mechanisms include escalating Firmicutes/Bacteroidetes ratio and reducing inflammatory infiltration. The study confirms the important role of Bacteroidetes in microbiota composition and the effects of gut flora on CVD through the anti-inflammation mechanism. Another study performed FMT on hypertensive rats also produced positive results.85) Transplantation of gut microbiota from healthy rats to hypertensive rats decreased blood pressure. The pro-inflammatory cytokines and reactive oxygen species have been found to contribute to the progress of the disease, indicating that balancing the gut composition by FMT can be a promising approach for hypertension treatment. Other preclinical studies strengthen the knowledge and potential of FMT by transplanting the feces of hypertensive patients and healthy individuals into germ-free mice. As predicted, the mice receiving microbiomes from the patients had higher blood pressure than those mice receiving microbiomes from the healthy ones.86) Similar results were obtained when they conducted the study on obese donors. It showed that FMT induced adiposity development in germ-free mice regardless of the food they took.87) Interestingly, the adiposity was ameliorated when the mice with a "lean" microbiome were added to the cages.87) In addition, FMT effects on atherosclerosis were also studied.88) An atherosclerosis-prone mouse model and control mice received FMT from healthy donor mice. After transplantation, atherosclerotic lesions were significantly reduced compared to before treatment. The authors also evaluated the gut flora of atherosclerosis mouse models on control mice. The result showed that the transplantation induced the progression of atherosclerosis. Collectively, it suggested the strong effects of FMT on metabolism.
Encouraged by promising preclinical outcomes, clinical studies have been conducted. A study examined the FMT strategy by transplanting the stools of healthy donors into cardiometabolic syndrome patients. The result presented an improvement in peripheral insulin sensitivity in patients with more bacterial diversity.89) Another study suggested that FMT can improve intestinal microbiota composition by conducting FMT on vegan donors to metabolic syndrome patients. However, it failed to obtain improvement in TMAO production.90) Recently, Fan et al.91) published their clinical study design of FMT effects on primary hypertension patients. The expected outcomes include microbiome profile, blood pressure, blood glucose and lipids, ankle-branchial index, as well as the mechanisms of events. This study might be one of the first clinical trials of FMT impacts on hypertension, providing important evidence for further evaluations. A modified practice of FMT called washed microbiota transplantation effect was reported on hypertensive patients.92) The method removes fecal particles, parasite eggs, and fungi from fecal suspension, and the suspension is then transferred to the patients. Although promising result on lower pressure was observed, the study did not provide the underlying mechanisms. Moreover, the preparation protocol in aerobic conditions may eliminate most of the anaerobic bacteria, which is a main limitation of the study. Hence, further examinations are needed to develop this protocol. Nevertheless, there has been no clinical trial employing FMT on heart diseases recorded on clinicaltrials.gov. The main concern could be the transfer of endotoxins or infectious agents, inducing worse effects on the recipients.93) Besides, the complex preparation procedure of FMT has been listed as a limitation of this therapy. Moreover, clinical trials might not have enough support and evidence to be conducted due to the lack of preclinical trials of FMT on CVD, such as CAD, PAD, and heart failure. Therefore, the FMT strategy in CVD treatment and prevention still needs more research.
