

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Fatigue is a physiological phenomenon caused by a lack of energy and oxidative stress [12]. Fatigue related to biological regulation and the immune system can reduce the exercise intensity and interfere with daily activities [34] and it is a social problem because it can also induce various disorders related to the immune, endocrine, autonomic nervous, and bio-regulatory systems [5]. During exercise, many energy sources (e.g., glycogen and fatty acid) are depleted, leading to physical fatigue [6]. He et al. [7] reported higher levels of oxidative stress-related biomarkers in chronic fatigue syndrome mice than in normal mice. Therefore, it can be seen that lack of energy and oxidative stress are closely related to fatigue. Regular exercise can improve the body functions, but strenuous exercise can result in the accumulation of reactive oxygen species (ROS) and lipid peroxide, which can damage organs and cause fatigue [8]. Some biomarkers including lactate dehydrogenase (LDH), lactate, ammonia, blood urea nitrogen (BUN), glucose, inflammatory cytokines, and creatine kinase (CK) are used widely to evaluate fatigue [910]. Oxidative stress results from the accumulation of nitrate radicals, superoxide anion radicals, hydroxyl radicals, and lipid peroxyl radicals such as ROS and malondialdehyde (MDA) [11]. Oxygen consumption by strenuous exercise generates reactive metabolites and free radicals that contribute to oxidative stress and inflammatory reaction [12]. Oxidative stress caused by strenuous exercise contributes to increased muscle fatigue and damage to myofibers, leading to immune system imbalance [5]. In addition, oxidative stress helps reduce cell proliferation, leading to cell death, DNA damage, causing mutation, and inflammatory reaction [13]. H2O2 is used widely as an oxidative stress inducer and inflammatory reaction stimulator because it activates the signaling pathways that generate ROS [14].
Tormentic acid (TA) is a component of Eriobotrya japonica Lindl (Loquat) [15]. Previous studies reported that TA has anti-cancer, anti-oxidant, and anti-inflammatory properties [1617]. In addition, TA inhibited the generation of ROS, expression of NADPH oxidase and inducible nitric oxide synthase, and the production of tumor necrosis factor (TNF)-α, interleukin (IL)-6, and IL-1β on rat vascular smooth muscle cells [14]. Therefore, it was hypothesized that TA would improve fatigue because fatigue is closely related to the oxidative and inflammatory responses. This study examined the anti-fatigue effect of TA using myoblast C2C12 cells and treadmill stress test (TST) and forced swimming test (FST) models.
MATERIALS AND METHODS
C2C12 cell culture
C2C12 cells were grown in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin (Sigma, St. Louis, MO, USA) at 37°C in 5% CO2. When the C2C12 cells reached approximately 80% confluence, they were rinsed with phosphate-buffered saline (PBS) and incubated with DMEM containing 2% horse serum (HS, 12449C, Sigma-Aldrich, St. Louis, MO, USA) to induce C2C12 cell differentiation. After 4 days, differentiated C2C12 cells were pre-treated with prepared TA (0.01, 0.1, and 1 μg/mL; Chemfaces, Wuhan, China) for 1 h and then stimulated with H2O2 for 24 h [15].
Fatigue-related biochemical indicators analysis in cell extract, serum, muscle, or liver
Cells were rinsed with PBS and lysed in radioimmunoprecipitation assay buffer (Sigma) containing protease inhibitors. The extracted proteins were assayed using the bicinchoninic acid (Sigma) method as reported previously [18]. Blood was collected from the heart and centrifuged at 800 g for 20 min, and the serum was isolated. The muscle and liver tissues were also collected. The amount of aspartate aminotransferase (AST), alanine transaminase (ALT), glucose, CK, BUN, and LDH in the serum was measured by a DRI CHEM NX500 analyzer (Fujifilm, Tokyo, Japan). The amount of superoxide dismutase (SOD; DoGenBio, Seoul, Korea), MDA (DoGenBio), catalase (CAT; DoGenBio), free fatty acid (FFA; Abcam, Cambridge, UK), glycogen (Biovision Inc., Milpitas, CA, USA), citrate synthase (MyBiosource Inc., San Diego, CA, USA), lactate (DoGenBio), and cortisol (Sigma Chemical Co., St. Louis, MO, USA) were measured using each assay kit. A serum sample (10 μL) and tissue samples from the liver and muscle (50 μL) were used for each experiment.
Enzyme-linked immunosorbent assay (ELISA)
Cytokine antibodies were purchased from BD Biosciences Pharmingen (San Diego, CA, USA). After overnight incubation of the capture antibody at 4°C, the wells were washed twice with PBS containing 0.05% Tween 20 and blocked with 180 μL of 10% FBS diluted in PBS for 2 h at room temperature. The samples or recombinant cytokines were added to each well and incubated for 4 h at room temperature. After wash the wells, secondary antibodies were incubated, and the horseradish peroxidase-conjugated antibody was then added. After washing 5 times, 3,3,5,5'-tetramethylethylenediamine (BD Biosciences Pharmingen) substrate solution was added, and a stop solution was applied to read at 450 nm.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
MTT (Sigma Chemical Co.) assay is a classical method used for cell viability determination, by testing the activity of dehydrogenase enzymes. The cytotoxic effect of TA on C2C12 cells was analyzed using an MTT assay. The differentiated C2C12 cells were seeded at a density of 3 × 105 cells/well into 24-well plates, and allowed to adhere for 24 h. The cells were treated with TA for 24 h. MTT was then added to each well, as performed previously [1920]. An insoluble purple formazan was solubilized in dimethyl sulfoxide, and the absorbance at 570 nm for each well was then recorded using an ELISA reader.
Animals
Male ICR mice (4-week-old, 21.5 ± 1.5 g) were obtained from the Dae-Han Experimental Animal Center (Eumsung, Korea) in a specific pathogen-free environment. They were kept in individual cages (n = 5/cage) for 1 week under controlled environmental conditions of 22 ± 2°C and 50 ± 5% humidity, and a 12:12 h light: dark cycle. The animals were given tap water and standard laboratory food ad libitum (Nestlé Purina, St. Louis, MO, USA). All animal research was performed according to the guidelines for the care and use of laboratory animals approved by the Animal Ethics Committee of Kyung Hee University [KHUASP (SE)-20-263]. The group without TST or FST was considered the normal group because it did not induce fatigue. The control group was considered a group in which fatigue was induced by performing TST or FST. The TA group was the group that orally administered TA before inducing fatigue by performing TST or FST.
TST
The mice in each group were forced to run on the treadmill for 30 min once a week for 4 weeks. The pattern of the loaded exercise was composed of forced running at a 10 m/min during the 10 min, in sequence 16 m/min during the 10 min, and last 21 m/min during the 10 min. On the 28th day, the speeds used to determine exhaustion were 10 m/min for 5 min, in sequence for 3 min each, then 40 m/min during the 30 min [10]. TA (1 mg/kg) or distilled water (control group) was administered orally every day for 28 days. The exhaustion time of TST was measured on the last day of this experiment. The TA dose was determined to be 1 mg/kg according to a previous report [15].
FST
The mice were split into TA (1 mg/kg) or distilled water (control group) groups based on the recorded swimming times after the initial measurement at the immobility time. The mice in each group had FST once a week for 6 min for 4 weeks. The device was comprised of 2 Plexiglas cylinders placed side-by-side in a Makrolon cage that was filled with water to a height of 10 cm, at a temperature of 23 to 25°C. After a 2 min delay, the immobility was timed for 4 min. A trained observer unaware of the experimental treatments, used a stopwatch to time the periods of immobility [10]. TA (1 mg/kg) or distilled water (control group) was administered orally simultaneously every day for 28 days. The immobility time of FST was measured on day 28.
Statistical analysis
The in vitro experimental results are expressed as the mean ± standard error mean (SEM) from the results of at least 3 independent experiments. The in vivo experimental results were presented as the mean ± SEM (n = 5/group). The statistical analysis of data was done using SPSS 12.0 software (SPSS, Chicago, IL, USA). One-way analysis of variance, followed by Tukey’s multiple comparisons test and the independent t-test were used to analyze the fatigue improvement effects of TA. P-values < 0.05 were considered significant.