Prebiotic consumption
Prebiotic administration can be a promising approach to CVD prevention and treatment. In general, prebiotics is substances that can support or rehabilitate healthy gut microbiota. They should be resistant to digestion in the upper sections of the GI tract, can be used by beneficial microbiota in the colon, induce good effects on host health, etc.94) They mainly include non-digestible carbohydrates obtained from plants, such as disaccharides, oligosaccharides (fructooligosaccharides, lactulose, and inulin), polysaccharides (resistant starch, pectin, and dextrin), and dietary fiber.95) They can be food for good microorganisms, such as Lactobacillus and Bifidobacterium, to ferment beneficial products. Consequently, the growth of harmful bacteria can be inhibited, leading to the balance of gut microbiota when homeostasis is affected. Indeed, researchers have shown that a diet poor in fiber is linked to hypertension and its complication through T regulatory cells.96) Moreover, the administration of a famous prebiotic—inulin, has been shown to ameliorate atherosclerosis in APOE-deficient mice by reducing plasma cholesterol and triacylglycerol levels.97) The study stated that long-chain inulin decreased 35% of the lesion area, while oligofructose-enriched inulin decreased 25% of the lesion area compared to the control.97) An active component of pectin–larch arabinogalactan (LAG) also showed therapeutic effects on the ischemia-reperfusion rat model.98) LAG strongly reduces the injured area by blocking apoptotic cascades, such as p38 phosphorylation in mitogen-activated protein kinase pathways and bax/bcl-2 ratio. This study contributes to the effect of prebiotics on CVD treatment.
Clinically, prebiotic consumption has been shown to benefit CVD patients by decreasing major risk factors. For example, a study demonstrated that oat beta-glucan decreased cholesterol, especially LDL cholesterol concentrations, in hypercholesterolemic people compared to the control group. Besides, oat beta-glucan also highly produced SCFAs in a batch model intestinal fermentation system which could be the action mechanisms of the prebiotic.99) Minolest, which contains guar gum and psyllium, was also tested on 83 hypercholesterolemic subjects.100) The result indicated that minolest lowered the cholesterol plasma level, although changes in triglyceride and body weight were insignificant. Chitosan oligosaccharides also demonstrated good effects on congenital heart disease (CHD) patients by adjusting the bacteria species in the gut microbiome, including increased Faecalibacterium, Alistipes, and Escherichia and decreased Bifidobacterium, Prevotella, and Roseburia, among others. As a result, it enhanced antioxidant capacities and lipid profiles, which subside CHD symptoms and improve patient health.101) Furthermore, prebiotic combination with probiotics has been a promising CVD prevention and treatment strategy. A mixture of prebiotic and probiotic which is also called synbiotic was tested on CAD patients with L.rhamnosus administration for 12 weeks. The result indicated that TNF-α, high sensitivity C-reactive protein, lipopolysaccharide concentrations, and Beck Depression Inventory were reduced.102) Synbiotic were also combined with L. acidophilus, L. casei, and Bifidobacterium bifidum strains to improve overweight, diabetes, and CHD patients. After 12 weeks, serum high-sensitivity C-reactive protein was decreased, and plasma nitric oxide was increased.103) Subsequently, another study examined the effects of selenium and a mixture of probiotics on type 2 diabetic patients with CHD. The outcomes were positive, with the reduction of insulin resistance and fasting plasma glucose. Besides, triglycerides, total cholesterol, and C-reactive protein were also decreased.104) Taken altogether, these studies contribute clear evidence that prebiotics and probiotics are helpful for CVD improvement. Most importantly, prebiotics provides the energy source for bacteria in the gut microbiota to produce SCFAs, improving cardiovascular health. As discussed above, SCFAs significantly affect immune system activities, intestinal membrane integrity, and metabolism. Therefore, SCFA production becomes an important mechanism of prebiotic consumption, inflammation reactions, exacerbating CVDs.