RESULTS
Anti-fatigue and anti-inflammatory effects of TA in H2O2-stimulated C2C12 cells
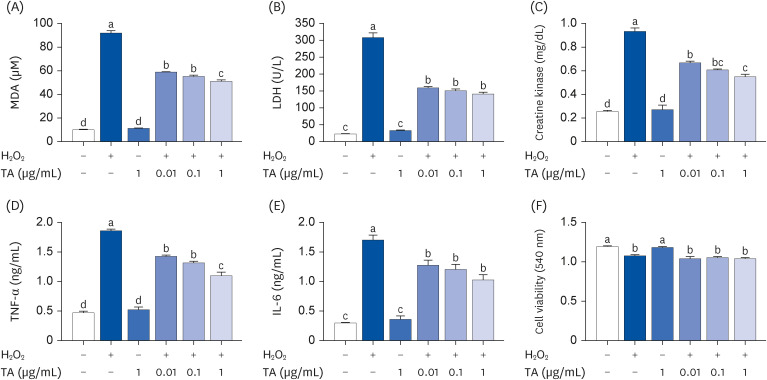
First, this study investigated whether TA would show an anti-fatigue effect in the in vitro model. The levels of fatigue-related biomarkers were determined in H2O2-stimulated C2C12 cells. MDA is a biomarker of oxidative stress and fatigue. A considerable amount of MDA was significantly increased by H2O2 stimulation, whereas the increased MDA levels were remarkably decreased by treatment with TA (Fig. 1A, P < 0.05). The fatigue biomarkers, LDH and CK are produced by oxidative stress. The levels of LDH and CK were remarkably increased by H2O2 stimulation, whereas TA significantly down-regulated the levels of LDH and CK from the H2O2-stimulated cells (Fig. 1B and C, P < 0.05). In addition, the levels of inflammatory cytokines (TNF-α and IL-6) were significantly down-regulated by the treatment with TA on H2O2-stimulated C2C12 cells (Fig. 1D and E, P < 0.05). TA did not influence the cell viability of C2C12 cells (Fig. 1F). TA alone did not affect the fatigue-related biomarkers and cell viability (Fig. 1).
Fig. 1
Effect of TA on the production of fatigue-related factors and inflammatory cytokines in H2O2-stimulated C2C12 cells. After 4 days, differentiated C2C12 cells (3 × 105) were pretreated with TA (0.01, 0.1, and 1 μg/mL) for 1 h and then treated with H2O2 for 24 h. (A-C) The levels of MDA, LDH, and CK were analyzed by enzyme-linked immunosorbent assay. (D, E) The levels of TNF-α and IL-6 were analyzed by ELISA. (F) C2C12 cells (3 × 105) were pretreated with TA (0.01, 0.1, and 1 μg/mL) for 1 h and then treated with H2O2 for 24 h. Cell viability was determined using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Each datum represents the mean ± standard error mean of 3 independent experiments.
TA, tormentic acid; MDA, malondialdehyde; LDH, lactate dehydrogenase; CK, creatine kinase; TNF, tumor necrosis factor; IL, interleukin.
The different letters above the bars indicate significant intergroup differences as determined by a Tukey’s multiple comparisons test (P < 0.05).
Anti-oxidative effects of TA in H2O2-stimulated C2C12 cells
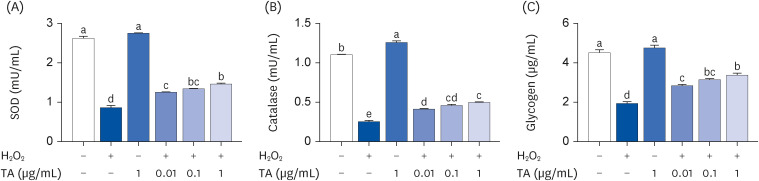
Next, this study investigated whether TA would show an antioxidative effect in the in vitro model. The levels of oxidative stress-related biomarkers were determined in H2O2-stimulated C2C12 cells. SOD is an important antioxidant enzyme that protects against superoxide toxicity [21]. CAT plays a major role in cellular anti-oxidant defense by decomposing H2O2 [22]. The levels of SOD and CAT were significantly decreased by H2O2 stimulation, whereas their decreased levels were remarkably increased by the TA treatment in a dose-dependent manner (Fig. 2A and B, P < 0.05). The glycogen levels, an anti-fatigue-related biomarker, was significantly increased by the TA treatment on H2O2-stimulated C2C12 cells (Fig. 2C, P < 0.05).
Fig. 2
Effect of TA on the levels of SOD, CAT, and glycogen in H2O2-stimulated C2C12 cells. After 4 days, differentiated C2C12 cells (3 × 105) were pretreated with TA (0.01, 0.1, and 1 μg/mL) for 1 h and then treated with H2O2 for 24 h. The levels of (A) SOD, (B) CAT, and (C) glycogen were analyzed by enzyme-linked immunosorbent assay. Each datum represents the mean ± standard error mean of 3 independent experiments.
TA, tormentic acid; SOD, superoxide dismutase; CAT, catalase.
The different letters above the bars indicate significant intergroup differences as determined by a Tukey’s multiple comparisons test (P < 0.05).
Anti-fatigue effects of TA in TST and FST animal models
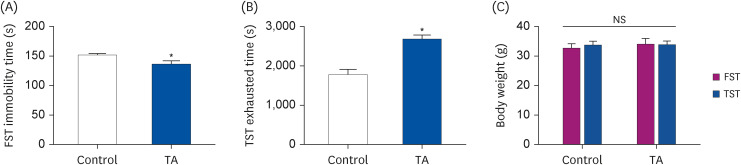
Based on the above in vitro experiment, this study examined the anti-fatigue effects of TA in the TST and FST animal models. A daily dose of TA (1 mg/kg) or distilled water was orally administered for 28 days. The immobility times in the FST model were significantly lower in the TA-administered group than in the control group (Fig. 3A, P < 0.05). Furthermore, the administration of TA resulted in a significant increase in TST exhaustion time (Fig. 3B, P < 0.05). On the other hand, the body weight was similar to the control group (Fig. 3C, P < 0.05).
Fig. 3
Effect of TA on the exercise ability of TST and FST. (A) FST immobility time and (B) TST exhaustion time were measured by trained researchers. (C) The body weight after TST and FST. There were measured on the last day of the experiment. The values are the mean ± standard error mean (n = 5 per group). Control group: TST- or FST-performing group; TA group: TA-administered TST- or FST-performing group.
TA, tormentic acid; TST, treadmill stress test; FST, forced swimming test; NS, no significance.
*P < 0.05, significantly different from the control mice (independent t-test).
Anti-oxidative effects of TA in TST and FST animal models
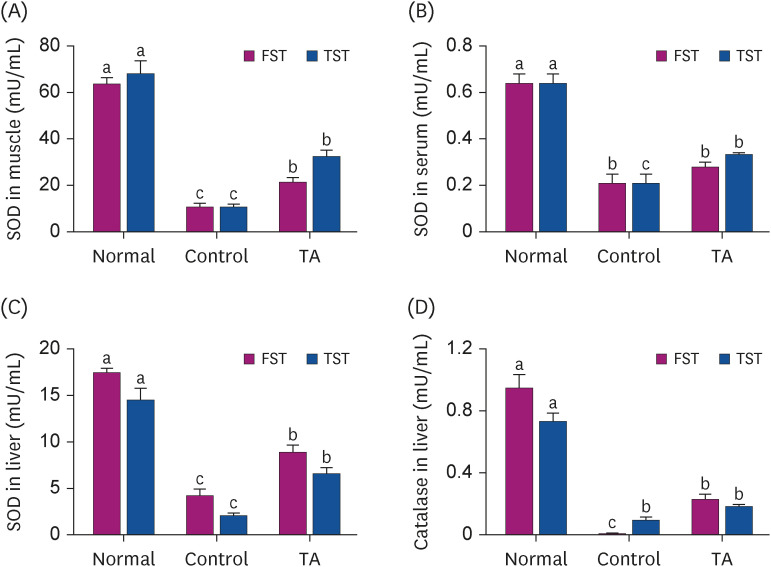
This study examined whether the anti-fatigue effect of TA is related to the anti-oxidative reaction in TST and FST animal models. The administration of TA significantly increased the activities of SOD compared to the control group in the muscle, serum, and liver in the TST or FST animal model (Fig. 4A-C, P < 0.05). In addition, the levels of CAT in the liver of TA-administered mice were remarkably increased compared to the control group in the FST animal model (Fig. 4D, P < 0.05).