Engineered bacteria for cardiovascular disease prevention
Taking advantage of research on the gut microbiome, engineered bacteria have been studied and tested. The bacteria will be modified gene expression to produce targeted proteins or molecular compounds for disease treatment, prevention, or diagnosis. It needs to meet certain standards, such as colonization ability, safety, and therapeutic efficacy to be used. Engineered bacteria have advantages in direct modulation of gut flora, leading to stable and systemic delivery of proteins/substances. Engineered bacteria is a new and promising strategy to modulate gut microbes that can be employed to prevent CVDs. Indeed, genetically engineered bacteria have been tested to treat various diseases starting from IBD to cancer, diabetes, obesity, and among others.105) The most common bacteria involved are Escherichia coli, Lactobacillus, and Salmonella.105) For example, in 2000, Lactobacillus was used to express IL-10—an anti-inflammatory cytokine to treat IBD and obtained positive results.106) Subsequently, 15 years later, a study altered the gene expression of L. plantarum NC8 to produce angiotensin-converting enzyme inhibitory peptides and delivered the engineered bacteria to spontaneously hypertensive rats. The results indicated a significant reduction of blood pressure with no severe side effects, suggesting a potential strategy to cure hypertension.107) Although the preclinical trials have provided positive outcomes, the clinical trial is still limited. So far, only 3 clinical trials employing engineered bacteria have been found on clinicaltrials.gov, including 2 terminated studies and one completed study on cancer treatment. There have been no engineered bacteria for CVD treatment and prevention entering clinical trials. The possible reasons could be safety concerns regarding genetic modification and potential pathogenicity.105) Therefore, the remaining gap is huge, leaving high demand and challenge for the field. Since SCFAs have been proven to have important prevention effects on CVD, novel ideas for improving SCFA levels in the blood to protect the circulation system are promising.108) However, until now, it still has been a big challenge. Supplying SCFA in drinking water has failed to increase plasma SCFA concentration.109) Subsequently, oral gavage of SCFAs was also tested and showed an increase of SCFA in the blood. However, the concentration rapidly decreased after 1 hour of administration. Intraperitoneal also did not prolong the improvement of SCFA concentrations in the blood.109) Another study tried to deliver microencapsulated butyrate by oral administration.110) However, the number of patients was small (n=30 in the tested group and n=20 in the placebo control group), and there has been no follow-up study, indicating that the approach was unsuccessful. Therefore, a new strategy to deliver and maintain SCFA levels in the blood needs to be explored.
SUMMARY AND CONCLUSION
Over the past few decades, CVD has been a global leading cause of death, accounting for more than 32% of all deaths. Regardless of the continuous research and efforts in developing treatment and prevention strategies, the mortality rate and the financial burden of CVD are expected to increase continuously, emphasizing the necessity for identifying a new approach towards CVD. The recent advancement of large-scale metagenomics allowed scientists to further understand the structural and functional capacity of microbiomes in the human body, and many studies have discovered the linkage between the gut microbiome and CVD. In general, patients with CVD had decreased beneficial microbes but increased opportunistic pathogens in their intestines. More specifically, CVD patients had decreased SCFA-producing species and increased TMAO-producing microbiomes. SCFA, an anti-inflammatory metabolite, increases the expression of anti-inflammatory factors, improves intestinal barrier integrity, and inhibits pro-inflammatory cytokines. Meanwhile, TMAO, a pro-inflammatory and pro-atherosclerotic metabolite, is involved in foam cell formation, maturation, and secretion of inflammatory cytokines and adhesion molecules that result in vascular inflammation. Therefore, the imbalance of SCFA and TMAO contributes to inflammation, a characteristic risk factor for CVD.
Multiple novel treatment and prevention strategies were identified by targeting the gut microbiome and the associated metabolites. First, fecal microbiome transplantation, which transplants fecal solution from a healthy donor into the intestine of affected patients, is expected to restore healthy gut microbiota, compete with pathogens, and support good microbes in the gut. Second, prebiotic consumption, such as non-digestible carbohydrates, dietary fibers, and beneficial microbes, showed promising results in supporting and restoring healthy gut microbiota by inhibiting the growth of harmful bacteria and providing an energy source for bacteria to produce SCFA.
Although much research is being conducted on the relationship between the gut microbiome and CVD, and many novel treatment and prevention measures have been developed, it is still unclear whether the alterations in the microbiome and its related metabolites can be either the cause or the result of CVD, also, more understanding of the mechanism of how the gut microbiome and its metabolites exert their effect on the development of CVD is needed. Moreover, the number of preclinical and clinical studies is still limited. Therefore, additional clinical studies and research must be done to verify the established treatment and preventive strategies targeting the gut microbiome in the future.
 XML Download
XML Download