Fig. 4
Effect of TA on the SOD and CAT activities after TST and FST. The activities of SOD in (A) muscle, (B) serum, and (C) liver. (D) The activities of CAT in the liver. The values are the mean ± standard error mean (n = 5 per group). Normal group: non-TST or non-FST group; Control group: TST- or FST-performing group; TA group: TA-administered TST- or FST-performing group.
TA, tormentic acid; SOD, superoxide dismutase; CAT, catalase; TST, treadmill stress test; FST, forced swimming test.
The different letters above the bars indicate significant intergroup differences as determined by a Tukey’s multiple comparisons test (P < 0.05).
Improvement effect of TA on energy utilization
This study examined whether the anti-fatigue effect of TA is related to energy utilization in the TST and FST animal models. As shown in Fig. 5A, the serum glucose contents of TA-administered group were significantly lower than the control group (P < 0.05). The levels of FFA significantly decreased in the control group the decreased levels were significantly increased in the TA-administered group (Fig. 5B, P < 0.05). In the muscle, the levels of glycogen were remarkably higher in the TA-administered group than in the control group (Fig. 5C, P < 0.05). TA increased the activity of citrate synthase, but the increase was not significant (Fig. 5D).
Fig. 5
Effect of TA on the levels of glucose, FFA, glycogen, and citrate synthase after TST and FST. (A) Serum levels of glucose were analyzed by a DRI CHEM NX500 analyzer. (B) Levels of FFA in serum were analyzed using commercially available kits. (C) Levels of glycogen and (D) activity of citrate synthase in the muscle were analyzed using the appropriate kits. The values are the mean ± standard error mean (n = 5 per group). Normal group: non-TST or non-FST group; Control group: TST- or FST-performing group; TA group: TA-administered TST- or FST-performing group.
TA, tormentic acid; FFA, free fatty acid; TST, treadmill stress test; FST, forced swimming test.
The different letters above the bars indicate significant intergroup differences as determined by a Tukey’s multiple comparisons test (P < 0.05).

TA reduced fatigue-inducing factors in TST and FST animal models
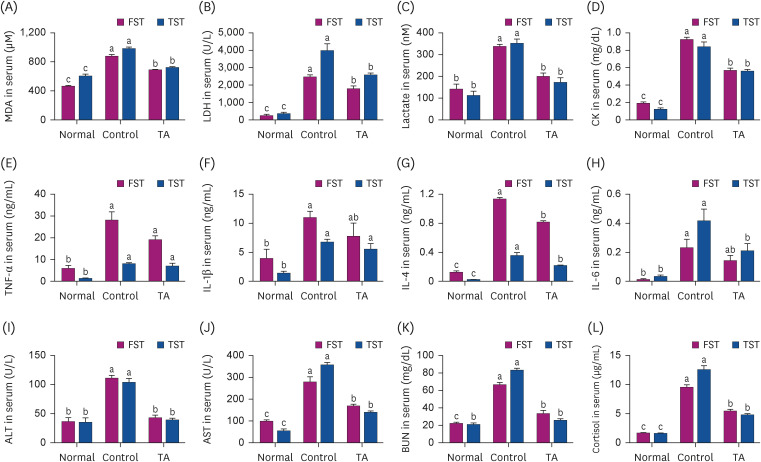
Finally, this study investigated whether TA could regulate fatigue-inducing factors in animal models. MDA, LDH, and lactate are major biomarkers associated with fatigue. As shown in Fig. 6A-C, TA remarkably decreased the levels of MDA, LDH, and lactate compared to the control group (P < 0.05). CK is related to muscle injuries. The levels of CK in the serum were significantly higher in the control group than in the TA group (Fig. 6D, P < 0.05). The serum levels of IL-4 and IL-6 were significantly down-regulated by TA administration compared to the control group but not TNF-α and IL-1β (Fig. 6E-H, P < 0.05). ALT and AST are particularly associated with liver injuries [23]. As shown in Fig. 6I and J, the serum levels of ALT and AST increased by FST or TST were down-regulated by TA administration (P < 0.05). The serum levels of BUN were remarkably increased by FST or TST in the control group, whereas the increased levels were down-regulated by TA administration (Fig. 6K, P < 0.05). Furthermore, the levels of cortisol in the serum were decreased in the TA-administered group compared to the control group (Fig. 6L, P < 0.05).
Fig. 6
Effect of TA on the levels of fatigue-inducing factors in serum after TST and FST. (A) MDA, (B) LDH, (C) lactate, and (D) CK were measured with each kit. The levels of (E) TNF-α, (F) IL-1β, (G) IL-4, and (H) IL-6 were analyzed by enzyme-linked immunosorbent assay. (I) ALT, (J) AST, (K) BUN, and (L) cortisol were measured using the appropriate kits. The values are the mean ± standard error mean (n = 5 per group). Normal group: non-TST or non-FST group; Control group: TST- or FST-performing group; TA group: TA-administered TST- or FST-performing group.
TA, tormentic acid; FFA, free fatty acid; TST, treadmill stress test; FST, forced swimming test; MDA, malondialdehyde; LDH, lactate dehydrogenase; CK, creatine kinase; TNF, tumor necrosis factor; IL, interleukin; ALT, alanine transaminase; AST, aspartate aminotransferase; BUN, blood urea nitrogen.
The different letters above the bars indicate significant intergroup differences as determined by a Tukey's multiple comparisons test (P < 0.05).
DISCUSSION
Fatigue is a common phenomenon commonly observed in healthy and unhealthy people, affecting individual performance and quality of life. Fatigue is a phenomenon caused by oxidative stress [9]. This study showed that TA has an anti-fatigue effect, which is related to its anti-oxidative effect and energy metabolism property.
Biochemical molecules, including MDA, LDH, CK, inflammatory cytokines, SOD, CAT, glycogen, glucose, FFA, and citrate synthase, are closely related to fatigue [10]. MDA is considered an indicator of oxidative stress and fatigue [24]. LDH is an enzyme that catalyzes the production of lactic acid, and an increase in LDH activity caused by strenuous exercise induces myoblast cell death and promotes muscle disruption [25]. Lactate increased by LDH is a major source of systemic energy and critical in anaerobic exercise. Elevated lactate concentration is considered the most important biomarker of muscle fatigue during exercise [26]. CK is an important enzyme indicative of muscle damage [27]. An increase in CK in the blood is considered an indirect indicator of muscle damage [28]. Therefore, MDA, LDH, lactate, and CK are useful biomarkers in strenuous exercise physiology and sports medicine for the detecting muscle injury and overwork [2429]. Previous studies showed that the levels of proinflammatory cytokines including TNF-α, IL-1β, IL-4, and IL-6 were elevated in patients with chronic fatigue syndrome and fatigue animal models [103031]. Powers et al. [32] reported that oxidative stress caused by ROS or free radicals augments the inflammatory response and aggravates fatigue. SOD catalyzes the conversion of superoxide radicals to H2O2 which acts a key role in defense against superoxide radicals [20]. CAT catalyzes the formation of H2O2 from O2 [22]. He et al. [7] reported that cells have enzyme antioxidants such as SOD, CAT, and glutathione peroxidase to neutralize excess ROS and reduce oxidation. These anti-oxidants also alleviate the inflammatory reaction [7].
During strenuous exercise, the aerobic energy supply turns to anaerobic glycolysis in the muscle, and muscle glycogen is rapidly consumed, producing large amounts of lactic acid [33]. The accumulation of lactic acid in the muscles and blood causes muscle fatigue. Glycogen provides energy during exercise as a form of energy storage [34]. Exercise promotes gluconeogenesis and glycolysis to meet the body’s energy needs. During excessive exercise, stored glycogen is broken down into glucose to be used as an alternative energy source [35]. During exercise, FFA are hydrolyzed from triacylglycerol and released into the circulatory system as an energy source for muscles [36]. Skeletal muscle function is significantly influenced by mitochondrial biogenesis and mitochondrial dysfunction and reduced muscular contractility are symptoms of fatigue [37]. In addition, impaired mitochondrial function in skeletal muscle reduces mitochondrial biogenesis, expression of mitochondrial respiration complex components, and ATP levels, resulting in fatigue. Zhu et al. [38] reported that Lepidium meyenii Walp improved fatigue by enhancing the mitochondrial function via mitochondrial dynamics. Citrate synthase is an important enzyme of the mitochondrial tricarboxylic acid cycle (citric acid cycle) and has often been used as a mitochondrial marker in human and animal research. Citrate produced by citrate synthase plays an important role in ATP synthesis through mitochondrial matrix oxidation [39]. Although not significant, TA improved citrate synthase activity in the present study. Therefore, an increase in anti-oxidants, such as SOD and CAT, and regulation of energy metabolism will help improve fatigue by reducing the fatigue-inducing factors. Compared to the control group, TA down-regulated the levels of MDA, LDH, CK, inflammatory cytokines (IL-4 and IL-6), and glucose. The levels of SOD, CAT, glycogen, and FFA decreased by oxidative stress were up-regulated by TA in in vitro and/or in vivo models. Therefore, we suggest that TA has an anti-fatigue effect by the increasing anti-oxidative effect and regulating the energy metabolism.
Elevated serum levels of AST and ALT reflect hepatocytotoxicity [40]. An increased in the serum levels of BUN reflect protein breakdown, adversely affecting muscle contraction strength and causing fatigue [2]. Bekker et al. [41] reported that cortisol is an indicator of the stress response. High cortisol levels are observed in various chronic conditions, such as anxiety, hypertension, type 2 diabetes, depression, and chronic fatigue syndrome [1042]. Thus, there is a strong relationship between high cortisol levels and fatigue. Hsu et al. [42] reported that capsaicin has an anti-fatigue function by regulating lactate, ammonia, glucose, BUN, CK, AST, ALT, and albumin. In the current study, TA down-regulated the levels of AST, ALT, BUN, and cortisol in the TST and FST models. Therefore, we propose that TA has an anti-fatigue effect by protecting against tissue damage caused by oxidative stress.
Numerous studies on materials with an anti-fatigue effect have been reported. Qu et al. [43] reported that the Auxis thazard oligopeptide reduces oxidative stress and enhances energy metabolism to prevent fatigue. Lepidium meyenii Walp has anti-fatigue properties by reducing mitochondria-mediated muscle damage and oxidative stress [38]. Taking phytochemicals, such as curcumin, capsaicin, and resveratrol, increase the forelimb grip strength and endurance in an FST animal model [424445]. Therefore, we can suggest that TA has an anti-fatigue impact by boosting anti-oxidant activity and energy metabolism, which is similar to earlier studies. Unlike prior research on fatigue, this study did not examine food intake, despite food intake analysis being required to confirm the anti-fatigue effect of TA. Therefore, it is necessary to analyze food intake in future research.
In this study, TA down-regulated the fatigue causative factors, and up-regulated the fatigue improvement factors in both in vivo and in vitro models. TA up-regulated the energy metabolism-related factors. Furthermore, TA blocked the tissue damage caused by oxidative stress in TST and FST models. In conclusion, TA improves fatigue by exhibiting an anti-oxidative effect and energy metabolism-modulating properties. Therefore, we suggest that TA can be a powerful substance in health functional foods and therapeutics to improve fatigue.
 XML Download
XML Download